
Identification of the EdcR Estrogen-Dependent Repressor in Caenibius tardaugens NBRC 16725: Construction of a Cellular Estradiol Biosensor


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Growth Media
2.3. DNA Manipulation
2.4. RNA Manipulation
2.5. Construction of C. tardaugens Knockout Strains
2.6. Complementation of C. tardaugens Knockout Strains
2.7. Fluorescence Measurement of Cultures on a Plate Reader
2.8. In Silico Genomic Analysis
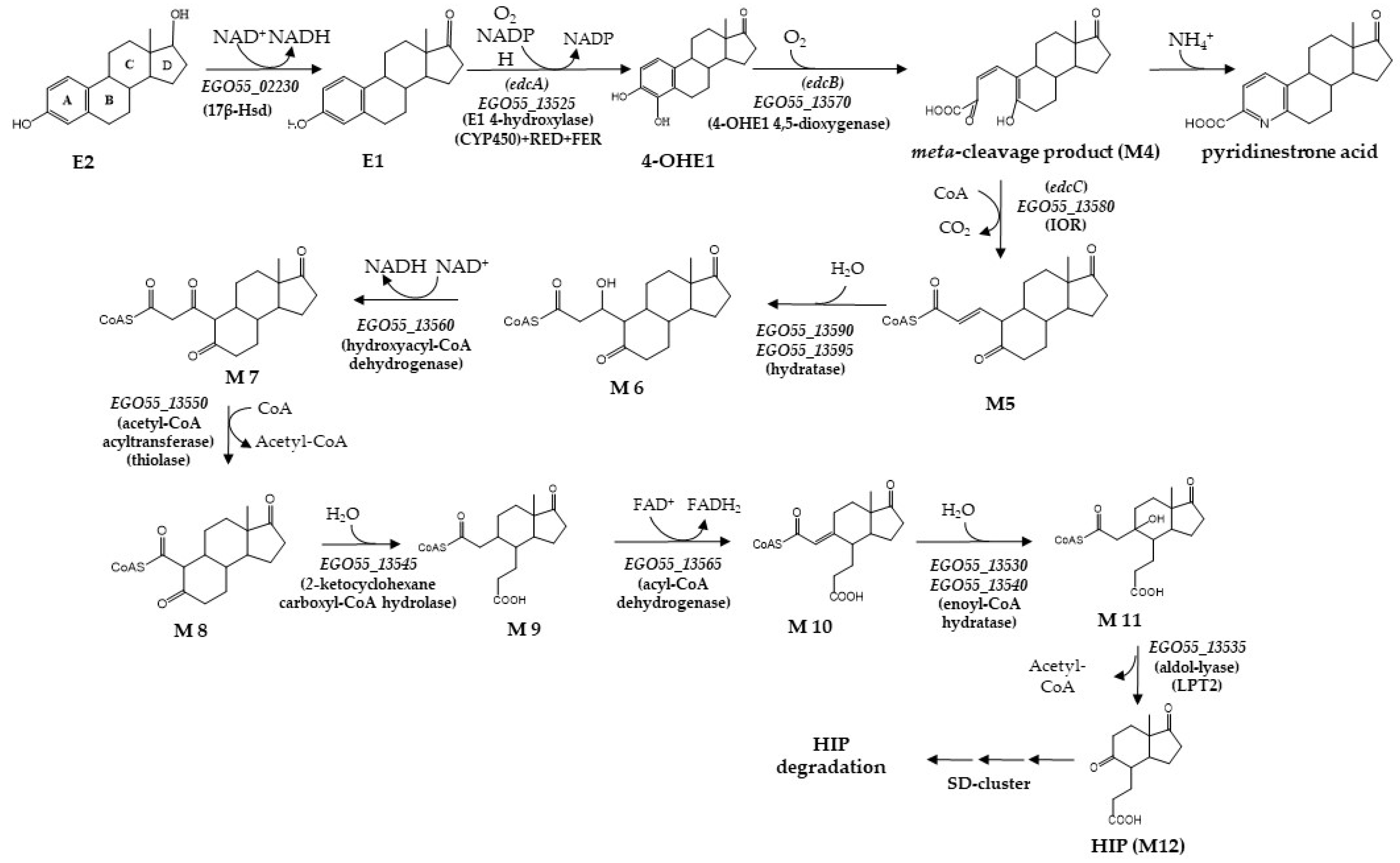
3. Results
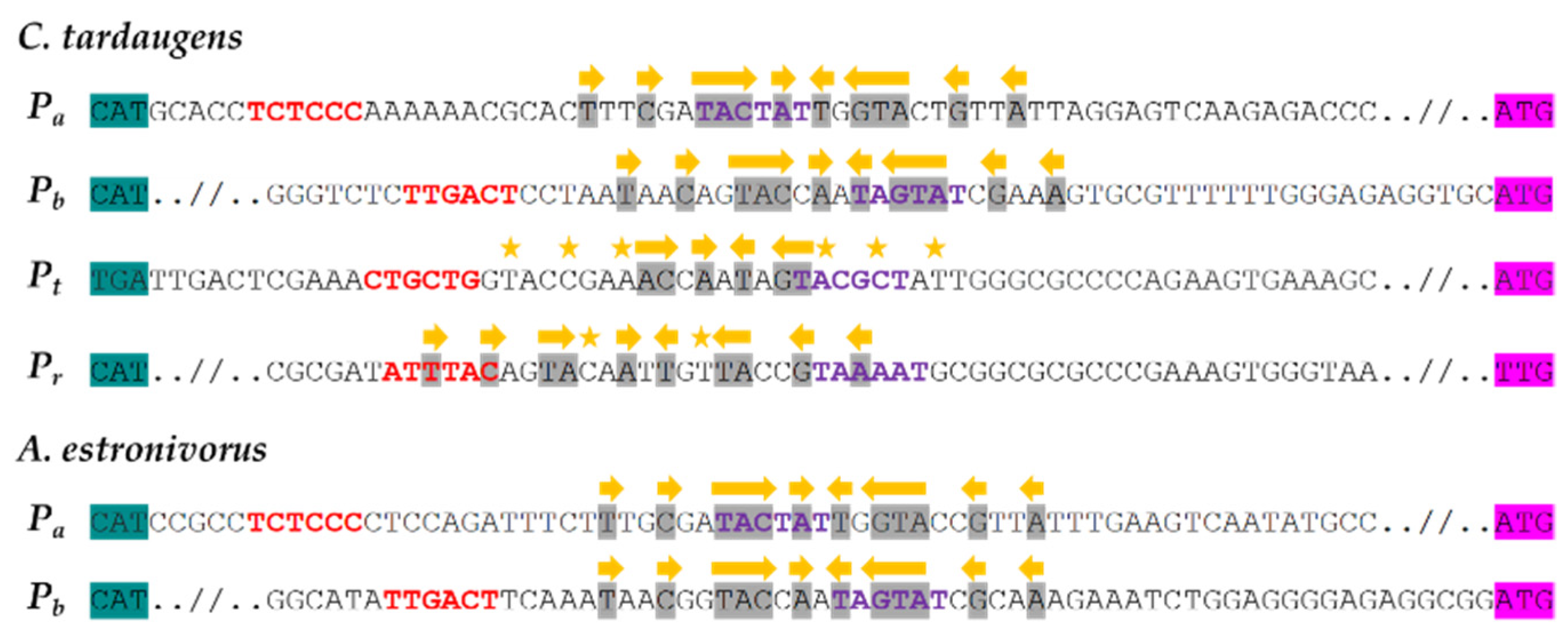
3.1. In Silico Analysis of the Main Promoters Responsible for the Expression of C. tardaugens Edc Gene Cluster
3.2. Analysis of the EdcR Function
3.3. Analysis of the EdcR Effectors
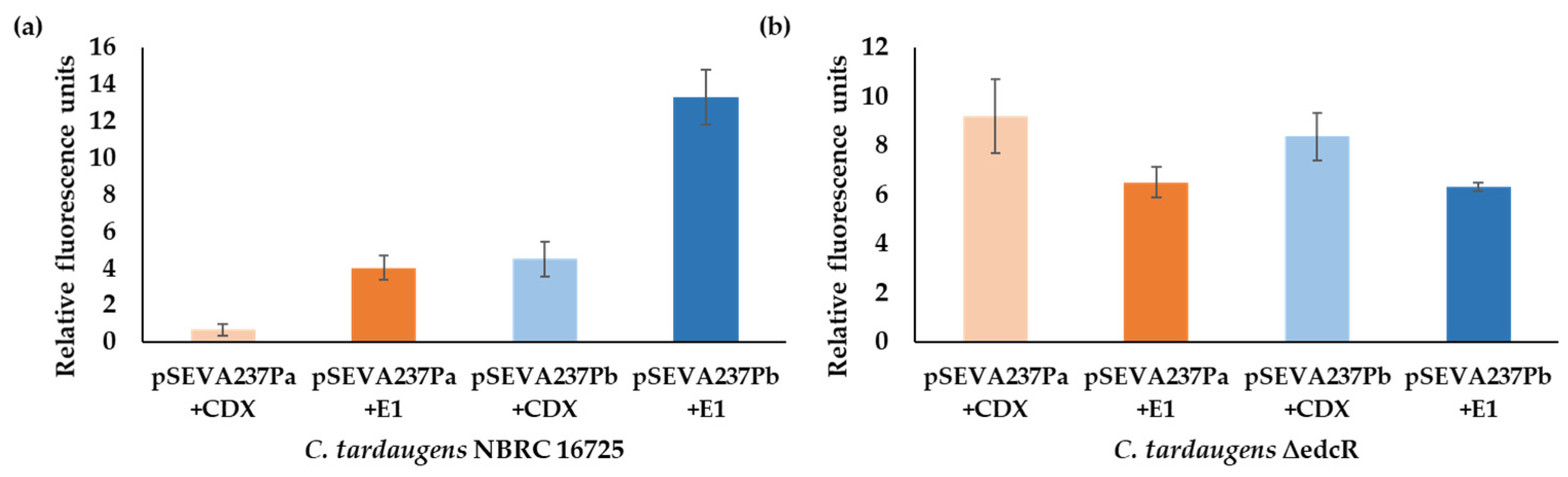
3.4. Analysis of the Promoters of the Edc Cluster in E. coli. Construction of an Estrogen Biosensor
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, L.H.M.L.M.; Araújo, A.N.; Fachini, A.; Pena, A.; Delerue-Matos, C.; Montenegro, M.C.B.S.M. Ecotoxicological aspects related to the presence of pharmaceuticals in the aquatic environment. J. Hazard. Mater. 2010, 175, 45–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Casey, F.X.M.; Hakk, H.; Larsen, G.L. Persistence and fate of 17β-estradiol and testosterone in agricultural soils. Chemosphere 2007, 67, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.M.; Chapelle, F.H.; Barber, L.B.; McMahon, P.B.; Gray, J.L.; Kolpin, D.W. Biodegradation of 17β-estradiol, estrone, and testosterone in stream sediments. Environ. Sci. Technol. 2009, 43, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Wu, S.; Chang, H.; Hu, J. Behaviors of glucocorticoids, androgens and progestogens in a municipal sewage treatment plant: Comparison to estrogens. Environ. Sci. Technol. 2011, 45, 2725–2733. [Google Scholar] [CrossRef]
- Fujii, K.; Kikuchi, S.; Satomi, M.; Ushio-Sata, N.; Morita, N. Degradation of 17β-estradiol by a gram-negative bacterium isolated from activated sludge in a sewage treatment plant in Tokyo, Japan. Appl. Environ. Microbiol. 2002, 68, 2057–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahrbach, M.; Kuever, J.; Meinke, R.; Kampfer, P.; Hollender, J. Denitratisoma oestradiolicum gen. nov., sp. nov., a 17beta-oestradiol-degrading, denitrifying betaproteobacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-P.; Roh, H.; Chu, K.-H. 17β-estradiol-degrading bacteria isolated from activated sludge. Environ. Sci. Technol. 2007, 41, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Wang, P.; Liu, D.; Gao, R.; Shao, H.; Zhao, H.; Ma, Z.; Wang, D.; Huo, H. Degradation characteristics and metabolic pathway of 17β-estradiol (E2) by Rhodococcus sp. DS201. Biotechnol. Bioprocess Eng. 2016, 21, 804–813. [Google Scholar] [CrossRef]
- Kurisu, F.; Ogura, M.; Saitoh, S.; Yamazoe, A.; Yagi, O. Degradation of natural estrogen and identification of the metabolites produced by soil isolates of Rhodococcus sp. and Sphingomonas sp. J. Biosci. Bioeng. 2010, 109, 576–582. [Google Scholar] [CrossRef]
- Chen, Y.L.; Yu, C.P.; Lee, T.H.; Goh, K.S.; Chu, K.H.; Wang, P.H.; Ismail, W.; Shih, C.J.; Chiang, Y.R. Biochemical mechanisms and catabolic enzymes involved in bacterial estrogen degradation pathways. Cell Chem. Biol. 2017, 24, 712–724.e7. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zheng, D.; Liang, R. Isolation and characterization of an estrogen-degrading Pseudomonas putida strain SJTE-1. 3 Biotech 2019, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, J.; Williams, M.A.; Ling, W.; Sun, K.; Lu, C.; Gao, Y.; Waigi, M.G. Metabolism of 17β-estradiol by Novosphingobium sp. ES2-1 as probed via HRMS combined with 13C3-labeling. J. Hazard. Mater. 2020, 389, 121875. [Google Scholar] [CrossRef]
- Qin, D.; Ma, C.; Lv, M.; Yu, C.-P. Sphingobium estronivorans sp. nov. and Sphingobium bisphenolivorans sp. nov., isolated from a wastewater treatment plant. Int. J. Syst. Evol. Microbiol 2020, 70, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Ma, C.; Hu, A.; Zhang, F.; Hu, H.; Yu, C.P. Altererythrobacter estronivorus sp. nov., an estrogen-degrading strain isolated from Yundang Lagoon of Xiamen City in China. Curr. Microbiol. 2016, 72, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Chen, Y.L.; Wei, S.T.S.; Wu, K.; Lee, T.H.; Wu, T.Y.; Chiang, Y.R. Retroconversion of estrogens into androgens by bacteria via a cobalamin-mediated methylation. Proc. Natl. Acad. Sci. USA 2020, 117, 1395–1403. [Google Scholar] [CrossRef]
