Gene Polymorphisms of TLR4 and TLR9 and Haemophilus influenzae Meningitis in Angolan Children

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Laboratory Tests, Clinical Findings, and Severity Factors of BM
2.3. DNA Isolation
2.4. Genotyping
2.5. Statistical Analyses
3. Results
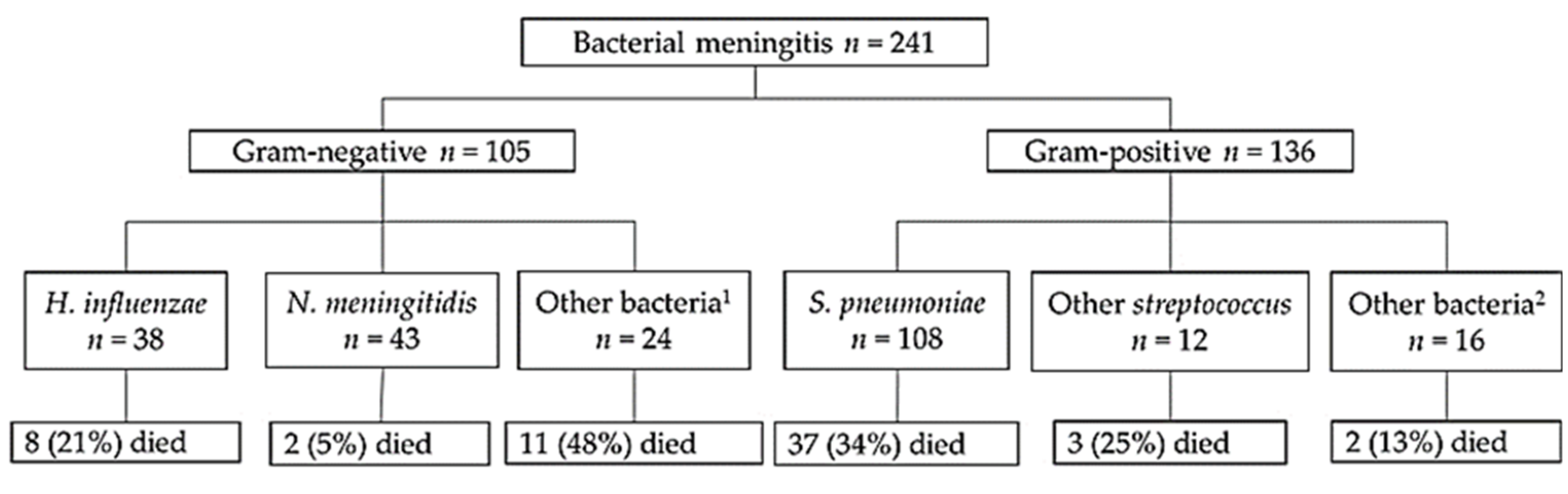
3.1. Detection of Pathogens and Laboratory Values
3.2. Association between Susceptability to BM and the Gene Polymorphisms of TLR4 and TLR9
3.3. Association between Laboratory Values, Severity Factors of BM, and the TLR4 and TLR9 Polymorphisms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paireau, J.; Chen, A.; Broutin, H.; Grenfell, B.; Basta, N.E. Seasonal dynamics of bacterial meningitis: A time-series analysis. Lancet Glob. Health 2016, 4, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.S.; van Well, G.T.J.; Ouburg, S.; Morré, S.A.; van Furth, A.M. Genetic variation of innate immune response genes in invasive pneumococcal and meningococcal disease applied to the pathogenesis of meningitis. Genes Immun. 2011, 12, 321–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Zhang, N.; Liu, L.; Zheng, K.; Zhu, L.; Zhu, J.; Cao, L.; Jiang, Y.; Liu, G.; He, Q. Polymorphisms of toll-like receptors 2 and 9 and severity and prognosis of bacterial meningitis in Chinese children. Sci. Rep. 2017, 7, 42796. [Google Scholar] [CrossRef] [PubMed]
- Weller, R.O.; Sharp, M.M.; Christodoulides, M.; Carare, R.O.; Møllgård, K. The meninges as barriers and facilitators for the movement of fluid, cells and pathogens related to the rodent and human CNS. Acta Neuropathol 2018, 135, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Pelkonen, T.; Roine, I.; Monteiro, L.; Correia, M.; Pitkäranta, A.; Bernardino, L.; Peltola, H. Risk Factors for Death and Severe Neurological Sequelae in Childhood Bacterial Meningitis in Sub-Saharan Africa. Clin. Infect. Dis 2009, 48, 1107–1110. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.M.C.; Maiden, M.C.J. A world without bacterial meningitis: How genomic epidemiology can inform vaccination strategy. F1000Res 2018, 7. [Google Scholar] [CrossRef]
- Saikia, K.K.; Das, B.K.; Bewal, R.K.; Kapil, A.; Arora, N.K.; Sood, S. Characterization of nasopharyngeal isolates of type b Haemophilus influenzae from Delhi. Indian J. Med. Res. 2012, 136, 855–861. [Google Scholar]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol 2018, 16, 355–367. [Google Scholar] [CrossRef]
- Heckenberg, S.G.B.; Brouwer, M.C.; van de Beek, D. Chapter 93. Bacterial meningitis. In Handbook of Clinical Neurology; Neurologic Aspects of Systemic Disease Part III; Biller, J., Ferro, J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 121, pp. 1361–1375. [Google Scholar]
