Export of Rgg Quorum Sensing Peptides is Mediated by the PptAB ABC Transporter in Streptococcus Thermophilus Strain LMD-9

 and
and
Abstract
:
Highlights
- PptAB exports signaling peptides from intracellular quorum sensing systems in Streptococcus thermophilus;
- PptAB only exports mature signaling peptides;
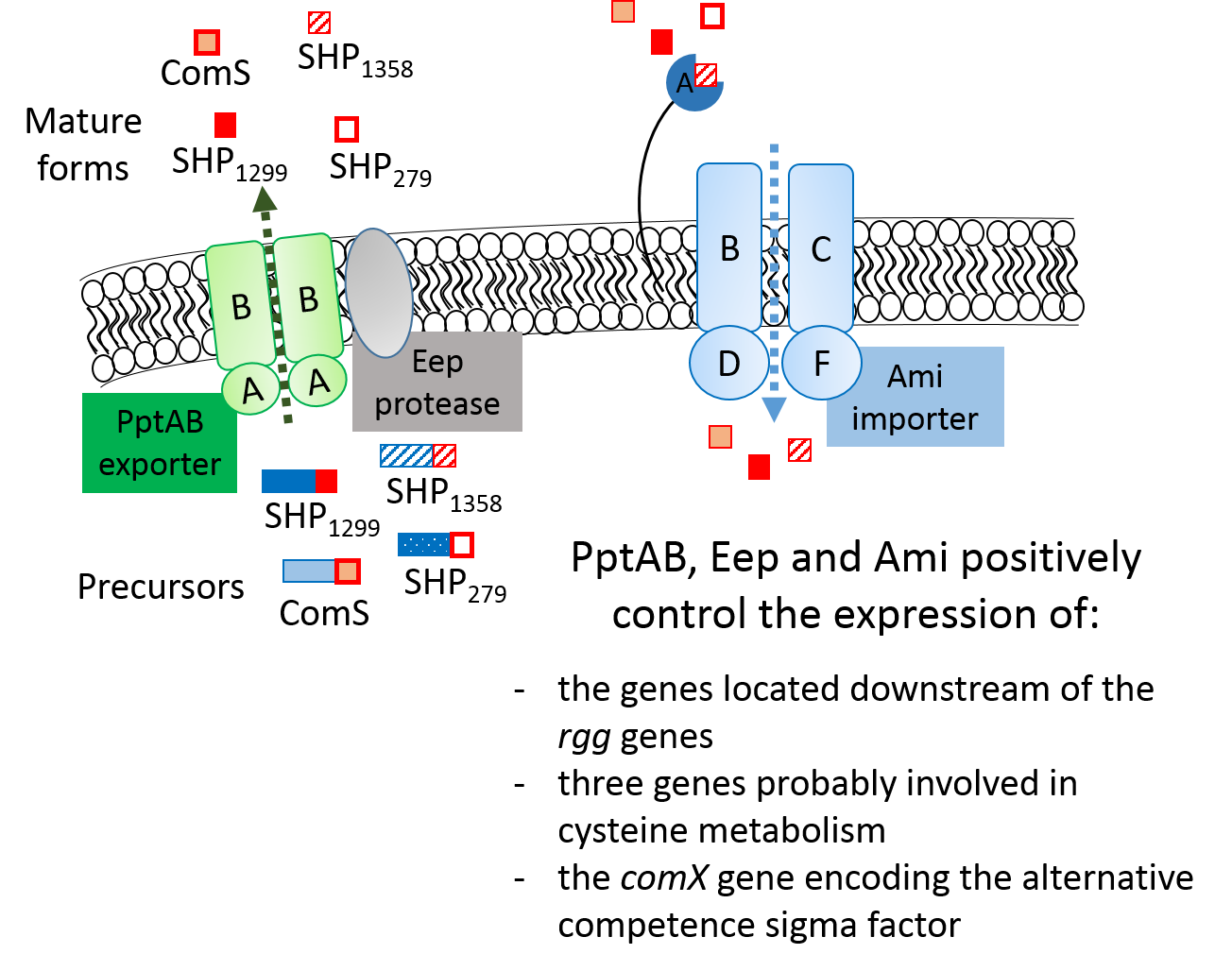
- in the S. thermophilus strain LMD-9, PptAB, Eep (a protease), and Ami (a peptide importer) are involved in a QS mechanism that controls the expression of genes located downstream all rgg genes but also of a few distal targets; and,
- at the strain level, crosstalk between the SHP/Rgg systems likely occurs and involves a few SHP signaling peptides that interact with multiple Rgg regulators.