- Yu, C.P.; Deeb, R.A.; Chu, K.H. Microbial degradation of steroidal estrogens. Chemosphere 2013, 91, 1225–1235. [Google Scholar] [CrossRef]
- Wang, P.H.; Yu, C.P.; Lee, T.H.; Lin, C.W.; Ismail, W.; Wey, S.P.; Kuo, A.T.; Chiang, Y.R. Anoxic androgen degradation by the denitrifying bacterium Sterolibacterium denitrificans via the 2,3-seco pathway. Appl. Environ. Microbiol. 2014, 80, 3442–3452. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zheng, D.; Wang, Y.; Liang, R. One 3-oxoacyl-(acyl-Carrier-protein) reductase functions as 17β-hydroxysteroid dehydrogenase in the estrogen-degrading Pseudomonas putida SJTE-1. Biochem. Biophys. Res. Commun. 2018, 505, 910–916. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, D.; Peng, W.; Wang, Y.; Wang, X.; Xiong, W.; Liang, R. Characterization of 17β-hydroxysteroid dehydrogenase and regulators involved in estrogen degradation in Pseudomonas putida SJTE-1. Appl. Microbiol. Biotechnol. 2019, 103, 2413–2425. [Google Scholar] [CrossRef]
- Fujii, K.; Satomi, M.; Morita, N.; Motomura, T.; Tanaka, T.; Kikuchi, S. Novosphingobium tardaugens sp. nov., an oestradiol-degrading bacterium isolated from activated sludge of a sewage treatment plant in Tokyo. Int. J. Syst. Evol. Microbiol. 2003, 53, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Sanz, D.; Galán, B.; Díaz, E.; JL, G. High-quality whole-genome sequence of an estradiol-degrading strain, Novosphingobium tardaugens NBRC 16725. Microb. Resour. Annoucements 2019, 8, e01715-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibero, J.; Galán, B.; Díaz, E.; García, J.L. Testosterone degradative pathway of Novosphingobium tardaugens. Genes 2019, 10, 871. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Galán, B.; Rivero-Buceta, V.; García, J.L. Unraveling the 17β-estradiol degradation pathway in Novosphingobium tardaugens NBRC 16725. Front. Microbiol. 2020, 11, 588300. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Cuthbertson, L.; Nodwell, J.R. The TetR family of regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [Green Version]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Schafer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Fernández, H.; Prandoni, N.; Fernández-Pascual, M.; Fajardo, S.; Morcillo, C.; Díaz, E.; Carmona, M. Azoarcus sp. CIB, an anaerobic biodegrader of aromatic compounds shows an endophytic lifestyle. PLoS ONE 2014, 9, e110771. [Google Scholar] [CrossRef]
- Silva-Rocha, R.; Martínez-García, E.; Calles, B.; Chavarría, M.; Arce-Rodríguez, A.; De Las Heras, A.; Páez-Espino, A.D.; Durante-Rodríguez, G.; Kim, J.; Nikel, P.I.; et al. The Standard European Vector Architecture (SEVA): A coherent platform for the analysis and deployment of complex prokaryotic phenotypes. Nucleic Acids Res. 2013, 41, D666–D675. [Google Scholar] [CrossRef]
- Wirth, R.; Friesenegger, A.; Fiedler, S. Transformation of various species of gram-negative bacteria belonging to 11 different genera by electroporation. Mol. Gen. Genet. MGG 1989, 216, 175–177. [Google Scholar] [CrossRef]
- Herrero, M.; De Lorenzo, V.; Timmis, K.N. Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram-negative bacteria. J. Bacteriol. 1990, 172, 6557–6567. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Signer, E.R. Temporal and spatial regulation of the symbiotic genes of Rhizobium meliloti in planta revealed by transposon Tn5-gusA. Genes Dev. 1990, 4, 344–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovyev, V.; Salamov, A. Automatic annotation of microbial genomes and metagenomic sequences. Metagenomics Appl. Agric. Biomed. Environ. Stud. 2011, 62–78. [Google Scholar]
- Coppens, L.; Lavigne, R. SAPPHIRE: A neural network based classifier for σ70 promoter prediction in Pseudomonas. BMC Bioinform. 2020, 21, 415. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Abio Madeira, F.; Mi Park, Y.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, M.; Court, D. Regulatory sequences involved in the promotion and termination of RNA transcription. Annu. Rev. Genet. 1979, 13, 319–353. [Google Scholar] [CrossRef]
- Mutalik, V.K.; Guimaraes, J.C.; Cambray, G.; Lam, C.; Christoffersen, M.J.; Mai, Q.A.; Tran, A.B.; Paull, M.; Keasling, J.D.; Arkin, A.P.; et al. Precise and reliable gene expression via standard transcription and translation initiation elements. Nat. Methods 2013, 10, 354–360. [Google Scholar] [CrossRef]