- Gowin, E.; Januszkiewicz-Lewandowska, D. Genes and their single nucleotide polymorphism involved in innate immune response in central nervous system in bacterial meningitis: Review of literature data. Inflamm. Res. 2018, 67, 655–661. [Google Scholar] [CrossRef]
- Urtti, S.; Cruzeiro, M.L.; de Gouveia, L.; Peltola, H.; von Gottberg, A.; Kyaw, M.H.; Pelkonen, T. Surveillance of bacterial meningitis in an Angolan pediatric hospital after the introduction of pneumococcal conjugate vaccines. J. Glob. Health Rep. 2019, 3, e2019091. [Google Scholar] [CrossRef]
- WHO | Number of Suspected Meningitis Cases and Deaths Reported. Available online: https://www.who.int/gho/epidemic_diseases/meningitis/suspected_cases_deaths_text/en/ (accessed on 29 January 2019).
- Angola. Available online: https://www.gavi.org/programmes-impact/country-hub/africa/angola (accessed on 14 July 2020).
- Peltola, H.; Pelkonen, T.; Bernardino, L.; Monteiro, L.; Silvestre, S. da C.; Anjos, E.; Cruzeiro, M.L.; Pitkäranta, A.; Roine, I. Vaccine-induced waning of Haemophilus influenzae Empyema and Meningitis, Angola. Emerg. Infect. Dis 2014, 20, 1887–1890. [Google Scholar] [CrossRef] [PubMed]
- Malley, R.; Henneke, P.; Morse, S.C.; Cieslewicz, M.J.; Lipsitch, M.; Thompson, C.M.; Kurt-Jones, E.; Paton, J.C.; Wessels, M.R.; Golenbock, D.T. Recognition of pneumolysin by Toll-like receptor 4 confers resistance to pneumococcal infection. Proc. Natl Acad Sci USA 2003, 100, 1966–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanke, M.L.; Kielian, T. Toll-like receptors in health and disease in the brain: Mechanisms and therapeutic potential. Clin. Sci. 2011, 121, 367–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Csakai, A.; Jin, J.; Zhang, F.; Yin, H. Therapeutic developments targeting oll-like receptor 4 mediated neuroinflammation. ChemMedChem 2016, 11, 154–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moresco, E.M.Y.; LaVine, D.; Beutler, B. Toll-like receptors. Curr. Biol. 2011, 21, R488–R493. [Google Scholar] [CrossRef] [Green Version]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F. TLR4 Signaling pathway modulators as potential therapeutics in inflammation and sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ohto, U.; Shimizu, T. Toward a structural understanding of nucleic acid-sensing Toll-like receptors in the innate immune system. FEBS Lett. 2017, 591, 3167–3181. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H.; Paludan, S.R.; Kilian, M.; Østergaard, L. Live Streptococcus pneumoniae, Haemophilus influenzae, and Neisseria meningitidis activate the inflammatory response through Toll-like receptors 2, 4, and 9 in species-specific patterns. J. Leukocyte Biol. 2006, 80, 267–277. [Google Scholar] [CrossRef]