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. DNA Manipulation and Sequencing
2.3. Construction of the Mutant Strains
2.4. Mass Spectrometry Analysis
2.5. Luciferase Assays
2.6. Total RNA Extraction
2.7. RNA Sequencing and Transcriptome Analysis
2.8. Bioinformatic Tools
3. Results
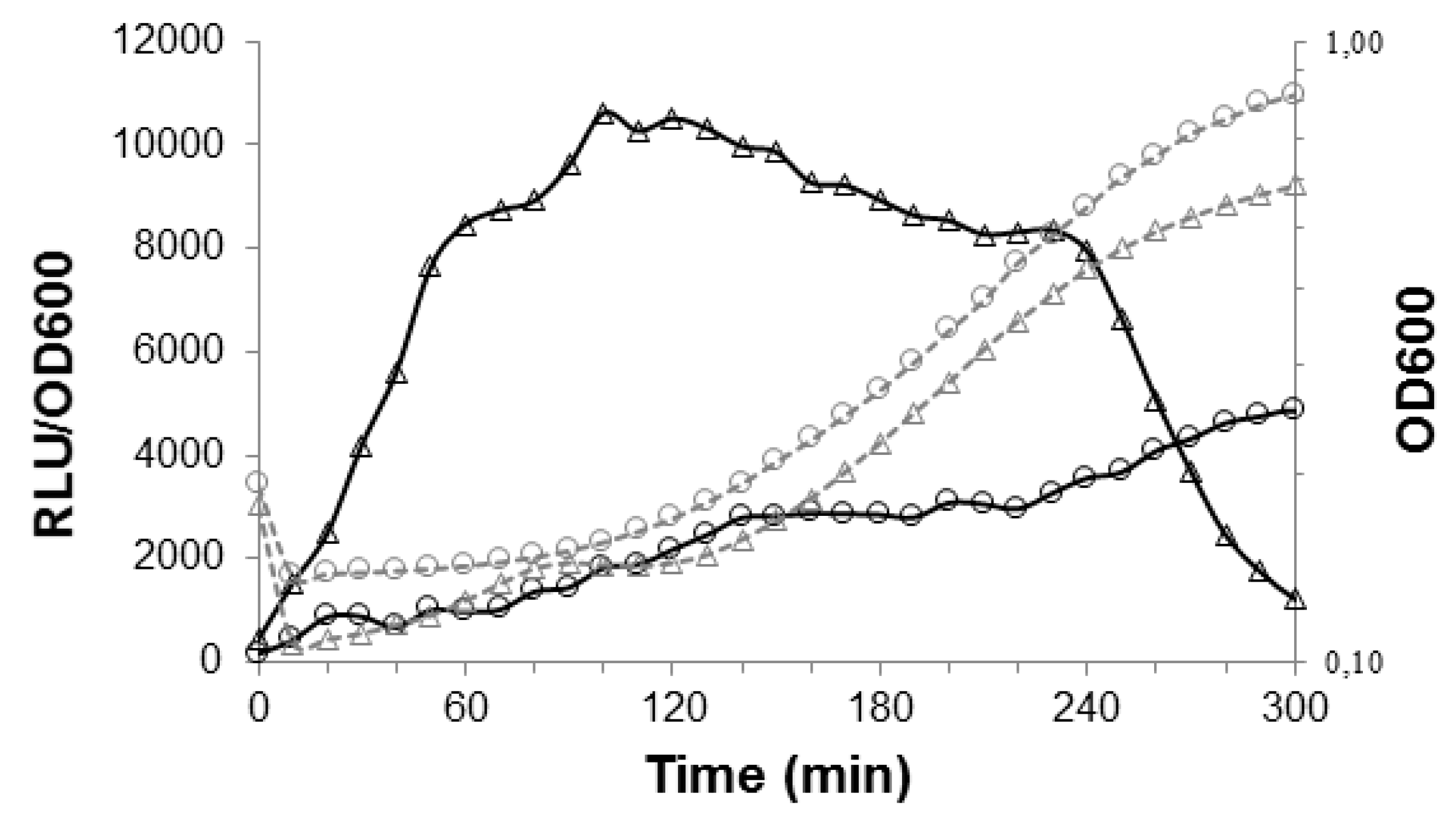
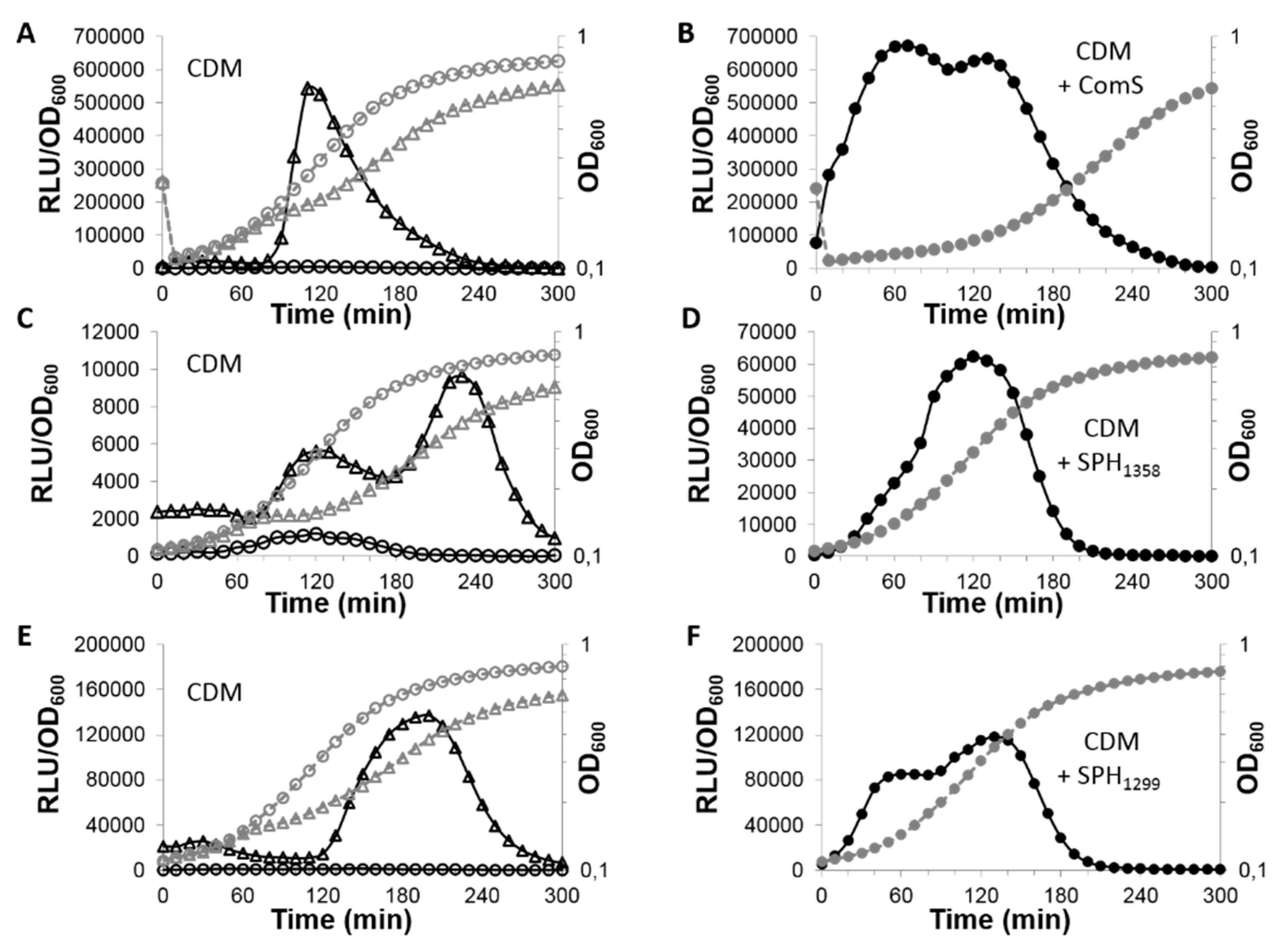
3.1. The PptAB Genes Are Expressed in S. thermophilus and Required for the Positive Control of Three Signaling Systems
3.2. PptAB is Involved in the Export of SHP1358 and SHP1299
3.3. PptAB Only Exports the Mature Form of SHP1358
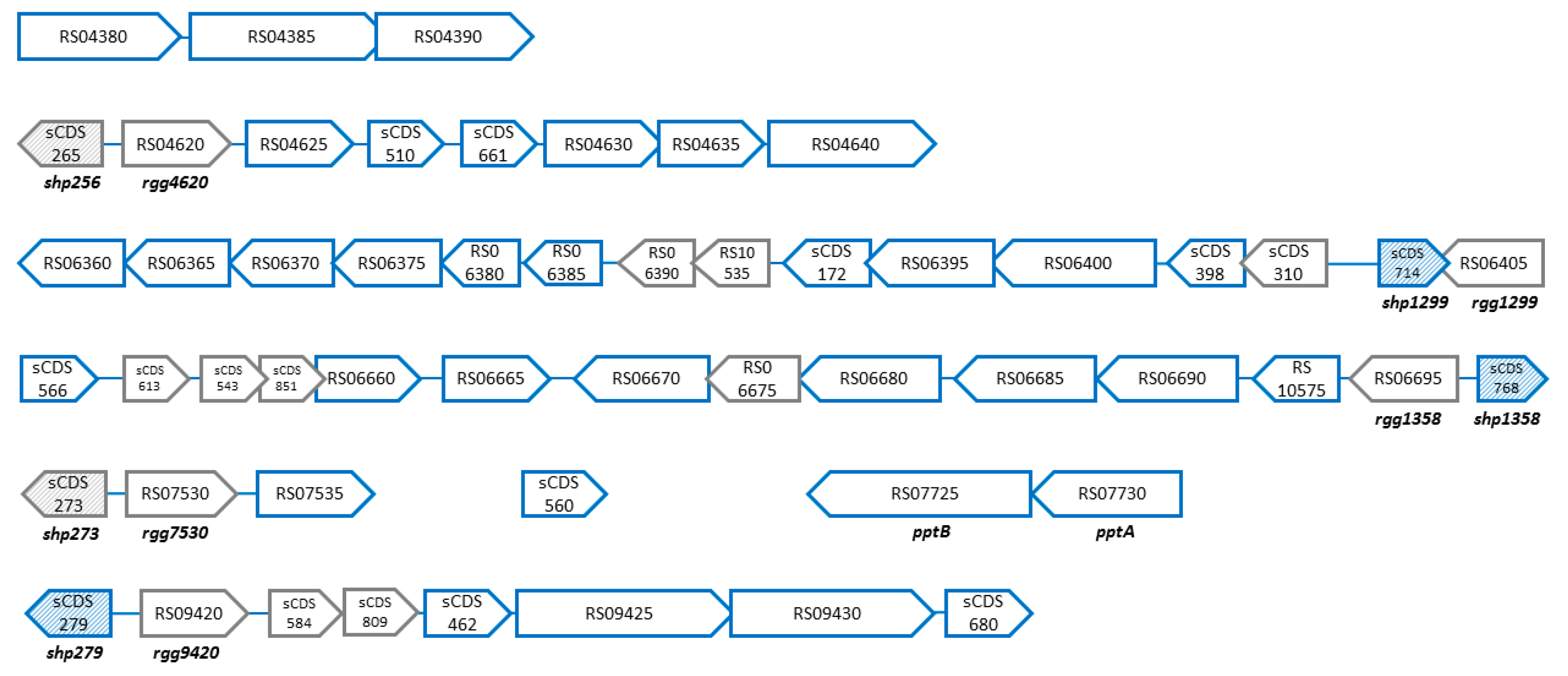
3.4. PptAB Controls Transcription of the Genes Located Downstream from the Rgg Genes in S. thermophilus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nealson, K.H. On the 50th Anniversary of the discovery of autoinduction and the ensuing birth of quorum sensing. Environ. Microbiol. 2020, 22, 801–807. [Google Scholar] [CrossRef]
- Verma, S.C.; Miyashiro, T. Quorum sensing in the squid-Vibrio symbiosis. Int. J. Mol. Sci. 2013, 14, 16386–16401. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, L.; Wahl, A.; Fléchard, M.; Mignolet, J.; Hols, P. Regulation of competence for natural transformation in streptococci. Infect. Genet. Evol. 2015, 33, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M. Quorum sensing control of lantibiotic production; nisin and subtilin autoregulate their own biosynthesis. Peptides 2004, 25, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Le, K.Y.; Otto, M. Quorum-sensing regulation in staphylococci-an overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnet, V.; Juillard, V.; Gardan, R. Peptide conversations in Gram-positive bacteria. Crit. Rev. Microbiol. 2014, 42, 339–351. [Google Scholar] [CrossRef]
- Kalamara, M.; Spacapan, M.; Mandic-Mulec, I.; Stanley-Wall, N.R. Social behaviours by Bacillus subtilis: Quorum sensing, kin discrimination and beyond. Mol. Microbiol. 2018, 110, 863–878. [Google Scholar] [CrossRef] [Green Version]
- Shanker, E.; Federle, M.J. Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans. Genes 2017, 8, 15. [Google Scholar] [CrossRef]
- Rocha-Estrada, J.; Aceves-Diez, A.E.; Guarneros, G.; de la Torre, M. The RNPP family of quorum-sensing proteins in Gram-positive bacteria. Appl. Microbiol. Biotechnol. 2010, 87, 913–923. [Google Scholar] [CrossRef]
- Neiditch, M.B.; Capodagli, G.C.; Prehna, G.; Federle, M.J. Genetic and structural analyses of RRNPP intercellular peptide signaling of Gram-positive bacteria. Annu. Rev. Genet. 2017, 51, 311–333. [Google Scholar] [CrossRef]
- Pottathil, M.; Lazazzera, B.A. The extracellular Phr Peptide-Rap phosphatase signaling circuit of Bacillus subtilis. Front. Biosci. 2003, 8, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Slamti, L.; Perchat, S.; Huillet, E.; Lereclus, D. Quorum sensing in Bacillus thuringiensis is required for completion of a full infectious cycle in the insect. Toxins 2014, 6, 2239–2255. [Google Scholar] [CrossRef] [PubMed]
