
Marek’s Disease Virus Telomeric Integration Profiles of Neoplastic Host Tissues Reveal Unbiased Chromosomal Selection and Loss of Cellular Diversity during Tumorigenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Resources and MDV Challenge
2.2. Tumor Collection and Processing
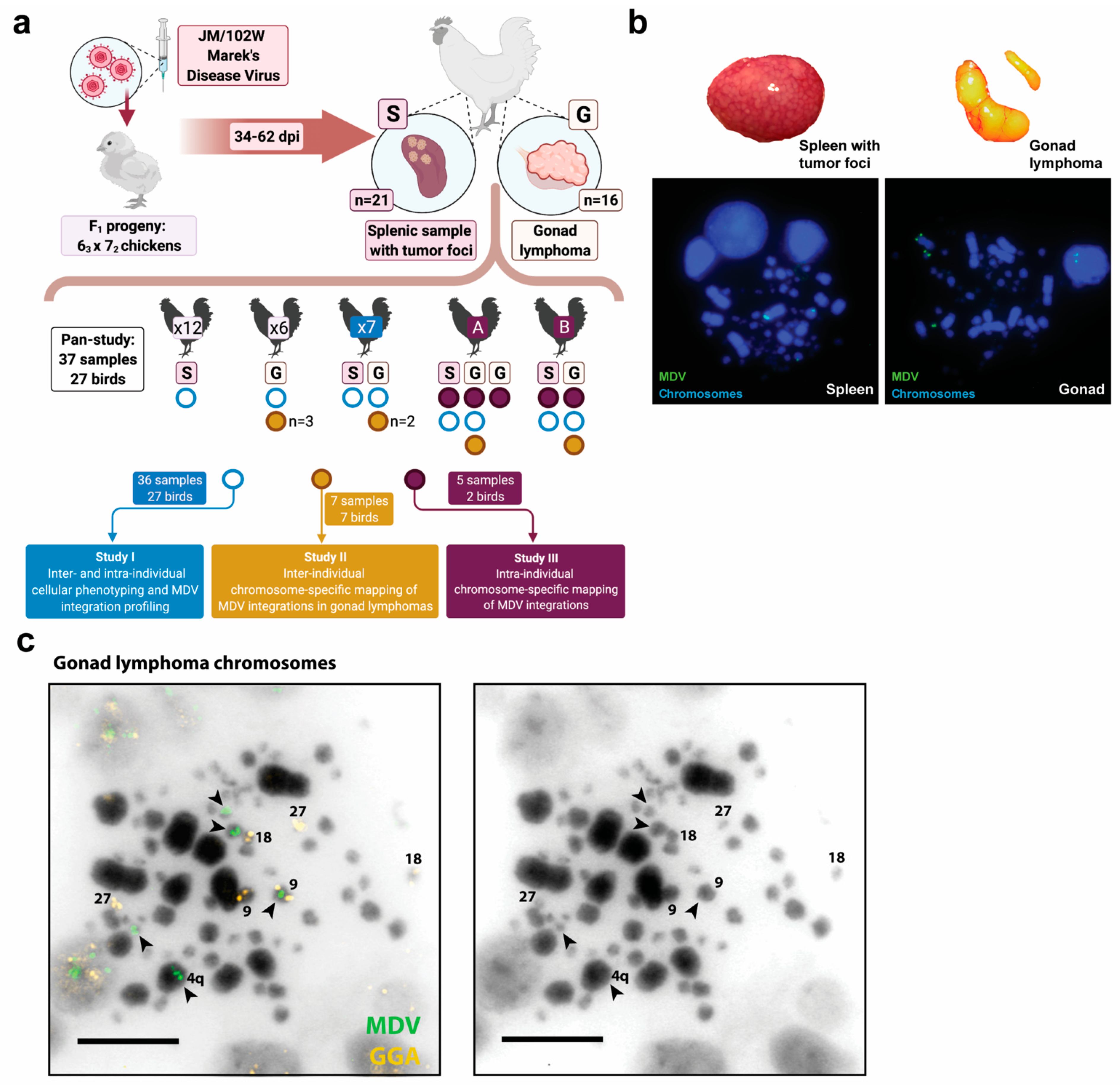
2.3. Study Design
2.4. Cytogenetic Studies and Analyses
2.5. MDV and Chromosome-Specific Probes
2.6. Probe Labeling and Fluorescence In Situ Hybridization (FISH)
2.7. Data Visualization and Statistical Analysis
3. Results
3.1. Study Part I. Inter- and Intra-Individual MDV Cellular Phenotyping and Integration Profiling in MD Lymphomas from Gonadal and Splenic Tissues
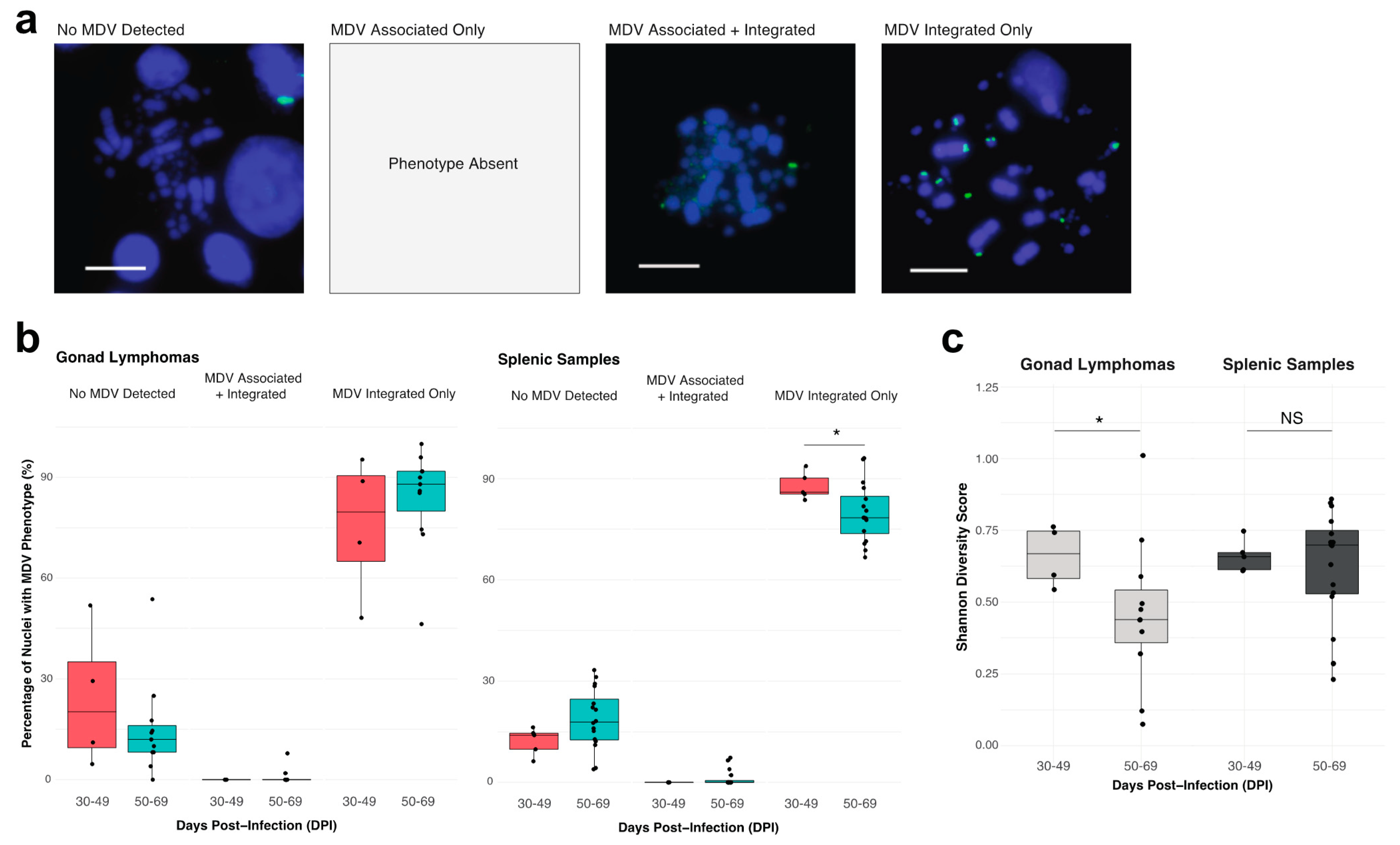
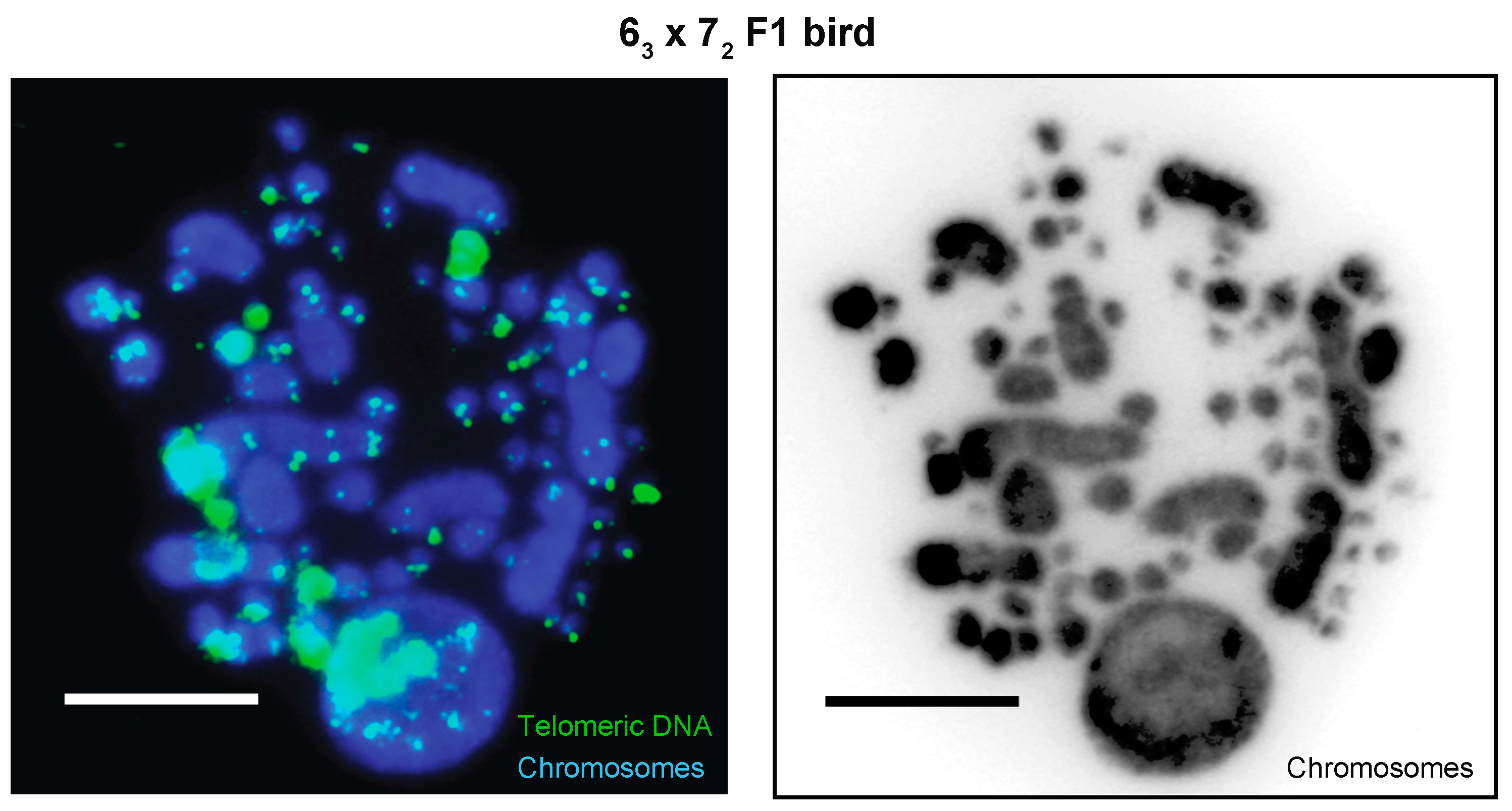
3.1.1. MDV Cellular Phenotyping
3.1.2. MDV Integration Profiling
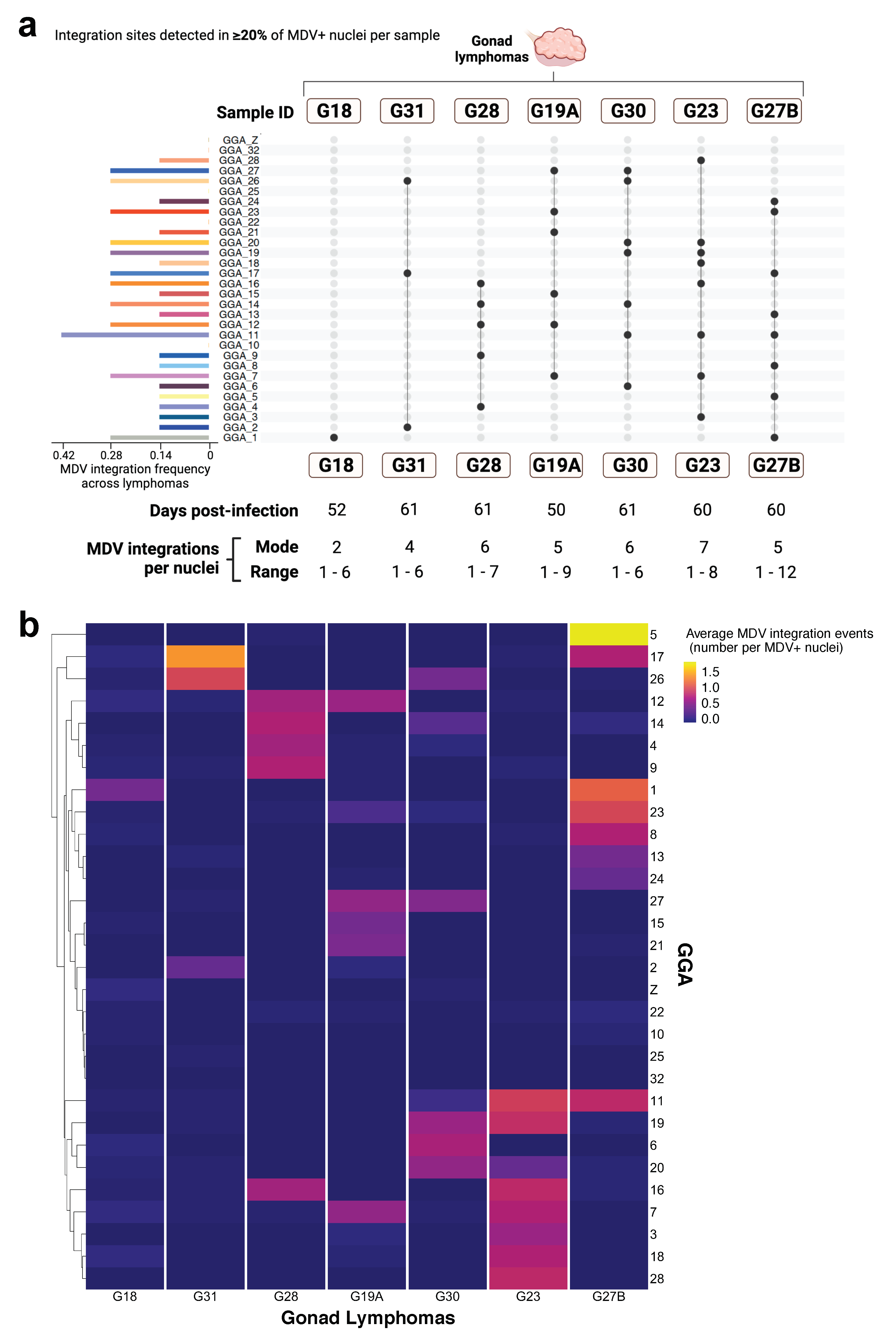
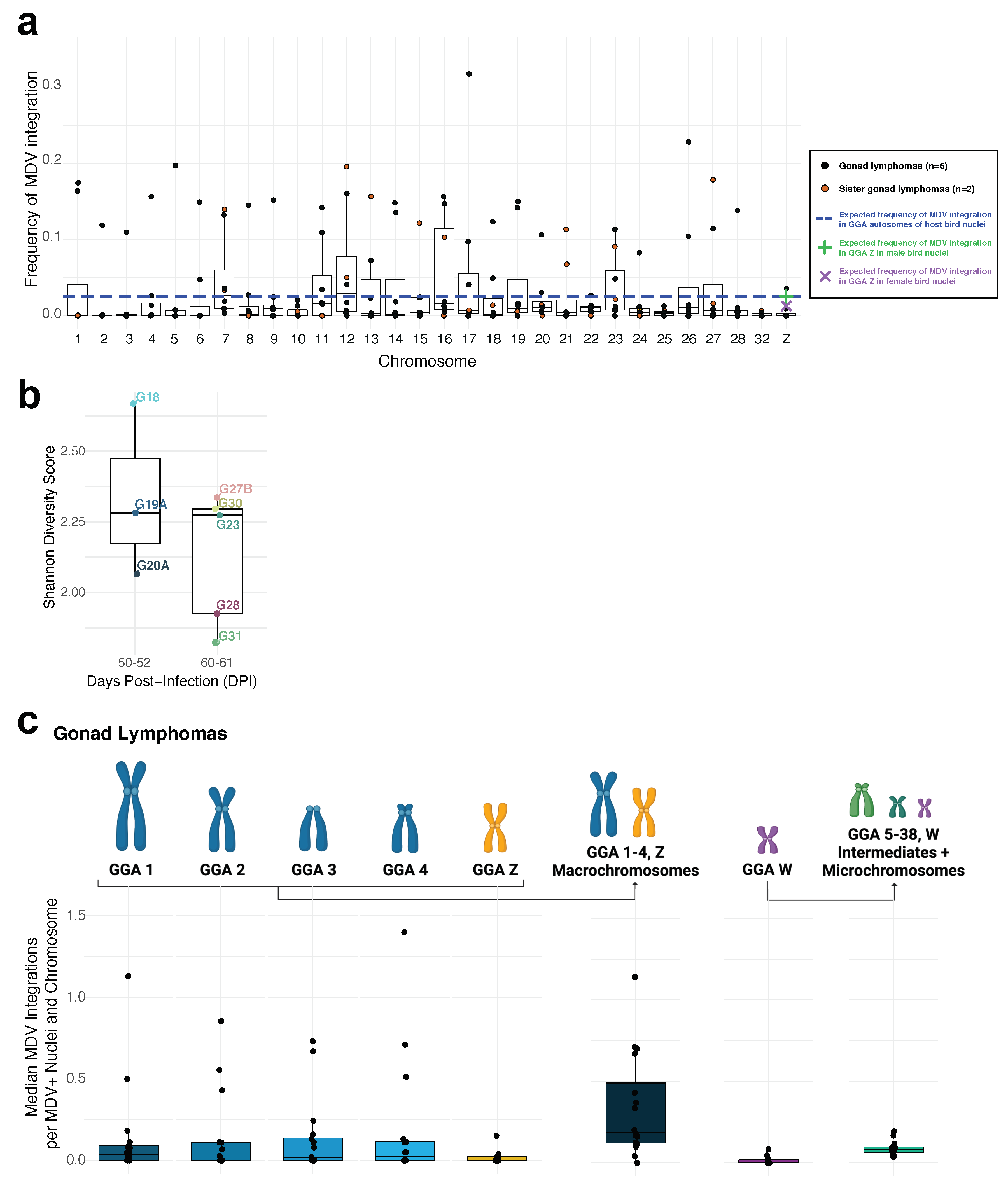
3.2. Study Part II. Inter-Individual Chromosome-Specific Mapping of MDV Integration in Gonad Lymphomas
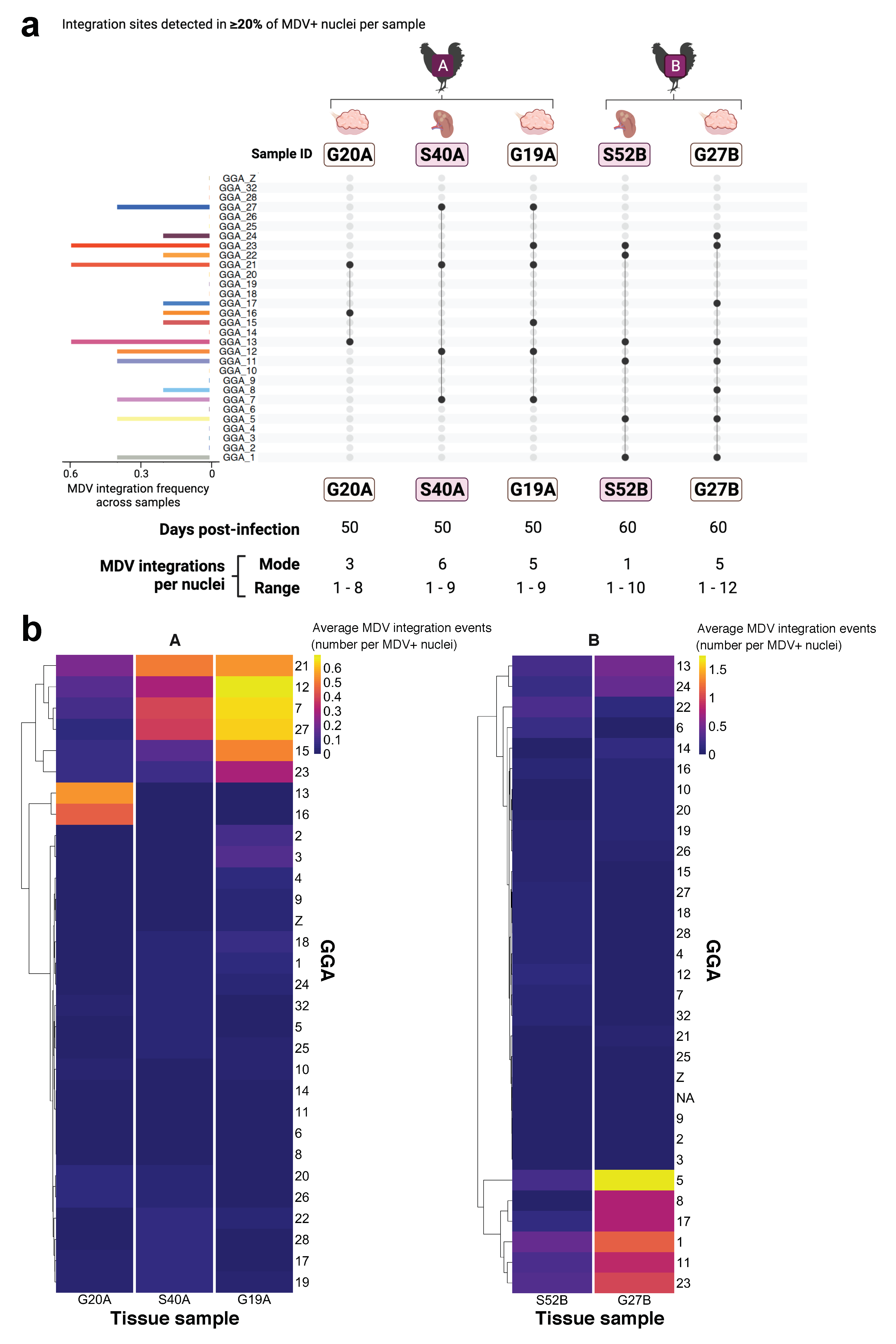
3.3. Study Part III. Intra-Individual Chromosome-Specific Mapping of MDV Integration in Gonad Lymphomas and Splenic Tissue Samples
3.4. Study Parts I–III. Cross-Experiment Analysis of MDV Integration Mapping
4. Discussion
4.1. MD Gonad Lymphomas and Splenic Lymphoproliferative Foci Samples Primarily Consist of Cells with an MDV Telomere-Integrated-Only Phenotype and Demonstrate Unique Temporal Dynamics in MDV Cytogenomic Phenotypes
4.2. MD Gonad Lymphomas Exhibit Significantly Lower Diversity in Terms of MDV Integration Profiles at Later Stages of Tumorigenesis
4.3. MDV FISH Analysis Indicates Heterogenous Inter-Individual MDV Integration Profiles among MD Gonad Lymphomas from Different Host Birds
4.4. MDV FISH Analysis Reveals Similar Intra-Individual MDV Integration Profiles for MDV-Transformed Tissue Samples
4.5. Cross-Experiment Analysis Reveals No Significant MDV Integration ‘Hotspots’ Amongst Host Chromosomes and Lower Diversity of MDV Integration Profiles over Time in MD Gonad Lymphomas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s Disease Virus: From Miasma to Model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef]