- Landgraf, D. Quantifying Localizations and Dynamics in Single Bacterial Cells. Ph.D. Thesis, Harvard University, Cambridge, UK, 2012. [Google Scholar]
- Kendall, S.L.; Withers, M.; Soffair, C.N.; Moreland, N.J.; Gurcha, S.; Sidders, B.; Frita, R.; Ten Bokum, A.; Besra, G.S.; Lott, J.S.; et al. A highly conserved transcriptional repressor controls a large regulon involved in lipid degradation in Mycobacterium smegmatis and Mycobacterium tuberculosis. Mol. Microbiol. 2007, 65, 684–699. [Google Scholar] [CrossRef] [Green Version]
- Uhía, I.; Galán, B.; Kendall, S.L.; Stoker, N.G.; García, J.L. Cholesterol metabolism in Mycobacterium smegmatis. Environ. Microbiol. Rep. 2012, 4, 168–182. [Google Scholar] [CrossRef]
- Uhía, I.; Galán, B.; Medrano, F.J.; García, J.L. Characterization of the KstR-dependent promoter of the gene for the first step of the cholesterol degradative pathway in Mycobacterium smegmatis. Microbiology 2011, 157, 2670–2680. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, E.; Medrano, F.J.; Galán, B.; García, J.L. Deciphering the transcriptional regulation of cholesterol catabolic pathway in mycobacteria: Identification of the inducer of KstR repressor. J. Biol. Chem. 2014, 289, 17576–17588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Fernández, J.; Galán, B.; Medrano, F.J.; García, J.L. Characterization of the KstR2 regulator responsible of the lower cholesterol degradative pathway in Mycobacterium smegmatis. Environ. Microbiol. Rep. 2015, 7, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Kendall, S.L.; Burgess, P.; Balhana, R.; Withers, M.; Ten Bokum, A.; Lott, J.S.; Gao, C.; Uhia-Castro, I.; Stoker, N.G. Cholesterol utilization in mycobacteria is controlled by two TetR-type transcriptional regulators: kstR and kstR2. Microbiology 2010, 156, 1362–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.Y.; Shiu, J.H.; Chen, W.M.; Chiang, Y.R.; Tang, S.L. Genomic insight into the host-endosymbiont relationship of Endozoicomonas montiporae CL-33T with its coral host. Front. Microbiol. 2016, 7, 251. [Google Scholar] [CrossRef] [Green Version]
- Casabon, I.; Zhu, S.H.; Otani, H.; Liu, J.; Mohn, W.W.; Eltis, L.D. Regulation of the KstR2 regulon of Mycobacterium tuberculosis by a cholesterol catabolite. Mol. Microbiol. 2013, 89, 1201–1212. [Google Scholar] [CrossRef]
- Linares, M.; Pruneda-Paz, J.L.; Reyna, L.; Genti-Raimondi, S. Regulation of testosterone degradation in Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2008, 112, 145–150. [Google Scholar] [CrossRef]
- Pruneda-Paz, J.L.; Linares, M.; Cabrera, J.E.; Genti-Raimondi, S. Identification of a novel steroid inducible gene associated with the βhsd locus of Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2004, 88, 91–100. [Google Scholar] [CrossRef]
- Pruneda-Paz, J.L.; Linares, M.; Cabrera, J.E.; Genti-Raimondi, S. TeiR, a LuxR-type transcription factor required for testosterone degradation in Comamonas testosteroni. J. Bacteriol. 2004, 186, 1430–1437. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, M.; Kurita, T.; Yamamoto, T.; Hatori, E.; Hayashi, T.; Kudo, T. Steroid degradation gene cluster of Comamonas testosteroni consisting of 18 putative genes from meta-cleavage enzyme gene tesB to regulator gene tesR. Biochem. Biophys. Res. Commun. 2004, 324, 597–604. [Google Scholar] [CrossRef]
- Göhler, A.; Xiong, G.; Paulsen, S.; Trentmann, G.; Maser, E. Testosterone-inducible regulator is a kinase that drives steroid sensing and metabolism in Comamonas testosteroni. J. Biol. Chem. 2008, 283, 17380–17390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.; Maser, E. Regulation of the steroid-inducible 3α-hydroxysteroid dehydrogenase/carbonyl reductase gene in Comamonas testosteroni. J. Biol. Chem. 2001, 276, 9961–9970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.; Martin, H.J.; Maser, E. Identification and characterization of a novel translational repressor of the steroid-inducible 3α-hydroxysteroid dehydrogenase/carbonyl reductase gene in Comamonas testosteroni. J. Biol. Chem. 2003, 278, 47400–47407. [Google Scholar] [CrossRef] [Green Version]