- Dunny, G.M.; Berntsson, R.P. Enterococcal sex pheromones: Evolutionary pathways to complex, two-signal systems. J. Bacteriol. 2016, 198, 1556–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Declerck, N.; Bouillaut, L.; Chaix, D.; Rugani, N.; Slamti, L.; Hoh, F.; Lereclus, D.; Arold, S.T. Structure of PlcR: Insights into virulence regulation and evolution of quorum sensing in Gram-positive bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 18490–18495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parashar, V.; Aggarwal, C.; Federle, M.J.; Neiditch, M.B. Rgg protein structure-function and inhibition by cyclic peptide compounds. Proc. Natl. Acad. Sci. USA 2015, 112, 5177–5182. [Google Scholar] [CrossRef] [Green Version]
- Lanigan-Gerdes, S.; Dooley, A.N.; Faull, K.F.; Lazazzera, B.A. Identification of subtilisin, Epr and Vpr as enzymes that produce CSF, an extracellular signalling peptide of Bacillus subtilis. Mol. Microbiol. 2007, 65, 1321–1333. [Google Scholar] [CrossRef]
- Pomerantsev, A.P.; Pomerantseva, O.M.; Camp, A.S.; Mukkamala, R.; Goldman, S.; Leppla, S.H. PapR peptide maturation: Role of the NprB protease in Bacillus cereus 569 PlcR/PapR global gene regulation. FEMS Immunol. Med. Microbiol. 2009, 55, 361–377. [Google Scholar] [CrossRef]
- Clewell, D.B.; Weaver, K.E.; Dunny, G.M.; Coque, T.M.; Francia, M.V.; Hayes, F. Extrachromosomal and Mobile Elements in Enterococci: Transmission, Maintenance, and Epidemiology. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Varahan, S.; Harms, N.; Gilmore, M.S.; Tomich, J.M.; Hancock, L.E. An ABC transporter is required for secretion of peptide sex pheromones in Enterococcus faecalis. MBio 2014, 5, e01726-14. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.; Guillot, A.; Wessner, F.; Algaron, F.; Besset, C.; Courtin, P.; Gardan, R.; Monnet, V. Control of the transcription of a short gene encoding a cyclic peptide in Streptococcus thermophilus: A new quorum-sensing system? J. Bacteriol. 2007, 189, 8844–8854. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.C.; LaSarre, B.; Jimenez, J.C.; Aggarwal, C.; Federle, M.J. Two Group A streptococcal peptide pheromones act through opposing Rgg regulators to control biofilm development. PLoS Pathog. 2011, 7, e1002190. [Google Scholar] [CrossRef]
- Chang, J.C.; Jimenez, J.C.; Federle, M.J. Induction of a quorum sensing pathway by environmental signals enhances group A streptococcal resistance to lysozyme. Mol. Microbiol. 2015, 97, 1097–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junges, R.; Salvadori, G.; Shekhar, S.; Amdal, H.A.; Periselneris, J.N.; Chen, T.; Brown, J.S.; Petersen, F.C. A Quorum-Sensing System That Regulates Streptococcus pneumoniae Biofilm Formation and Surface Polysaccharide Production. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Pascual, D.; Gaudu, P.; Fleuchot, B.; Besset, C.; Rosinski-Chupin, I.; Guillot, A.; Monnet, V.; Gardan, R. RovS and its associated signaling peptide form a cell-to-cell communication system required for Streptococcus agalactiae pathogenesis. MBio 2015, 6, e02306–e02314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, X.; Abdullah, I.T.; Gazioglu, O.; Manzoor, I.; Shafeeq, S.; Kuipers, O.P.; Hiller, N.L.; Andrew, P.W.; Yesilkaya, H. Rgg-Shp regulators are important for pneumococcal colonization and invasion through their effect on mannose utilization and capsule synthesis. Sci. Rep. 2018, 8, 6369. [Google Scholar] [CrossRef] [PubMed]
- Mignolet, J.; Fontaine, L.; Sass, A.; Nannan, C.; Mahillon, J.; Coenye, T.; Hols, P. Circuitry rewiring directly couples competence to predation in the gut dweller Streptococcus salivarius. Cell Rep. 2018, 22, 1627–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleuchot, B.; Gitton, C.; Guillot, A.; Vidic, J.; Nicolas, P.; Besset, C.; Fontaine, L.; Hols, P.; Leblond-Bourget, N.; Monnet, V.; et al. Rgg proteins associated with internalized small hydrophobic peptides: A new quorum-sensing mechanism in streptococci. Mol. Microbiol. 2011, 80, 1102–1119. [Google Scholar] [CrossRef]
- Gardan, R.; Besset, C.; Gitton, C.; Guillot, A.; Fontaine, L.; Hols, P.; Monnet, V. The extracellular life cycle of ComS, the competence stimulating peptide of Streptococcus thermophilus. J. Bacteriol. 2013, 195, 1845–1855. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.; Rukke, H.V.; Ricomini Filho, A.P.; Fimland, G.; Arntzen, M.O.; Thiede, B.; Petersen, F.C. Extracellular identification of a processed type II ComR/ComS pheromone of Streptococcus mutans. J. Bacteriol. 2012, 194, 3781–3788. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.C.; Federle, M.J. PptAB exports Rgg quorum-sensing peptides in Streptococcus. PLoS ONE 2016, 11, e0168461. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, L.; Boutry, C.; de Frahan, M.H.; Delplace, B.; Fremaux, C.; Horvath, P.; Boyaval, P.; Hols, P. A novel pheromone quorum-sensing system controls the development of natural competence in Streptococcus thermophilus and Streptococcus salivarius. J. Bacteriol. 2010, 192, 1444–1454. [Google Scholar] [CrossRef] [Green Version]
- Schramma, K.R.; Bushin, L.B.; Seyedsayamdost, M.R. Structure and biosynthesis of a macrocyclic peptide containing an unprecedented lysine-to-tryptophan crosslink. Nat. Chem. 2015, 7, 431–437. [Google Scholar] [CrossRef]
- Fleuchot, B.; Guillot, A.; Mézange, C.; Besset, C.; Chambellon, E.; Monnet, V.; Gardan, R. Rgg-associated SHP signaling peptides mediate cross-talk in streptococci. PLoS ONE 2013, 8, e66042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letort, C.; Juillard, V. Development of a minimal chemically-defined medium for the exponential growth of Streptococcus thermophilus. J. Appl. Microbiol. 