- Gimeno, I.M.; Witter, R.L.; Cortes, A.L.; Reed, W.M. Replication Ability of Three Highly Protective Marek’s Disease Vaccines: Implications in Lymphoid Organ Atrophy and Protection. Avian Pathol. 2011, 40, 573–579. [Google Scholar] [CrossRef]
- Baigent, S.J.; Davison, F. Marek’s Disease virus: Biology and life cycle. In Marek’s Disease, an Evolving Problem; Davison, F., Nair, V., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 62–77. ISBN 9780120883790. [Google Scholar]
- Arumugaswami, V.; Kumar, P.M.; Konjufca, V.; Dienglewicz, R.L.; Reddy, S.M.; Parcells, M.S. Latency of Marek’s Disease Virus (MDV) in a Reticuloendotheliosis Virus-Transformed T-Cell Line. II: Expression of the Latent MDV Genome. Avian Dis. 2009, 53, 156–165. [Google Scholar] [CrossRef]
- Adldinger, H.; Calnek, B. Pathogenesis of Marek’s Disease: Early Distribution of Virus and Viral Antigens in Infected Chickens. J. Natl. Cancer Inst. 1973, 50, 1287–1298. [Google Scholar] [CrossRef]
- Biggs, P.M. Marek’s Disease-Current State of Knowledge. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 1968; pp. 92–125. [Google Scholar]
- Robinson, C.M.; Cheng, H.H.; Delany, M.E. Temporal Kinetics of Marek’s Disease Herpesvirus: Integration Occurs Early after Infection in Both B and T Cells. Cytogenet. Genome Res. 2014, 144, 142–154. [Google Scholar] [CrossRef]
- Robinson, C.M.; Hunt, H.D.; Cheng, H.H.; Delany, M.E. Chromosomal Integration of an Avian Oncogenic Herpesvirus Reveals Telomeric Preferences and Evidence for Lymphoma Clonality. Herpesviridae 2010, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Kaschka-Dierich, C.; Nazerian, K.; Thomssen, R. Intracellular State of Marek’s Disease Virus DNA in Two Tumour-Derived Chicken Cell Lines. J. Gen. Virol. 1979, 44, 271–280. [Google Scholar] [CrossRef]
- Delecluse, H.J.; Hammerschmidt, W. Status of Marek’s Disease Virus in Established Lymphoma Cell Lines: Herpesvirus Integration Is Common. J. Virol. 1993, 67, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witter, R.L. A New Strategy for Marek’s Disease Immunisation -Bivalent Vaccine. Avian Pathol. 1984, 13, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calnek, B.W. Pathogenesis of Marek’s Disease Virus Infection. In Marek’s Disease; Hirai, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 25–55. [Google Scholar]