- Gong, W.; Xiong, G.; Maser, E. Identification and characterization of the LysR-type transcriptional regulator HsdR for steroid-inducible expression of the 3α-hydroxysteroid dehydrogenase/carbonyl reductase gene in Comamonas testosteroni. Appl. Environ. Microbiol. 2012, 78, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Pan, T.; Zhang, Y.; Xiong, G.; Yu, Y. Functional analysis of a novel repressor LuxR in Comamonas testosteroni. Chem. Biol. Interact. 2017, 276, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Yang, J.; Gao, L.; Xiong, G.; Yu, Y.; Zhang, Y. Characterization of a LuxR repressor for 3,17β-HSD in Comamonas testosteroni ATCC11996. Chem. Biol. Interact. 2021, 336, 109271. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xiong, G.; Maser, E. A novel transcriptional repressor PhaR for the steroid-inducible expression of the 3,17β-hydroxysteroid dehydrogenase gene in Comamonas testosteroni ATCC 11996. In Proceedings of the Chemico-Biological Interactions; Elsevier: Amsterdam, The Netherlands, 2013; Volume 202, pp. 116–125. [Google Scholar]
- Pan, T.; Huang, P.; Xiong, G.; Maser, E. Isolation and identification of a repressor TetR for 3,17β-HSD expressional regulation in Comamonas testosteroni. Chem. Biol. Interact. 2015, 234, 205–212. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, P.; Xiong, G.; Maser, E. Identification and isolation of a regulator protein for 3,17β-HSD expressional regulation in Comamonas testosteroni. Chem. Biol. Interact. 2015, 234, 197–204. [Google Scholar] [CrossRef]
- Gong, W.; Xiong, G.; Maser, E. Cloning, expression and characterization of a novel short-chain dehydrogenase/reductase (SDRx) in Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 129, 15–21. [Google Scholar] [CrossRef]
- Gong, W.; Xiong, G.; Maser, E. Oligomerization and negative autoregulation of the LysR-type transcriptional regulator HsdR from Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 132, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Shen-Orr, S.S.; Milo, R.; Mangan, S.; Alon, U. Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 2002, 31, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Reichheld, S.E.; Savchenko, A.; Parkinson, J.; Davidson, A.R. A comprehensive analysis of structural and sequence conservation in the TetR family transcriptional regulators. J. Mol. Biol. 2010, 400, 847–864. [Google Scholar] [CrossRef] [PubMed]
- Fahrbach, M.; Kuever, J.; Remesch, M.; Huber, B.E.; Kämpfer, P.; Dott, W.; Hollender, J. Steroidobacter denitrificans gen. nov., sp. nov., a steroidal hormone-degrading gammaproteobacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2215–2223. [Google Scholar] [CrossRef] [Green Version]
- Bertram, R.; Hillen, W. The application of Tet repressor in prokaryotic gene regulation and expression. Microb. Biotechnol. 2008, 1, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Z.; Su, T.; Sun, H.; Zhu, Y.; Qi, Q.; Wang, Q. Tuning the binding affinity of heme-responsive biosensor for precise and dynamic pathway regulation. iScience 2020, 23, 101067. [Google Scholar] [CrossRef]
- Galluzzi, L.; Virtanen, P.; Karp, M. A real-time analysis of QacR-regulated multidrug resistance in Staphylococcus aureus. Biochem. Biophys. Res. Commun. 2003, 301, 24–30. [Google Scholar] [CrossRef]
- Xiong, D.; Lu, S.; Wu, J.; Liang, C.; Wang, W.; Wang, W.; Jin, J.M.; Tang, S.Y. Improving key enzyme activity in phenylpropanoid pathway with a designed biosensor. Metab. Eng. 2017, 40, 115–123. [Google Scholar] [CrossRef]
- Tahlan, K.; Ahn, S.K.; Sing, A.; Bodnaruk, T.D.; Willems, A.R.; Davidson, A.R.; Nodwell, J.R. Initiation of actinorhodin export in Streptomyces coelicolor. Mol. Microbiol. 2007, 63, 951–961. [Google Scholar] [CrossRef]
- Espinosa-Urgel, M.; Serrano, L.; Ramos, J.L.; Fernández-Escamilla, A.M. Engineering biological approaches for detection of toxic compounds: A new microbial biosensor based on the Pseudomonas putida TtgR repressor. Mol. Biotechnol. 2015, 57, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, X.; Xiong, D.; Jin, J.M.; Tang, S.Y. Engineering the effector specificity of regulatory proteins for the in vitro detection of biomarkers and pesticide residues. Appl. Microbiol. Biotechnol. 2019, 103, 3205–3213. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Lopez De Alda, M.J.; Barceló, D. Picogram per liter level determination of estrogens in natural waters and waterworks by a fully automated on-line solid-phase extraction-liquid chromatography-electrospray tandem mass spectrometry method. Anal. Chem. 2005, 76, 6998–7006. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mozaz, S.; Lopez de Alda, M.J.; Barceló, D. Advantages and limitations of on-line solid phase extraction coupled to liquid chromatography-mass spectrometry technologies versus biosensors for monitoring of emerging contaminants in water. J. Chromatogr. A 2007, 1152, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Padeste, C.; Grubelnik, A.; Tiefenauer, L. Amperometric immunosensing using microperoxidase MP-11 antibody conjugates. Anal. Chim. Acta 1998, 374, 167–176. [Google Scholar] [CrossRef]