2001, 91, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Gardan, R.; Besset, C.; Guillot, A.; Gitton, C.; Monnet, V. The oligopeptide transport system is essential for the development of natural competence in Streptococcus thermophilus strain LMD-9. J. Bacteriol. 2009, 191, 4647–4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, I.; Gruss, A.; Ehrlich, S.D.; Maguin, E. High-efficiency gene inactivation and replacement system for gram-positive bacteria. J. Bacteriol. 1993, 175, 3628–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Débarbouillé, M.; Arnaud, M.; Fouet, A.; Klier, A.; Rapoport, G. The sacT gene regulating the sacPA operon in Bacillus subtilis shares strong homology with transcriptional antiterminators. J. Bacteriol. 1990, 172, 3966–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, P.; Bize, L.; Muri, F.; Hoebeke, M.; Rodolphe, F.; Ehrlich, S.D.; Prum, B.; Bessières, P. Mining Bacillus subtilis chromosome heterogeneities using hidden Markov models. Nucleic Acids Res. 2002, 30, 1418–1426. [Google Scholar] [CrossRef]
- Ibrahim, M.; Nicolas, P.; Bessières, P.; Bolotin, A.; Monnet, V.; Gardan, R. A genome-wide survey of short coding sequences in streptococci. Microbiology 2007, 153, 3631–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutry, C.; Wahl, A.; Delplace, B.; Clippe, A.; Fontaine, L.; Hols, P. Adaptor protein MecA is a negative regulator of the expression of late competence genes in Streptococcus thermophilus. J. Bacteriol. 2012, 194, 1777–1788. [Google Scholar] [CrossRef] [Green Version]
- Thévenard, B.; Besset, C.; Choinard, S.; Fourcassié, P.; Boyaval, P.; Monnet, V.; Rul, F. Response of S. thermophilus LMD-9 to bacitracin: Involvement of a BceRS/AB-like module and of the rhamnose-glucose polysaccharide synthesis pathway. Int. J. Food Microbiol. 2014, 177, 89–97. [Google Scholar] [CrossRef]
- Schilcher, K.; Caesar, L.K.; Cech, N.B.; Horswill, A.R. Processing, export, and identification of novel linear peptides from Staphylococcus aureus. mBio 2020, 11, e00112–e00120. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Lv, H.; Yao, Y.; Cheng, S.; Huang, Q.; Wang, H.; Liu, X.; Bae, T.; Li, M.; Liu, Q. Roles of the Site 2 Protease Eep in Staphylococcus aureus. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Ye, J.; Rawson, R.B.; Goldstein, J.L. Regulated intramembrane proteolysis: A control mechanism conserved from bacteria to humans. Cell 2000, 100, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Quentin, Y.; Fichant, G.; Claverys, J.P. Independent evolution of competence regulatory cascades in streptococci? Trends Microbiol. 2006, 14, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Medlin, J.S.; Nguyen, D.R.; Disbennett, W.M.; Dawid, S. Molecular determinants of substrate selectivity of a pneumococcal Rgg-regulated peptidase-containing ABC transporter. mBio 2020, 11, e02502–e02519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junges, R.; Sturod, K.; Salvadori, G.; Amdal, H.A.; Chen, T.; Petersen, F.C. Characterization of a signaling system in Streptococcus mitis that mediates interspecies communication with Streptococcus pneumoniae. Appl. Environ. Microbiol. 