- Burgess, S.C.; Davison, T.F. Identification of the Neoplastically Transformed Cells in Marek’s Disease Herpesvirus-Induced Lymphomas: Recognition by the Monoclonal Antibody AV37. J. Virol. 2002, 76, 7276–7292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, V. Evolution of Marek’s Disease--a Paradigm for Incessant Race between the Pathogen and the Host. Vet. J. 2005, 170, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Trapp, S.; Parcells, M.S.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Kumar, P.M.; Nair, V.K.; Osterrieder, N. A Virus-Encoded Telomerase RNA Promotes Malignant T Cell Lymphomagenesis. J. Exp. Med. 2006, 203, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Calnek, B.W.; Witter, R.L. Marek’s Disease—A Model for Herpesvirus Oncology. CRC Crit. Rev. Microbiol. 1985, 12, 293–320. [Google Scholar] [CrossRef]
- Lee, S.-I.; Ohashi, K.; Morimura, T.; Sugimoto, C.; Onuma, M. Re-Isolation of Marek’s Disease Virus from T Cell Subsets of Vaccinated and Non-Vaccinated Chickens. Arch. Virol. 1999, 144, 45–54. [Google Scholar] [CrossRef]
- Mwangi, W.N.; Smith, L.P.; Baigent, S.J.; Smith, A.L.; Nair, V. Induction of Lymphomas by Inoculation of Marek’s Disease Virus-Derived Lymphoblastoid Cell Lines: Prevention by CVI988 Vaccination. Avian Pathol. 2012, 41, 589–598. [Google Scholar] [CrossRef]
- Schat, K.A.; Chen, C.L.; Calnek, B.W.; Char, D. Transformation of T-Lymphocyte Subsets by Marek’s Disease Herpesvirus. J. Virol. 1991, 65, 1408–1413. [Google Scholar] [CrossRef] [Green Version]
- Calnek, B.W.; Schat, K.A.; Ross, L.J.N.; Shek, W.R.; Chen, C.-L.H. Further Characterization of Marek’s Disease Virus-Infected Lymphocytes. I.In Vivo Infection. Int. J. Cancer 1984, 33, 389–398. [Google Scholar] [CrossRef]