- Skevaki, C.; Pararas, M.; Kostelidou, K.; Tsakris, A.; Routsias, J.G. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious diseases: TLR SNPs and infectious diseases. Clin. Exp. Immunol. 2015, 180, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Long, H.; O’Connor, B.P.; Zemans, R.L.; Zhou, X.; Yang, I.V.; Schwartz, D.A. The Toll-Like Receptor 4 Polymorphism Asp299Gly but Not Thr399Ile Influences TLR4 Signaling and Function. PLoS ONE 2014, 9, e093550. [Google Scholar] [CrossRef] [Green Version]
- Teräsjärvi, J.T.; Toivonen, L.; Vuononvirta, J.; Mertsola, J.; Peltola, V.; He, Q. TLR4 Polymorphism, Nasopharyngeal Bacterial Colonization, and the development of childhood asthma: A prospective birth-cohort study in Finnish children. Genes 2020, 11, 768. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, E.; Mira, J.P.; Frees, K.L.; Schwartz, D.A. Relevance of mutations in the TLR4 receptor in patients with gram-negative septic shock. Arch. Intern. Med. 2002, 162, 1028–1032. [Google Scholar] [CrossRef]
- Tulic, M.K.; Hurrelbrink, R.J.; Prêle, C.M.; Laing, I.A.; Upham, J.W.; Souef, P.L.; Sly, P.D.; Holt, P.G. TLR4 Polymorphisms mediate impaired responses to respiratory syncytial virus and lipopolysaccharide. J. Immunol. 2007, 179, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, R.C.; Aragaki, C.C.; Rivera-Chavez, F.A.; Purdue, G.F.; Hunt, J.L.; Horton, J.W. TLR4 and TNF-α polymorphisms are associated with an increased risk for severe sepsis following burn injury. J. Med. Genet. 2004, 41, 808–813. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, M.C.; de Gans, J.; Heckenberg, S.G.; Zwinderman, A.H.; van der Poll, T.; van de Beek, D. Host genetic susceptibility to pneumococcal and meningococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2009, 9, 31–44. [Google Scholar] [CrossRef]
- Allen, A.; Obaro, S.; Bojang, K.; Awomoyi, A.A.; Greenwood, B.M.; Whittle, H.; Sirugo, G.; Newport, M.J. Variation in Toll-like receptor 4 and susceptibility to group a meningococcal meningitis in Gambian children. Pediatric Infect. Dis. J. 2003, 22, 1018. [Google Scholar] [CrossRef] [PubMed]
- Gowin, E.; Świątek-Kościelna, B.; Kałużna, E.; Nowak, J.; Michalak, M.; Wysocki, J.; Januszkiewicz-Lewandowska, D. Analysis of TLR2, TLR4, and TLR9 single nucleotide polymorphisms in children with bacterial meningitis and their healthy family members. Int. J. Infect. Dis. 2017, 60, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Zhang, L.; Yang, T.; Wei, X.; Zhang, L.; Yu, Y.; Li, Y.; Cao, D.; Yang, X. The Associations between Toll-Like Receptor 9 Gene Polymorphisms and Cervical Cancer Susceptibility. Mediat. Inflamm. 2018, 2018, 9127146. [Google Scholar] [CrossRef]