2019, 85, e02297-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, A.; Borges, F.; Thibessard, A.; Gintz, B.; Decaris, B.; Leblond-Bourget, N. Characterisation of Streptococcus thermophilus CNRZ368 oxidative stress-resistant mutants: Involvement of a potential Rgg-like transcriptional regulator. Le Lait 2004, 84, 77–85. [Google Scholar] [CrossRef]
- Aggarwal, C.; Jimenez, J.C.; Nanavati, D.; Federle, M.J. Multiple length peptide-pheromone variants produced by Streptococcus pyogenes directly bind Rgg proteins to confer transcriptional regulation. J. Biol. Chem. 2014, 289, 22427–22436. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Resistance a | Description b | Source or Reference |
|---|---|---|---|---|
| LMD-9 | wild-type | [34] | ||
| CB001 | blp::PcomX-luxAB | [31] | ||
| TIL773 | Δeep | [27] | ||
| TIL883 | ΔamiCDE | [20] | ||
| TIL1038 | blp::Pshp1299-luxAB aphA3 | Km | [33] | |
| TIL1165 | blp::Pshp1358-luxAB | [27] | ||
| TIL1179 | pBV5030::P32 | Er | [28] | |
| TIL1213 | ΔamiCDE pBV5030::P32-shp1358 | Er | [27] | |
| TIL1486 | pptAB::erm | Er | PCR fragment pptAB::erm → LMD-9 | This study |
| TIL1488 | blp::PcomX-luxABpptAB::erm | Er | TIL1486 DNA → CB001 | This study |
| TIL1489 | blp::Pshp1358-luxAB pptAB::erm | Er | TIL1486 DNA → TIL1165 | This study |
| TIL1491 | blp::Pshp1299-luxAB aphA3 pptAB::erm | Er Km | TIL1486 DNA → TIL1038 | This study |
| TIL1557 | blp::PpptAB-luxAB aphA3 | Km | pGICB004a::PpptAB → LMD-9 | This study |
| TIL1558 | ΔpptAB | pG+host9::updown.pptAB → LMD-9 | This study | |
| TIL1559 | pBV5030::P32-shp1358 | Er | pBV5030::P32-shp1358 → LMD-9 | This study |
| TIL1560 | comR::aphA3 | Km | pG+host9::updown.comRaphA3 → LMD-9 | This study |
| TIL1561 | Δeep comR::aphA3 | Km | pG+host9::updown.comRaphA3 → TIL773 | This study |
| TIL1562 | ΔpptAB comR::aphA3 | Km | pG+host9::updown.comRaphA3 → TIL1558 | This study |
| TIL1563 | ΔpptAB pBV5030::P32-shp1358 | Er | pBV5030::P32-shp1358 → TIL1558 | This study |
| TIL1564 | ΔamiCDE comR::aphA3 | Km | pG+host9::updown.comRaphA3. → TIL883 | This study |
| TIL1565 | ΔpptAB pBV5030::P32 | Er | pBV5030::P32 → TIL1558 | This study |
| Locus Tag | Old Locus Tag | Fold Change | TIL1560 * | TIL1562 * | Start | Stop | Strand | Gene Name | Protein Length | Description |
|---|---|---|---|---|---|---|---|---|---|---|
| Cluster STER_RS04380-STER_RS04390 | ||||||||||
| STER_RS04380 | STER_0885 | 3.6 | 2552 | 704 | 817570 | 818481 | + | 303 | cysteine synthase family protein | |
| STER_RS04385 | STER_0886 | 3.8 | 3818 | 1006 | 818503 | 819687 | + | 394 | aminotransferase class V-fold PLP-dependent enzyme | |
| STER_RS04390 | STER_0887 | 3.9 | 1704 | 433 | 819629 | 820210 | + | 193 | serine acetyltransferase | |
| Locus sCDS_265/Ster_RS04620 | ||||||||||
| STER_sCDS_265 | STER_0933 | NR | 20 | 7 | 864863 | 864934 | − | shp265 | 23 | |