- Jones, D.; Lee, L.; Liu, J.L.; Kung, H.J.; Tillotson, J.K. Marek Disease Virus Encodes a Basic-Leucine Zipper Gene Resembling the Fos/Jun Oncogenes That Is Highly Expressed in Lymphoblastoid Tumors. Proc. Natl. Acad. Sci. USA 1992, 89, 4042–4046. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.C.; Smith, L.P.; Kgosana, L.; Baigent, S.J.; Nair, V.; Allday, M.J. Homodimerization of the Meq Viral Oncoprotein Is Necessary for Induction of T-Cell Lymphoma by Marek’s Disease Virus. J. Virol. 2009, 83, 11142–11151. [Google Scholar] [CrossRef] [Green Version]
- Kaufer, B.B.; Arndt, S.; Trapp, S.; Osterrieder, N.; Jarosinski, K.W. Herpesvirus Telomerase RNA (VTR) with a Mutated Template Sequence Abrogates Herpesvirus-Induced Lymphomagenesis. PLoS Pathog. 2011, 7, e1002333. [Google Scholar] [CrossRef] [PubMed]
- Fragnet, L.; Kut, E.; Rasschaert, D. Comparative Functional Study of the Viral Telomerase RNA Based on Natural Mutations. J. Biol. Chem. 2005, 280, 23502–23515. [Google Scholar] [CrossRef] [Green Version]
- Jarosinski, K.W.; Tischer, B.K.; Trapp, S.; Osterrieder, N. Marek’s Disease Virus: Lytic Replication, Oncogenesis and Control. Expert Rev. Vaccines 2006, 5, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Artandi, S.E.; DePinho, R.A. Telomeres and Telomerase in Cancer. Carcinogenesis 2010, 31, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Zou, Y.; Hiyama, E.; Wright, W.E. Telomerase and Cancer. Hum. Mol. Genet. 2001, 10, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufer, B.B.; Jarosinski, K.W.; Osterrieder, N. Herpesvirus Telomeric Repeats Facilitate Genomic Integration into Host Telomeres and Mobilization of Viral DNA during Reactivation. J. Exp. Med. 2011, 208, 605–615. [Google Scholar] [CrossRef]
- Greco, A.; Fester, N.; Engel, A.T.; Kaufer, B.B. Role of the Short Telomeric Repeat Region in Marek’s Disease Virus Replication, Genomic Integration, and Lymphomagenesis. J. Virol. 2014, 88, 14138–14147. [Google Scholar] [CrossRef] [Green Version]
- McPherson, M.C.; Cheng, H.H.; Delany, M.E. Marek’s Disease Herpesvirus Vaccines Integrate into Chicken Host Chromosomes yet Lack a Virus-Host Phenotype Associated with Oncogenic Transformation. Vaccine 2016, 34, 5554–5561. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.F.; Dunn, J.R.; Cheng, H.H.; Niikura, M. A MEQ-Deleted Marek’s Disease Virus Cloned as a Bacterial Artificial Chromosome Is a Highly Efficacious Vaccine. Avian Dis. 2010, 54, 862–869. [Google Scholar] [CrossRef]