- Sesay, A.M.; Cullen, D.C. Detection of hormone mimics in water using a miniturised SPR sensor. Environ. Monit. Assess. 2001, 70, 83–92. [Google Scholar] [CrossRef]
- Sun, M.; Du, L.; Gao, S.; Bao, Y.; Wang, S. Determination of 17β-oestradiol by fluorescence immunoassay with streptavidin-conjugated quantum dots as label. Steroids 2010, 75, 400–403. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Zhang, J.; Feng, H.; Liu, X.; Wong, D.K.Y. Detection of estradiol at an electrochemical immunosensor with a Cu UPD|DTBP-Protein G scaffold. Biosens. Bioelectron. 2012, 35, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, I.; López-Montero, J.; Moreno-Guzmán, M.; Janegitz, B.C.; González-Cortés, A.; Yáñez-Sedeño, P.; Pingarrón, J.M. Electrochemical immunosensor for rapid and sensitive determination of estradiol. Anal. Chim. Acta 2012, 743, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Du, B.; Li, H.; Xin, X.; Ma, H.; Wu, D.; Yan, L.; Wei, Q. Metal ions-based immunosensor for simultaneous determination of estradiol and diethylstilbestrol. Biosens. Bioelectron. 2014, 52, 225–231. [Google Scholar] [CrossRef]
- Tomizawa, S.; Chuah, J.A.; Matsumoto, K.; Doi, Y.; Numata, K. Understanding the limitations in the biosynthesis of polyhydroxyalkanoate (PHA) from lignin derivatives. ACS Sustain. Chem. Eng. 2014, 2, 1106–1113. [Google Scholar] [CrossRef]
- Monerris, M.J.; Arévalo, F.J.; Fernández, H.; Zon, M.A.; Molina, P.G. Development of a very sensitive electrochemical immunosensor for the determination of 17β-estradiol in bovine serum samples. Sens. Actuators B Chem. 2015, 208, 525–531. [Google Scholar] [CrossRef]
- Du, L.; Ji, W.; Zhang, Y.; Zhang, C.; Liu, G.; Wang, S. Ultrasensitive detection of 17β-estradiol using gold nanoparticle-based fluorescence immunoassay. J. Mater. Chem. C 2015, 3, 10715–10722. [Google Scholar] [CrossRef] [PubMed]
- Piro, B.; Shi, S.; Reisberg, S.; Noël, V.; Anquetin, G. Comparison of electrochemical immunosensors and aptasensors for detection of small organic molecules in environment, food safety, clinical and public security. Biosensors 2016, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhuang, H.; Du, L.; Lin, S.; Wang, C. Determination of estradiol by biotin-avidin-amplified electrochemical enzyme immunoassay. Anal. Lett. 2007, 40, 887–896. [Google Scholar] [CrossRef]
- Xin, T.B.; Chen, H.; Lin, Z.; Liang, S.X.; Lin, J.M. A secondary antibody format chemiluminescence immunoassay for the determination of estradiol in human serum. Talanta 2010, 82, 1472–1477. [Google Scholar] [CrossRef]
- Kuramitz, H.; Matsuda, M.; Thomas, J.H.; Sugawara, K.; Tanaka, S. Electrochemical immunoassay at a 17β-estradiol self-assembled monolayer electrode using a redox marker. Analyst 2003, 128, 182–186. [Google Scholar] [CrossRef]
- Wang, S.; Lin, S.; Du, L.; Zhuang, H. Flow injection chemiluminescence immunoassay for 17β-estradiol using an immunoaffinity column. Anal. Bioanal. Chem. 2006, 384, 1186–1190. [Google Scholar] [CrossRef]
- Seifert, M. Luminescent enzyme-linked receptor assay for estrogenic compounds. Anal. Bioanal. Chem. 2004, 378, 684–687. [Google Scholar] [CrossRef]
- Habauzit, D.; Chopineau, J.; Roig, B. SPR-based biosensors: A tool for biodetection of hormonal compounds. Anal. Bioanal. Chem. 2007, 387, 1215–1223. [Google Scholar] [CrossRef]
- Tschmelak, J.; Proll, G.; Gauglitz, G. Immunosensor for estrone with an equal limit of detection as common analytical methods. Anal. Bioanal. Chem. 2004, 378, 744–745. [Google Scholar] [CrossRef] [PubMed]
- Hua, P.; Hole, J.P.; Wilkinson, J.S.; Proll, G.; Tschmelak, J.; Gauglitz, G.; Jackson, M.A.; Nudd, R.; Griffith, H.M.T.; Abuknesha, R.A.; et al. Integrated optical fluorescence multisensor for water pollution. Opt. Express 2005, 13, 1124. [Google Scholar] [CrossRef]
- Miyashita, M.; Shimada, T.; Miyagawa, H.; Akamatsu, M. Surface plasmon resonance-based immunoassay for 17β-estradiol and its application to the measurement of estrogen receptor-binding activity. Anal. Bioanal. Chem. 2005, 381, 667–673. [Google Scholar] [CrossRef]
- Pemberton, R.M.; Mottram, T.T.; Hart, J.P. Development of a screen-printed carbon electrochemical immunosensor for picomolar concentrations of estradiol in human serum extracts. J. Biochem. Biophys. Methods 2005, 63, 201–212. [Google Scholar] [CrossRef]