| STER_RS04620 | STER_0934 | NR | 1214 | 1296 | 865021 | 865878 | + | rgg4620 | 285 | MutR family transcriptional regulator |
| STER_RS04625 | STER_0935 | 3.0 | 3966 | 1322 | 865938 | 866738 | + | 266 | Fe-S oxidoreductase | |
| STER_sCDS_510 | 3.8 | 990 | 259 | 866798 | 866947 | + | 49 | |||
| STER_sCDS_661 | 2.5 | 351 | 141 | 867030 | 867119 | + | 29 | |||
| STER_RS04630 | STER_0937 | 2.7 | 3087 | 1132 | 867149 | 868246 | + | 365 | radical SAM protein | |
| STER_RS04635 | STER_0938 | 2.8 | 1748 | 629 | 868243 | 868986 | + | 247 | arginase family protein | |
| STER_RS04640 | STER_0939 | 2.2 | 3534 | 1641 | 868993 | 870165 | + | 390 | MFS transporter | |
| Locus shp/rgg1299 | ||||||||||
| STER_RS06360 | STER_1290 | 2.0 | 601 | 296 | 1198824 | 1199426 | − | rr06 | 200 | response regulator transcription factor |
| STER_RS06365 | STER_1291 | 1.9 | 1125 | 593 | 1199423 | 1200517 | − | hk06 | 364 | sensor histidine kinase |
| STER_RS06370 | STER_1292 | 11.2 | 504 | 45 | 1200510 | 1201247 | − | 245 | ABC transporter permease | |
| STER_RS06375 | STER_1293 | 12.8 | 545 | 43 | 1201244 | 1202125 | − | 293 | MULTISPECIES: ABC transporter ATP-binding protein | |
| STER_RS06380 | STER_1294 | 13.3 | 111 | 8 | 1202118 | 1202297 | − | 59 | hypothetical protein | |
| STER_RS06385 | STER_1295 | 18.7 | 183 | 10 | 1202308 | 1202517 | − | 69 | hypothetical protein | |
| STER_RS06390 | ND | ND | ND | 1202668 | 1202934 | − | 87 | hypothetical protein (incomplete) | ||
| STER_RS10535 | ND | ND | ND | 1202925 | 1203152 | − | 74 | hypothetical protein (incomplete) | ||
| STER_sCDS_172 | 56.6 | 8877 | 157 | 1203195 | 1203290 | − | 31 | |||
| STER_RS06395 | STER_1296 | 59.7 | 93273 | 1563 | 1203277 | 1204506 | − | 409 | MFS transporter permease | |
| STER_RS06400 | STER_1297 | 49.7 | 195871 | 3942 | 1204484 | 1206817 | − | 777 | oligoendopeptidase F | |
| STER_sCDS_398 | 46.3 | 97 | 2 | 1206867 | 1206920 | − | 17 | |||
| STER_sCDS_310 | NR | 1 | 1 | 1206895 | 1206960 | − | 21 | |||
| STER_sCDS_714 | STER_1298 | 67.4 | 1015 | 15 | 1207133 | 1207213 | + | shp1299 | 26 | |
| STER_RS06405 | STER_1299 | NR | 924 | 1060 | 1207191 | 1208066 | − | rgg1299 | 291 | XRE family transcriptional regulator |
| Locus shp/rgg1358 | ||||||||||
| STER_sCDS_566 | 9.8 | 76 | 8 | 1255722 | 1255835 | + | 37 | |||
| STER_sCDS_613 | NR | 25 | 31 | 1256064 | 1256165 | + | 33 | |||
| STER_sCDS_543 | NR | 26 | 14 | 1256228 | 1256353 | + | 41 | |||
| STER_sCDS_851 | NR | 0 | 0 | 1256331 | 1256390 | + | 19 | |||
| STER_RS06660 | STER_1350 | 3.1 | 137 | 44 | 1256335 | 1256604 | + | 89 | hypothetical protein | |
| STER_RS06665 | STER_1351 | NR | 55 | 70 | 1256764 | 1257063 | + | 99 | hypothetical protein | |
| STER_RS06670 | STER_1352 | 114.6 | 32175 | 281 | 1257163 | 1258074 | − | 303 | sodium transporter | |
| STER_RS06675 | ND | ND | ND | 1258071 | 1258393 | − | 107 | hypothetical protein (incomplete) | ||