- Morissette, G.; Flamand, L. Herpesviruses and Chromosomal Integration. J. Virol. 2010, 84, 12100–12109. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Wang, Z.; Lieberman, P.M. Telomeres and Viruses: Common Themes of Genome Maintenance. Front. Oncol. 2012, 2, 201. [Google Scholar] [CrossRef] [Green Version]
- Truyen, U.; Löchelt, M. Relevant Oncogenic Viruses in Veterinary Medicine: Original Pathogens and Animal Models for Human Disease. Contrib. Microbiol. 2006, 13, 101–117. [Google Scholar] [CrossRef]
- Delecluse, H.J.; Bartnizke, S.; Hammerschmidt, W.; Bullerdiek, J.; Bornkamm, G.W. Episomal and Integrated Copies of Epstein-Barr Virus Coexist in Burkitt Lymphoma Cell Lines. J. Virol. 1993, 67, 1292–1299. [Google Scholar] [CrossRef] [Green Version]
- Kaschka-Dierich, C.; Adams, A.; Lindahl, T.; Bornkamm, G.W.; Bjursell, G.; Klein, G.; Giovanella, B.C.; Singh, S. Intracellular Forms of Epstein–Barr Virus DNA in Human Tumour Cells in Vivo. Nature 1976, 260, 302–306. [Google Scholar] [CrossRef]
- Daibata, M.; Taguchi, T.; Taguchi, H.; Miyoshi, I. Integration of Human Herpesvirus 6 in a Burkitt’s Lymphoma Cell Line. Br. J. Haematol. 1998, 102, 1307–1313. [Google Scholar] [CrossRef]
- Green, M.; Michaels, M.G. Epstein–Barr Virus Infection and Posttransplant Lymphoproliferative Disorder. Am. J. Transplant. 2013, 13, 41–54. [Google Scholar] [CrossRef]
- Takada, K. Role of Epstein-Barr Virus in Burkitt’s Lymphoma. In Epstein-Barr Virus and Human Cancer; Takada, K., Ed.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2001; pp. 141–151. ISBN 978-3-642-56515-1. [Google Scholar]
- Mesri, E.A.; Feitelson, M.A.; Munger, K. Human Viral Oncogenesis: A Cancer Hallmarks Analysis. Cell Host Microbe 2014, 15, 266–282. [Google Scholar] [CrossRef] [Green Version]
- Shannon-Lowe, C.; Rickinson, A.B.; Bell, A.I. Epstein–Barr Virus-Associated Lymphomas. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160271. [Google Scholar] [CrossRef] [PubMed]