- Gębura, K.; Świerkot, J.; Wysoczańska, B.; Korman, L.; Nowak, B.; Wiland, P.; Bogunia-Kubik, K. Polymorphisms within Genes Involved in Regulation of the NF-κB Pathway in Patients with Rheumatoid Arthritis. Int. J. Mol. Sci. 2017, 18, 1432. [Google Scholar] [CrossRef]
- Bharti, D.; Kumar, A.; Mahla, R.S.; Kumar, S.; Ingle, H.; Shankar, H.; Joshi, B.; Raut, A.A.; Kumar, H. The role of TLR9 polymorphism in susceptibility to pulmonary tuberculosis. Immunogenetics 2014, 66, 675–681. [Google Scholar] [CrossRef]
- van Well, G.T.J.; Sanders, M.S.; Ouburg, S.; van Furth, A.M.; Morré, S.A. Polymorphisms in Toll-Like Receptors 2, 4, and 9 Are Highly Associated with Hearing Loss in Survivors of Bacterial Meningitis. PLoS ONE. 2012, 7, e35837. [Google Scholar] [CrossRef]
- Sanders, M.S.; van Well, G.T.J.; Ouburg, S.; Lundberg, P.S.J.; van Furth, A.M.; Morré, S.A. Single Nucleotide Polymorphisms in TLR9 Are Highly Associated with Susceptibility to Bacterial Meningitis in Children. Clin. Infect. Dis 2011, 52, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, T.; Roine, I.; Cruzeiro, M.L.; Pitkäranta, A.; Kataja, M.; Peltola, H. Slow initial β-lactam infusion and oral paracetamol to treat childhood bacterial meningitis: A randomised, controlled trial. Lancet Infect. Dis. 2011, 11, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Savonius, O.; Rugemalira, E.; Roine, I.; Cruzeiro, M.L.; Peltola, H.; Pelkonen, T. Extended Continuous β-Lactam Infusion with Oral Acetaminophen in Childhood Bacterial Meningitis: A Randomized, Double-Blind Clinical Trial. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Pelkonen, T.; Roine, I.; Monteiro, L.; Simões, M.J.; Anjos, E.; Pelerito, A.; Pitkäranta, A.; Bernardino, L.; Peltola, H. Acute childhood bacterial meningitis in Luanda, Angola. Scand. J. Infect. Dis. 2008, 40, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Teräsjärvi, J.; Hakanen, A.; Korppi, M.; Nuolivirta, K.; Gröndahl-Yli-Hannuksela, K.; Mertsola, J.; Peltola, V.; He, Q. Rapid detection of functional gene polymorphisms of TLRs and IL-17 using high resolution melting analysis. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Korppi, M.; Teräsjärvi, J.; Lauhkonen, E.; Huhtala, H.; Nuolivirta, K.; He, Q. Toll-like receptor 4 polymorphisms were associated with low serum pro-inflammatory cytokines in BCG osteitis survivors. Acta Paediatr. 2019. [Google Scholar] [CrossRef]
- Yuan, F.F.; Marks, K.; Wong, M.; Watson, S.; de Leon, E.; McIntyre, P.B.; Sullivan, J.S. Clinical relevance of TLR2, TLR4, CD14 and FcγRIIA gene polymorphisms in Streptococcus pneumoniae infection. Immunol. Cell Biol. 2008, 86, 268–270. [Google Scholar] [CrossRef]
- Smirnova, I.; Mann, N.; Dols, A.; Derkx, H.H.; Hibberd, M.L.; Levin, M.; Beutler, B. Assay of locus-specific genetic load implicates rare Toll-like receptor 4 mutations in meningococcal susceptibility. PNAS 2003, 100, 6075–6080. [Google Scholar] [CrossRef] [Green Version]