- Butler, D.; Guilbault, G.G. Disposable amperometric immunosensor for the detection of 17-β estradiol using screen-printed electrodes. Sensors Actuators, B Chem. 2006, 113, 692–699. [Google Scholar] [CrossRef]
- Liu, X.; Wong, D.K.Y. Picogram-detection of estradiol at an electrochemical immunosensor with a gold nanoparticle|Protein G-(LC-SPDP)-scaffold. Talanta 2009, 77, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Martínez, N.A.; Schneider, R.J.; Messina, G.A.; Raba, J. Modified paramagnetic beads in a microfluidic system for the determination of ethinylestradiol (EE2) in river water samples. Biosens. Bioelectron. 2010, 25, 1376–1381. [Google Scholar] [CrossRef]
- Olowu, R.A.; Arotiba, O.; Mailu, S.N.; Waryo, T.T.; Baker, P.; Iwuoha, E. Electrochemical aptasensor for endocrine disrupting 17β-estradiol based on a Poly(3,4-ethylenedioxylthiopene)-gold nanocomposite platform. Sensors 2010, 10, 9872–9890. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Chen, L.; Zhang, G.; Liu, Q.; Qiu, B.; Cai, Z.; Chen, G. Label-free aptamer-based electrochemical impedance biosensor for 17β-estradiol. Analyst 2012, 137, 819–822. [Google Scholar] [CrossRef]
- Chaisuwan, N.; Xu, H.; Wu, G.; Liu, J. A highly sensitive differential pulse anodic stripping voltammetry for determination of 17Β-estradiol (E2) using CdSe quantum dots based on indirect competitive immunoassay. Biosens. Bioelectron. 2013, 46, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Kanso, H.; Barthelmebs, L.; Inguimbert, N.; Noguer, T. Immunosensors for estradiol and ethinylestradiol based on new synthetic estrogen derivatives: Application to wastewater analysis. Anal. Chem. 2013, 85, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Liu, M.; Zhuang, L.; Li, Z.; Fan, L.; Zhao, G. A Fetomolar level 17β-estradiol electrochemical aptasensor constructed on hierachical dendritic gold modified boron-doped diamond electrode. Electrochim. Acta 2014, 137, 146–153. [Google Scholar] [CrossRef]
- Huang, K.J.; Liu, Y.J.; Shi, G.W.; Yang, X.R.; Liu, Y.M. Label-free aptamer sensor for 17β-estradiol based on vanadium disulfide nanoflowers and Au nanoparticles. Sensors Actuators, B Chem. 2014, 201, 579–585. [Google Scholar] [CrossRef]
- Huang, K.J.; Liu, Y.J.; Zhang, J.Z.; Liu, Y.M. A novel aptamer sensor based on layered tungsten disulfide nanosheets and Au nanoparticles amplification for 17β-estradiol detection. Anal. Methods 2014, 6, 8011–8017. [Google Scholar] [CrossRef]
- Zhu, B.; Alsager, O.A.; Kumar, S.; Hodgkiss, J.M.; Travas-Sejdic, J. Label-free electrochemical aptasensor for femtomolar detection of 17β-estradiol. Biosens. Bioelectron. 2015, 70, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Zhao, G.; Shi, H.; Liu, M. A simple and label-free aptasensor based on nickel hexacyanoferrate nanoparticles as signal probe for highly sensitive detection of 17β-estradiol. Biosens. Bioelectron. 2015, 68, 303–309. [Google Scholar] [CrossRef]
- Seifert, M.; Haindl, S.; Hock, B. Development of an enzyme linked receptor assay (ELRA) for estrogens and xenoestrogens. Anal. Chim. Acta 1999, 386, 191–199. [Google Scholar] [CrossRef]
- Usami, M.; Mitsunaga, K.; Ohno, Y. Estrogen receptor binding assay of chemicals with a surface plasmon resonance biosensor. J. Steroid Biochem. Mol. Biol. 2002, 81, 47–55. [Google Scholar] [CrossRef]
- Rich, R.L.; Hoth, L.R.; Geoghegan, K.F.; Brown, T.A.; Lemotte, P.K.; Simons, S.P.; Hensley, P.; Myszka, D.G. Kinetic analysis of estrogen receptor/ligand interactions. Proc. Natl. Acad. Sci. USA 2002, 99, 8562–8567. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.K.; Li, J.; Im, J.E.; Ahn, K.S.; Park, T.S.; Cho, S.I.; Kim, Y.R.; Lee, W.Y. Impedometric estrogen biosensor based on estrogen receptor alpha-immobilized gold electrode. J. Electroanal. Chem. 2012, 671, 106–111. [Google Scholar] [CrossRef]
- Pearson, J.; Gill, A.; Margison, G.P.; Vadgama, P.; Povey, A.C. A rapid receptor-ligand assay determination of estrogens using surface plasmon resonance. In Proceedings of the Sensors and Actuators, B: Chemical; Elsevier: Amsterdam, The Netherlands, 2001; Volume 76, pp. 1–7. [Google Scholar]