- Ringehan, M.; McKeating, J.A.; Protzer, U. Viral Hepatitis and Liver Cancer. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomples, U.A.; Macaulay, H.A. Characterization of Human Telomeric Repeat Sequences from Human Herpesvirus 6 and Relationship to Replication. J. Gen. Virol. 1995, 76, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Nacheva, E.P.; Ward, K.N.; Brazma, D.; Virgili, A.; Howard, J.; Leong, H.N.; Clark, D.A. Human Herpesvirus 6 Integrates Within Telomeric Regions as Evidenced by Five Different Chromosomal Sites. J. Med. Virol. 2008, 1958, 1952–1958. [Google Scholar] [CrossRef]
- Arbuckle, J.H.; Medveczky, M.M.; Luka, J.; Hadley, S.H.; Luegmayr, A.; Ablashi, D.; Lund, T.C.; Tolar, J.; De Meirleir, K.; Montoya, J.G.; et al. The Latent Human Herpesvirus-6A Genome Specifically Integrates in Telomeres of Human Chromosomes in Vivo and in Vitro. Proc. Natl. Acad. Sci. USA 2010, 107, 5563–5568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, R.-J.; Han, B.-W.; Cai, Q.-Q.; Zuo, X.-Y.; Xia, T.; Chen, J.-R.; Feng, L.-N.; Lim, J.Q.; Chen, S.-W.; Zeng, M.-S.; et al. Genomic and Transcriptomic Landscapes of Epstein-Barr Virus in Extranodal Natural Killer T-Cell Lymphoma. Leukemia 2019, 33, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Suzumiya, J.; Kanda, M.; Kato, A.; Kikuchi, M. Integrated and Episomal Forms of Epstein–Barr Virus (EBV) in EBV Associated Disease. Cancer Lett. 1998, 122, 43–50. [Google Scholar] [CrossRef]
- Kripalani-Joshi, S.; Law, H.Y. Identification of Integrated Epstein-Barr Virus in Nasopharyngeal Carcinoma Using Pulse Field Gel Electrophoresis. Int. J. Cancer 1994, 56, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kost, J.; Sulovari, A.; Wong, N.; Liang, W.; Cao, J.; Li, D. A Virome-Wide Clonal Integration Analysis Platform for Discovering Cancer Viral Etiology. Genome Res. 2019, 29, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, K.; Yu, Z.; Li, X.; Li, X.; Tang, K.; Tu, C.; Qi, P.; Liao, Q.; Chen, P.; Zeng, Z.; et al. Genome-Wide Analysis of Epstein-Barr Virus (EBV) Integration and Strain in C666-1 and Raji Cells. J. Cancer 2016, 7, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravorty, S.; Yan, B.; Wang, C.; Wang, L.; Quaid, J.T.; Lin, C.F.; Briggs, S.D.; Majumder, J.; Canaria, D.A.; Chauss, D.; et al. Integrated Pan-Cancer Map of EBV-Associated Neoplasms Reveals Functional Host–Virus Interactions. Cancer Res. 2019, 79, 6010–6023. [Google Scholar] [CrossRef] [Green Version]