- Moens, L.; Verhaegen, J.; Pierik, M.; Vermeire, S.; De Boeck, K.; Peetermans, W.E.; Bossuyt, X. Toll-like receptor 2 and Toll-like receptor 4 polymorphisms in invasive pneumococcal disease. Microbes Infect. 2007, 9, 15–20. [Google Scholar] [CrossRef]
- Read, R.C.; Pullin, J.; Gregory, S.; Borrow, R.; Kaczmarski, E.B.; di Giovine, F.S.; Dower, S.K.; Cannings, C.; Wilson, A.G. A functional polymorphism of toll-like receptor 4 Is not associated with likelihood or severity of eningococcal Disease. J. Infect. Dis 2001, 184, 640–642. [Google Scholar] [CrossRef] [Green Version]
- Biebl, A.; Muendlein, A.; Kazakbaeva, Z.; Heuberger, S.; Sonderegger, G.; Drexel, H.; Lau, S.; Nickel, R.; Kabesch, M.; Simma, B. CD14 C-159T and Toll-Like Receptor 4 Asp299Gly Polymorphisms in surviving Meningococcal Disease patients. PLoS ONE 2009, 4, e7374. [Google Scholar] [CrossRef]
- Davis, S.M.; Clark, E.A.S.; Nelson, L.T.; Silver, R.M. The association of innate immune response gene polymorphisms and puerperal group A streptococcal sepsis. Am. J. Obstet. Gynecol. 2010, 202, 308.e1–308.e8. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-H.; Zeng, L.; Gu, W.; Zhou, J.; Du, D.-Y.; Jiang, J.-X. Polymorphisms in the toll-like receptor 9 gene associated with sepsis and multiple organ dysfunction after major blunt trauma. BJS (Br. J. Surg.) 2011, 98, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Nurjadi, D.; Heeg, K.; Weber, A.N.R.; Zanger, P. Toll-like receptor 9 (TLR-9) promotor polymorphisms and gene expression are associated with persistent Staphylococcus aureus nasal carriage. Clin. Microbiol. Infect. 2018, 24, 1210.e7–1210.e12. [Google Scholar] [CrossRef] [Green Version]
- rs4986790 (SNP). Population Genetics. Homo Sapiens. Ensembl Genome Browser 98. Available online: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=9:117712524-117713524;v=rs4986790;vdb=variation;vf=152699755 (accessed on 17 December 2019).
- rs187084 (SNP). Population Genetics. Homo Sapiens. Ensembl Genome Browser 98. Available online: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=3:52226515-52227515;v=rs187084;vdb=variation;vf=37101106 (accessed on 17 December 2019).
- Nigrovic, L.E.; Kimia, A.A.; Shah, S.S.; Neuman, M.I. Relationship between Cerebrospinal Fluid Glucose and Serum Glucose. N. Eng. J. Med. 2012, 366, 576–578. [Google Scholar] [CrossRef] [Green Version]
- Savonius, O.; Roine, I.; Alassiri, S.; Tervahartiala, T.; Helve, O.; Fernández, J.; Peltola, H.; Sorsa, T.; Pelkonen, T. The Potential Role of Matrix Metalloproteinases 8 and 9 and Myeloperoxidase in Predicting Outcomes of Bacterial MENINGITIS of Childhood. Available online: https://www.hindawi.com/journals/mi/2019/7436932/ (accessed on 25 May 2020).