- La Spina, R.; Ferrero, V.E.V.; Aiello, V.; Pedotti, M.; Varani, L.; Lettieri, T.; Calzolai, L.; Haasnoot, W.; Colpo, P. Label-free biosensor detection of endocrine disrupting compounds using engineered estrogen receptors. Biosensors 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Furst, A.L.; Hoepker, A.C.; Francis, M.B. Quantifying hormone disruptors with an engineered bacterial biosensor. ACS Cent. Sci. 2017, 3, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Gierach, I.; Shapero, K.; Eyster, T.W.; Wood, D.W. Bacterial biosensors for evaluating potential impacts of estrogenic endocrine disrupting compounds in multiple species. Environ. Toxicol. 2013, 28, 179–189. [Google Scholar] [CrossRef]
- Liang, R.; Zhou, J.; Liu, J. Construction of a bacterial assay for estrogen detection based on an estrogen-sensitive intein. Appl. Environ. Microbiol. 2011, 77, 2488–2495. [Google Scholar] [CrossRef] [Green Version]
- Zutz, C.; Wagener, K.; Yankova, D.; Eder, S.; Möstl, E.; Drillich, M.; Rychli, K.; Wagner, M.; Strauss, J. A robust high-throughput fungal biosensor assay for the detection of estrogen activity. Steroids 2017, 126, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Wozei, E.; Hermanowicz, S.W.; Holman, H.Y.N. Developing a biosensor for estrogens in water samples: Study of the real-time response of live cells of the estrogen-sensitive yeast strain RMY/ER-ERE using fluorescence microscopy. Biosens. Bioelectron. 2006, 21, 1654–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, T.; Tag, K.; Riedel, K.; Uhlig, S.; Baronian, K.; Gellissen, G.; Kunze, G. A novel estrogen sensor based on recombinant Arxula adeninivorans cells. Biosens. Bioelectron. 2006, 21, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Fine, T.; Leskinen, P.; Isobe, T.; Shiraishi, H.; Morita, M.; Marks, R.S.; Virta, M. Luminescent yeast cells entrapped in hydrogels for estrogenic endocrine disrupting chemical biodetection. Biosens. Bioelectron. 2006, 21, 2263–2269. [Google Scholar] [CrossRef]
- Awais, M.; Sato, M.; Sasaki, K.; Umezawa, Y. A genetically encoded fluorescent indicator capable of discriminating estrogen agonists from antagonists in living cells. Anal. Chem. 2004, 76, 2181–2186. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.M.; Sonnenschein, C.; Chung, K.L.; Fernandez, M.F.; Olea, N.; Olea Serrano, F. The E-SCREEN assay as a tool to identify estrogens: An update on estrogenic environmental pollutants. In Proceedings of the Environmental Health Perspectives; Public Health Services, US Department of Health and Human Services: Boston, MA, USA, 1995; Volume 103, pp. 113–122. [Google Scholar]
- Legler, J.; Van Den Brink, C.E.; Brouwer, A.; Murk, A.J.; Van Der Saag, P.T.; Vethaak, A.D.; Van Der Burg, B. Development of a stably transfected estrogen receptor-mediated luciferase reporter gene assay in the human T47D breast cancer cell line. Toxicol. Sci. 1999, 48, 55–66. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Chua, N.H. An estrogen receptor-based transactivator XVE mediates highly inducible gene expression in transgenic plants. Plant J. 2000, 24, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas-Jones, E.; Thorpe, K.; Harrison, N.; Thomas, G.; Morris, C.; Hutchinson, T.; Woodhead, S.; Tyler, C. Dynamics of estrogen biomarker responses in rainbow trout exposed to 17β-estradiol and 17α-ethinylestradiol. Environ. Toxicol. Chem. 2003, 22, 3001–3008. [Google Scholar] [CrossRef]
- Lee, O.; Green, J.M.; Tyler, C.R. Transgenic fish systems and their application in ecotoxicology. Crit. Rev. Toxicol. 2015, 45, 124–141. [Google Scholar] [CrossRef]
- Kurauchi, K.; Nakaguchi, Y.; Tsutsumi, M.; Hori, H.; Kurihara, R.; Hashimoto, S.; Ohnuma, R.; Yamamoto, Y.; Matsuoka, S.; Kawai, S.; et al. In vivo visual reporter system for detection of estrogen-like substances by transgenic medaka. Environ. Sci. Technol. 2005, 39, 2762–2768. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Shan, T.; Tong, Y.; Lam, S.H.; Gong, Z. Development of estrogen-responsive transgenic medaka for environmental monitoring of endocrine disrupters. Environ. Sci. Technol. 2005, 39, 9001–9008. [Google Scholar] [CrossRef]
- Salam, M.A.; Sawada, T.; Ohya, T.; Ninomiya, K.; Hayashi, S. Detection of environmental estrogenicity using transgenic medaka hatchlings (Oryzias latipes) expressing the GFP-tagged choriogenin L Gene. J. Environ. Sci. Heal. Part A Toxic/Hazardous Subst. Environ. Eng. 2008, 43, 272–277. [Google Scholar] [CrossRef]