- Desfarges, S.; Ciuffi, A. Viral Integration and Consequences on Host Gene Expression. In Viruses: Essential Agents of Life; Springer: Berlin, Germany, 2012; pp. 147–175. [Google Scholar]
- Schat, K.A. Marek’s Disease: A Model for Protection against Herpesvirus-Induced Tumours. Cancer Surv. 1987, 6, 1–37. [Google Scholar]
- Witter, R.L. Characteristics of Marek’s Disease Viruses Isolated from Vaccinated Commercial Chicken Flocks: Association of Viral Pathotype with Lymphoma Frequency. Avian Dis. 1983, 27, 113–132. [Google Scholar] [CrossRef]
- Witter, R.L. Increased Virulence of Marek’s Disease Virus Field Isolates. Avian Dis. 1997, 41, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Bacon, L.D.; Hunt, H.D.; Cheng, H.H. A Review of the Development of Chicken Lines to Resolve Genes Determining Resistance to Diseases. Poult. Sci. 2000, 79, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Steep, A.C. A Genomic Investigation of Marek’s Disease Lymphomas; Michigan State University: East Lansing, MI, USA, 2019. [Google Scholar]
- Witter, R.L.; Sharma, J.M.; Fadly, A.M. Pathogenicity of Variant Marek’s Disease Virus Isolants in Vaccinated and Unvaccinated Chickens. Avian Dis. 1980, 24, 210–232. [Google Scholar] [CrossRef]
- Steep, A.C.; (Michigan State University, East Lansing, MI, USA). Personal Communication, 2019.
- Delany, M.E.; Gessaro, T.M.; Rodrigue, K.L.; Daniels, L.M. Chromosomal Mapping of Chicken Mega-Telomere Arrays to GGA9, 16, 28 and W Using a Cytogenomic Approach. Cytogenet. Genome Res. 2007, 117, 54–63. [Google Scholar] [CrossRef]
- McPherson, M.C.; Cheng, H.H.; Smith, J.M.; Delany, M.E. Vaccination and Host Marek’s Disease-Resistance Genotype Significantly Reduce Oncogenic Gallid Alphaherpesvirus 2 Telomere Integration in Host Birds. Cytogenet. Genome Res. 2018, 156, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Niikura, M.; Dodgson, J.; Cheng, H. Direct Evidence of Host Genome Acquisition by the Alphaherpesvirus Marek’s Disease Virus. Arch. Virol. 2006, 151, 537–549. [Google Scholar] [CrossRef]