- Leppert, D.; Leib, S.L.; Grygar, C.; Miller, K.M.; Schaad, U.B.; Holländer, G.A. Matrix Metalloproteinase (MMP)-8 and MMP-9 in Cerebrospinal fluid during Bacterial Meningitis: Association with blood-brain barrier damage and Neurological Sequelae. Clin. Infect. Dis. 2000, 31, 80–84. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Target | SNP ID | Location | Allele Frequency in the African Population | Allele Frequency in this Study | Potential Effect and Reported Associations |
|---|---|---|---|---|---|
| TLR4 | rs4986790 | Chromosome 9 In gene 896A > G | A: 0.93 G: 0.07 | A: 0.91 G: 0.09 | Diminished responsiveness: M. catarrhalis and H. influenzae, pneumococcal and meningococcal infections, septic shock, and respiratory syncytial virus (RSV) |
| TLR9 | rs187084 | Chromosome 3 Promoter area −1486T > C | T: 0.71 C: 0.29 | T: 0.73 C: 0.27 | Associated with autoimmune disorders and cancers, RSV, Acinetobacter baumannii, tuberculosis, and streptococcal infections |
| Characteristics | Total Subjects 1 | N (%) | Value Distribution | Range |
|---|---|---|---|---|
| Age in months, median (IQR) | 241 | - | 15 (6, 43) | 1–161 |
| Male | 241 | 127 (53%) | - | - |
| Weight (kg), median (IQR) | 241 | - | 9 (6.8, 12.7) | 3.6–36 |
| Clinical features | ||||
| Fever (°C), mean (SD) | 239 | 50 2 | 37.5 (0.99) | 35–40 |
| Length of hospital stay in days, median (IQR) 3 | 158 | - | 12 (10, 36.05) | 6–63 |
| Poor general condition | 238 | 142 (60%) | - | - |
| Convulsion in hospital | 235 | 155 (66%) | - | - |
| Altered consciousness during admission | 237 | 166 (70%) | - | - |
| Additional focus of infection present | 232 | 69 (30%) | - | - |
| Dyspnea | 238 | 125 (53%) | - | - |
| Glasgow coma score, median (IQR) | 213 | - | 12 (9, 15) | 3–15 |
| Blantyre coma score, median (IQR) | 212 | - | 4 (3, 5) | 1–5 |
| Fatal outcome | 241 | 63 (26%) | - | - |
| Severe neurological sequelae at discharge | 177 | 24 (14%) | - | - |
| Laboratory tests, median (IQR) | ||||
| CRP (mg/L) | 207 | - | 161 (128, 161) | 7–340 |
| Glucose in CSF (mg/dL) 4 | 233 | - | 10 (5.5, 19.55) | 0.5–270 |
| Protein in CSF (mg/dL) 4 | 118 | - | 201.15 (136.2, 269) | 10.2–1806 |
| Leukocytes in CSF (/mm3) 4 | 241 | - | 1560 (414.5, 3590) | 8–32,400 |
| Blood leukocytes (/µL) | 182 | - | 15.3 (10.2, 21.4) | 1.23–57 |
| MMP-8 in CSF (ng/mL) | 92 | - | 854.2 (250, 1220) | 3.74–3560 |
| Gram-Negative Bacteria | Gram-Positive Bacteria | |||||
|---|---|---|---|---|---|---|
| H. Influenzae | N. Meningitidis | Other Bacteria | S. Pneumoniae | Other Bacteria | Control Group | |
| TLR4 rs4986790 | ||||||
| AA (%) | 26 (68.4) | 40 (93.0) | 22 (91.6) | 92 (85.2) | 23 (82.1) | 224 (84.5) |
| AG (%) | 10 (26.3) | 2 (4.6) | 2 (8.3) | 15 (13.9) | 5 (17.9) | 37 (13.9) |