- Cho, Y.S.; Kim, D.S.; Nam, Y.K. Characterization of estrogen-responsive transgenic marine medaka Oryzias dancena germlines harboring red fluorescent protein gene under the control by endogenous choriogenin H promoter. Transgenic Res. 2013, 22, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Legler, J.; Broekhof, J.L.M.; Brouwer, A.; Lanser, P.H.; Murk, A.J.; Van Der Saag, P.T.; Dick Vethaak, A.; Wester, P.; Zivkovic, D.; Van Der Burg, B. A novel in vivo bioassay for (xeno-)estrogens using transgenic zebrafish. Environ. Sci. Technol. 2000, 34, 4439–4444. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hu, J.; Yang, J.; Wang, Y.; Xu, H.; Jiang, Q.; Gong, Y.; Gu, Y.; Song, H. Generation of a fluorescent transgenic zebrafish for detection of environmental estrogens. Aquat. Toxicol. 2010, 96, 53–61. [Google Scholar] [CrossRef]
- Brion, F.; Le Page, Y.; Piccini, B.; Cardoso, O.; Tong, S.K.; Chung, B.C.; Kah, O. Screening estrogenic activities of chemicals or mixtures in vivo using transgenic (cyp19a1b-GFP) zebrafish embryos. PLoS ONE 2012, 7, e36069. [Google Scholar] [CrossRef] [Green Version]
- Gorelick, D.A.; Iwanowicz, L.R.; Hung, A.L.; Blazer, V.S.; Halpern, M.E. Transgenic zebrafish reveal tissue-specific differences in estrogen signaling in response to environmental water samples. Environ. Health Perspect. 2014, 122, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Luo, Y.; Jin, S.; Maser, E. Cis- and trans-regulatory elements of 3α-hydroxysteroid dehydrogenase/carbonyl reductase as biosensor system for steroid determination in the environment. Chem. Biol. Interact. 2009, 178, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Maser, E.; Xiong, G. The Comamonas testosteroni steroid biosensor system (COSS)-Reflection on other methods. J. Steroid Biochem. Mol. Biol. 2010, 121, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Grazon, C.; Baer, R.C.; Kuzmanović, U.; Nguyen, T.; Chen, M.; Zamani, M.; Chern, M.; Aquino, P.; Zhang, X.; Lecommandoux, S.; et al. A progesterone biosensor derived from microbial screening. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Genotype and Characteristics | Source/Reference |
|---|---|---|
| C. tardaugens | ||
| NBRC 16725 | wild type strain (ARI-1) | [20] |
| RfR | RfR strain efficient for conjugation | [22] |
| ΔedcA | C. tardaugens NBRC 16725 ΔEGO55_13525 | [23] |
| ΔedcR | C. tardaugens NBRC 16725 ΔEGO55_13520 | This study |
| E. coli | ||
| DH10B | F−, mcrA, Δ(mrr hsdRMS-mcrBC), Φ80dlacZΔM15, ΔlacX74, deoR, recA1, araD139, Δ(ara-leu)7697, galU, galK, λ−, rpsL, endA1, nupG | Invitrogen |
| HB101 | supE44, ara14, galK2, leuB, lacY1, ∆(gpt-proA)62, rpsL20, xyl-5, mtl-1, recA13, ∆(mcrC-mrr), hsdS20 (rB−mB−), SmR | [27] |
| Plasmids | ||
| pK18mobsacB | KmR, ColE oriV, Mob+, lacZα, sacB; vector for allelic exchange homologous recombination mutagenesis | [28] |
| pK18edcR | pK18mobsacB derivative used for EGO55_13520 deletion | This study |
| pSEVA237PlexA | KmR, oriV (pBBR1), constituve expression of gfp gene under the control of the PlexA promoter | [29] |
| pSEVA23PlexA | pSEVA237PlexA where gfp was deleted using XbaI-SpeI restriction enzymes and served as empty vector | [23] |
| pSEVA23edcR | pSEVA237PlexA where gfp gene was replaced by EGO55_13520 | This study |
| pSEVA237Pa | pSEVA237PlexA where PlexA promoter was replaced by Pa | This study |
| pSEVA237Pb | pSEVA237PlexA where PlexA promoter was replaced by Pb | This study |
| pSEVA237Pt | pSEVA237PlexA where PlexA promoter was replaced by Pt | This study |
| pSEVA237M-BCD2-14g | KmR, pSEVA237M derivative, synthetic bicistronic RBS BCD2, P14g promoter | Kindly provided by P. Nikel |
| pSEVA237MPb | KmR, pSEVA237M-BCD2-14g derivative where P14g promoter was replaced by Pb | This study |
| pSEVA651 | GmR, oriV (RSF1010), standard MCS | [30] |
| pSEVA65edcR | pSEVA651 containing EGO55_13520 gene expressed constitutively under PlexA promoter | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibero, J.; Galán, B.; García, J.L. Identification of the EdcR Estrogen-Dependent Repressor in Caenibius tardaugens NBRC 16725: Construction of a Cellular Estradiol Biosensor. Genes 2021, 12, 1846. https://doi.org/10.3390/genes12121846
Ibero J, Galán B, García JL. Identification of the EdcR Estrogen-Dependent Repressor in Caenibius tardaugens NBRC 16725: Construction of a Cellular Estradiol Biosensor. Genes. 2021; 12(12):1846. https://doi.org/10.3390/genes12121846
Chicago/Turabian StyleIbero, Juan, Beatriz Galán, and José L. García. 2021. "Identification of the EdcR Estrogen-Dependent Repressor in Caenibius tardaugens NBRC 16725: Construction of a Cellular Estradiol Biosensor" Genes 12, no. 12: 1846. https://doi.org/10.3390/genes12121846