- Masabanda, J.S.; Burt, D.W.; O’Brien, P.C.M.; Vignal, A.; Fillon, V.; Walsh, P.S.; Cox, H.; Tempest, H.G.; Smith, J.; Habermann, F.; et al. Molecular Cytogenetic Definition of the Chicken Genome: The First Complete Avian Karyotype. Genetics 2004, 166, 1367–1373. [Google Scholar] [CrossRef] [Green Version]
- Bloom, S.E.; Delany, M.E.; Muscarella, D.E. Constant and Variable Features of Avian Chromosomes. In Manipulation of the Avian Genome; CRC Press: Boca Raton, FL, USA, 1993; pp. 39–60. ISBN 978-0-203-74828-2. [Google Scholar]
- Schmid, M.; Nanda, I.; Guttenbach, M.; Steinlein, C.; Hoehn, M.; Schartl, M.; Haaf, T.; Weigend, S.; Fries, R.; Buerstedde, J.-M.; et al. First Report on Chicken Genes and Chromosomes 2000. Cytogenet. Genome Res. 2000, 90, 169–218. [Google Scholar] [CrossRef]
- Auer, H.; Mayr, B.; Lambrou, M.; Schleger, W. An Extended Chicken Karyotype, Including the NOR Chromosome. Cytogenet. Genome Res. 1987, 45, 218–221. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975; ISBN 978-0-471-68925-6. [Google Scholar]
- O’Hare, T.H.; Delany, M.E. Genetic Variation Exists for Telomeric Array Organization within and among the Genomes of Normal, Immortalized, and Transformed Chicken Systems. Chromosome Res. 2009, 17, 947. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glass, M.C.; Smith, J.M.; Cheng, H.H.; Delany, M.E. Marek’s Disease Virus Telomeric Integration Profiles of Neoplastic Host Tissues Reveal Unbiased Chromosomal Selection and Loss of Cellular Diversity during Tumorigenesis. Genes 2021, 12, 1630. https://doi.org/10.3390/genes12101630
Glass MC, Smith JM, Cheng HH, Delany ME. Marek’s Disease Virus Telomeric Integration Profiles of Neoplastic Host Tissues Reveal Unbiased Chromosomal Selection and Loss of Cellular Diversity during Tumorigenesis. Genes. 2021; 12(10):1630. https://doi.org/10.3390/genes12101630
Chicago/Turabian StyleGlass, Marla C., Justin M. Smith, Hans H. Cheng, and Mary E. Delany. 2021. "Marek’s Disease Virus Telomeric Integration Profiles of Neoplastic Host Tissues Reveal Unbiased Chromosomal Selection and Loss of Cellular Diversity during Tumorigenesis" Genes 12, no. 10: 1630. https://doi.org/10.3390/genes12101630