| GG (%) | 2 (5.3) | 1 (2.3) | 0 | 1 (0.9) | 0 | 4 (1.5) |
| Total n | 38 | 43 | 24 | 108 | 28 | 265 |
| HWE | 0.44 | 0.002 | 0.831 | 0.661 | 0.604 | 0.099 |
| p-Value 1 | 0.022 | 0.164 | 0.549 | 0.999 | 0.784 | Ref. |
| OR (95% CI) | 2.5 (1.2–5.4) | 0.4 (0.1–1.4) | 0.5 (0.11–2.2) | 1 (0.5–1.7) | 1.2 (0.43–3.3) | |
| TLR9 rs187084 | ||||||
| TT (%) | 27 (71.1) | 21 (50.0) | 14 (58.3) | 57 (55.3) | 13 (46.4) | 136 (51.7) |
| CT (%) | 8 (21.1) | 20 (47.6) | 9 (37.5) | 45 (43.7) | 13 (46.4) | 106 (40.3) |
| CC (%) | 3 (7.9) | 1 (2.4) | 1 (4.2) | 5 (4.9) | 2 (7.1) | 21 (8.0) |
| Total n | 38 | 42 | 24 | 103 | 2 | 263 |
| HWE | 0.065 | 0.133 | 0.764 | 0.296 | 0.572 | 0.957 |
| p-Value | 0.036 | 0.869 | 0.670 | 0.819 | 0.692 | Ref. |
| OR (95% CI) | 0.4 (0.2–0.9) | 1.1 (0.6–2.1) | 0.76 (0.33–1.8) | 0.9 (0.6–1.5) | 1.2 (0.57–2.7) | |
| Clinical Features | TLR 4 | OR (95% Cl) | p-Value | |
|---|---|---|---|---|
| AA | AG and GG | |||
| Poor general condition 1 | 40 (46.0) | 11 (64.7) | 2.15 (0.73–6.35) | 0.191 |
| Convulsions during admission 1 | 32 (37.6) | 8 (50.0) | 1.66 (0.57–4.85) | 0.409 |
| Level of consciousness 1 | ||||
| Normal | 37 (43.5) | 5 (29.4) | - | 0.544 |
| Altered | 43 (50.6) | 11 (64.7) | - | |
| Coma | 5 (5.9) | 1 (5.9) | - | |
| Glasgow coma score (<12) 1 | 31 (38.3) | 9 (56.3) | 2.07 (0.70–6.14) | 0.266 |
| Other focus of infection during hospital stay | 49 (56.3) | 10 (58.8) | 1.11 (0.39–3.18) | 0.999 |
| Pneumonia during hospital stay | 31 (35.2) | 8 (47.1) | 1.63 (0.57–4.66) | 0.415 |
| Outcome | ||||
| Fatal | 15 (17.0) | 6 (35.3) | 2.66 (0.85–8.30) | 0.102 |
| Severe neurological sequelae 3 | 5 (6.9) | 2 (18.2) | 2.98 (0.50–17.68) | 0.231 |
| Deafness 3 | 5 (8.6) | 0 (0.0) | 0.91 (0.84–0.99) | 0.999 |
| Blindness 3 | 1 (1.4) | 2 (20.0) | 17.5 (1.42–215.21) - 2 | 0.039 (0.99) 2 |
| Any neurological sequelae (no ataxia) 3 | 6 (8.3) | 4 (40.0) | 7.33 (1.31–33.41) 11.85 (1.07–131.49) 2 | 0.017 (0.044) 2 |
| Ataxia 3 | 12 (16.9) | 7 (70.0) | 11.47 (2.59–50.80) 12.91 (1.52–109.80) 2 | 0.001 (0.019) 2 |
| Laboratory variables, median (IQR) | ||||
| CRP (mg/L) | 161.00 (131.5, 172.0) (n = 77) | 160.00 (70.5, 161.0) (n = 13) | - | 0.183 |
| CSF glucose 1 (mg/dL) | 10.40 (4.80, 22.10) (n = 88) | 9.30 (5.15, 26.63) (n = 16) | - | 0.786 |
| CSF protein 1 (mg/dL) | 191.85 (124.9, 257.15) (n = 46) | 187.05 (146.48, 282.55) (n = 6) | - | 0.748 |
| CSF leukocytes 1 (/mm3) | 1612.50 (970.00, 3830.50) (n = 88) | 1440.00 (857.50, 1440.00) (n = 17) | - | 0.748 |
| Blood leukocytes (/µL) | 16.11 (11.04, 24.35) (n = 73) | 13.96 (8.23, 22.20) (n = 12) | - | 0.434 |
| CSF MMP-8 1 (ng/mL) | 1132.57 (834.04, 1641.75) (n = 30) | 225.93 (63.56, 556.31) (n = 6) | - | 0.004 |