| STER_RS06680 | 132.9 | 21819 | 164 | 1258390 | 1258992 | − | 200 | SPASM domain-containing protein | ||
| STER_RS06685 | STER_1355 | 113.3 | 31202 | 275 | 1259086 | 1260177 | − | strC | 363 | MULTISPECIES: transporter |
| STER_RS06690 | STER_1356 | 109.6 | 53740 | 490 | 1260174 | 1261493 | − | strB | 439 | KxxxW cyclic peptide radical SAM maturase |
| STER_RS10575 | STER_1357 | 78.0 | 6544 | 84 | 1261563 | 1261655 | − | strA | 30 | KxxxW-cyclized peptide pheromone |
| STER_RS06695 | STER_1358 | NR | 450 | 565 | 1261738 | 1262598 | − | rgg1358 | 286 | MutR family transcriptional regulator |
| STER_sCDS_768 | 114.8 | 723 | 6 | 1262689 | 1262760 | + | shp1358 | 23 | ||
| Locus sCDS_273/Ster_RS07530 | ||||||||||
| STER_sCDS_273 | NR | 3 | 8 | 1434537 | 1434608 | − | shp273 | 23 | ||
| STER_RS07530 | STER_1530 | NR | 648 | 628 | 1434695 | 1435558 | + | rgg7530 | 287 | MutR family transcriptional regulator |
| STER_RS07535 | STER_1531 | 28.9 | 19052 | 660 | 1435675 | 1436853 | + | 392 | MFS transporter | |
| Gene STER_sCDS_560 | ||||||||||
| STER_sCDS_560 | 3.2 | 39 | 12 | 1441243 | 1441359 | + | 38 | |||
| pptAB | ||||||||||
| STER_RS07725 | STER_1571 | 37.9 | 5837 | 154 | 1472993 | 1474030 | − | pptB | 345 | ABC transporter permease |
| STER_RS07730 | STER_1572 | 2491 | 3491 | 1 | 1474027 | 1474758 | − | pptA | 243 | ABC transporter ATP-binding protein |
| Locus sCDS_279/Ster_RS09420 | ||||||||||
| STER_sCDS_279 | 1.9 | 139 | 74 | 1785690 | 1785761 | − | shp279 | 23 | ||
| STER_RS09420 | NR | 1102 | 1194 | 1786014 | 1786712 | + | rgg9420 | 232 | MutR family transcriptional regulator | |
| STER_sCDS_584 | NR | 71 | 92 | 1786790 | 1786897 | + | 35 | |||
| STER_sCDS_809 | NR | 0 | 0 | 1786854 | 1786919 | + | 21 | |||
| STER_sCDS_462 | 13.1 | 1242 | 95 | 1786909 | 1787088 | + | 59 | |||
| STER_RS09425 | STER_1924 | 14.9 | 15002 | 1007 | 1787097 | 1788152 | + | 351 | radical SAM protein | |
| STER_RS09430 | STER_1925 | 15.4 | 18315 | 1192 | 1788145 | 1789686 | + | 513 | ABC transporter ATP-binding protein | |
| STER_sCDS_680 | 19.6 | 150 | 8 | 1789819 | 1789905 | + | 28 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lingeswaran, A.; Metton, C.; Henry, C.; Monnet, V.; Juillard, V.; Gardan, R. Export of Rgg Quorum Sensing Peptides is Mediated by the PptAB ABC Transporter in Streptococcus Thermophilus Strain LMD-9. Genes 2020, 11, 1096. https://doi.org/10.3390/genes11091096
Lingeswaran A, Metton C, Henry C, Monnet V, Juillard V, Gardan R. Export of Rgg Quorum Sensing Peptides is Mediated by the PptAB ABC Transporter in Streptococcus Thermophilus Strain LMD-9. Genes. 2020; 11(9):1096. https://doi.org/10.3390/genes11091096
Chicago/Turabian StyleLingeswaran, Abarna, Coralie Metton, Céline Henry, Véronique Monnet, Vincent Juillard, and Rozenn Gardan. 2020. "Export of Rgg Quorum Sensing Peptides is Mediated by the PptAB ABC Transporter in Streptococcus Thermophilus Strain LMD-9" Genes 11, no. 9: 1096. https://doi.org/10.3390/genes11091096