| Clinical Features | TLR9 | OR (95% Cl) | p-Value | |
|---|---|---|---|---|
| TT | CT and CC | |||
| Poor general condition 1 | 28 (45.9) | 23 (54.8) | 1.43 (0.65–3.14) | 0.426 |
| Convulsions during admission 1 | 28 (46.7) | 12 (30,0) | 0.490 (0.21–1.14) | 0.144 |
| Level of consciousness 1 | ||||
| Normal | 25 (41.7) | 17 (41.5) | - | 0.928 |
| Altered | 31 (51.7) | 22 (53.7) | - | |
| Coma | 4 (6.7) | 2 (4.9) | - | |
| Glasgow coma score (<12) 1 | 23 (41.1) | 17 (42.5) | 1.06 (0.47–2.41) | 0.999 |
| Other focus of infection during hospital stay | 35 (57.4) | 23 (54.8) | 0.90 (0.41–1.99) | 0.841 |
| Pneumonia during hospital stay | 24 (38.7) | 15 (35.7) | 0.88 (0.39–1.98) | 0.838 |
| Outcome | ||||
| Fatal | 12 (19.4) | 9 (21.4) | 1.14 (0.43–3.00) | 0.808 |
| Severe neurological sequelae 2 | 7 (14.0) | 0 (0.0) | 0.86 (0.77–0.96) 0.00 (0.00–0.00) 3 | 0.039 0.999 2 |
| Deafness 2 | 3 (7.7) | 2 (7.4) | 0.96 (0.15–6.17) | 0.999 |
| Blindness 2 | 3 (3.1) | 0 (0.0) | 0.94 (0.87–1.01) | 0.279 |
| Any neurological sequelae (no ataxia) 2 | 9 (18.0) | 1 (3.2) | 0.15 (0.02–1.26) 0.17 (0.02–1.47) 3 | 0.049 (0.107) |
| Ataxia 2 | 16 (32.7) | 3 (9.7) | 0.22 (0.06–0.84) 0.25 (0.06–1.04) 2 | 0.029 (0.057) 2 |
| Laboratory variables, median (IQR) | ||||
| CRP (mg/L) | 161.00 (108.5,173.3) (n = 50) | 161.00 (133.0, 161.0) (n = 39) | - | 0.814 |
| CSF glucose (mg/dL) 1 | 8.75 (4.73, 17.33) (n = 60) | 16.40 (7.88, 25.53) (n = 38) | - | 0.252 |
| CSF protein (mg/dL) 1 | 189.10 (125.50, 275.00) (n = 31) | 189.40 (131.15, 249.35) (n = 20) | - | 0.549 |
| CSF leukocytes (/mm3) 1 | 1518.50 (850.00, 3125.00) (n = 62) | 1725.00 (990.00, 5607.50) (n = 42) | - | 0.991 |
| Blood leukocytes (/µL) | 15.74 (10.89, 26.05) (n = 52) | 15.63 (10.71, 21.14) (n = 32) | - | 0.826 |
| CSF MMP-8 (ng/mL) median | 958.28 (319.46, 1496.89) (n = 20) | 1187.47 (866.80, 1632.45) (n = 16) | - | 0.278 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenhu, E.; Teräsjärvi, J.; Cruzeiro, M.L.; Savonius, O.; Rugemalira, E.; Roine, I.; He, Q.; Pelkonen, T. Gene Polymorphisms of TLR4 and TLR9 and Haemophilus influenzae Meningitis in Angolan Children. Genes 2020, 11, 1099. https://doi.org/10.3390/genes11091099
Tenhu E, Teräsjärvi J, Cruzeiro ML, Savonius O, Rugemalira E, Roine I, He Q, Pelkonen T. Gene Polymorphisms of TLR4 and TLR9 and Haemophilus influenzae Meningitis in Angolan Children. Genes. 2020; 11(9):1099. https://doi.org/10.3390/genes11091099
Chicago/Turabian StyleTenhu, Elina, Johanna Teräsjärvi, Manuel Leite Cruzeiro, Okko Savonius, Emilie Rugemalira, Irmeli Roine, Qiushui He, and Tuula Pelkonen. 2020. "Gene Polymorphisms of TLR4 and TLR9 and Haemophilus influenzae Meningitis in Angolan Children" Genes 11, no. 9: 1099. https://doi.org/10.3390/genes11091099