Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. The Origin of Primary Human Mesenchymal Stem Cells
2.2. Adipogenesis of Human MSC
2.3. Fatty Acids Preparation [25 mM]
2.4. Genetic Material Extraction
2.5. Gene Expression
2.6. Glucose Uptake
2.7. Adipokine Levels
2.8. Assessment of the Accumulated Lipids
2.9. Assessment of the Size of Adipocytes
2.10. Statistical Analysis
3. Results
3.1. Adipogenesis of Stem Cells with the Addition of Fatty Acids
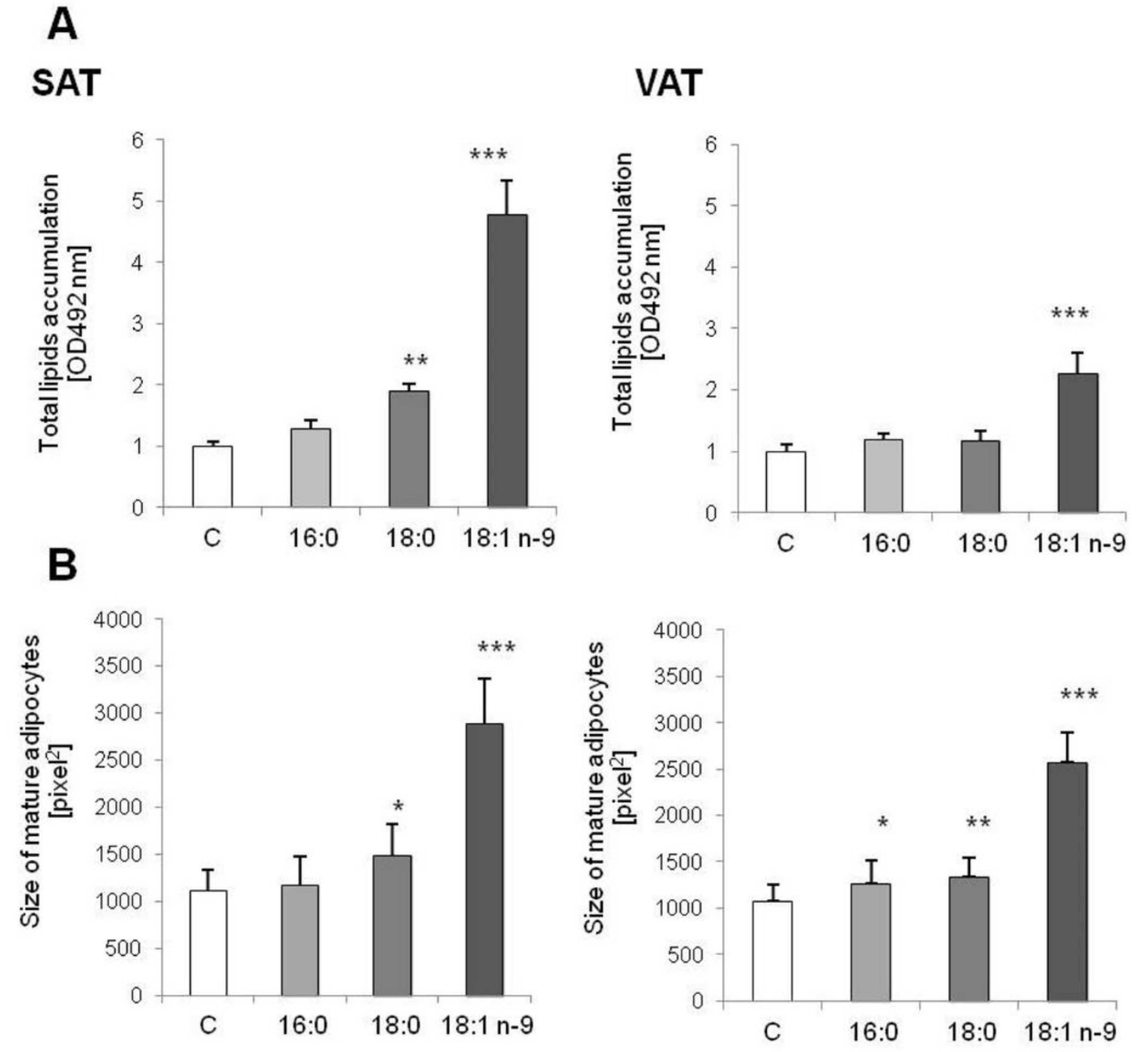
3.1.1. Morphological Changes and Lipid Accumulation
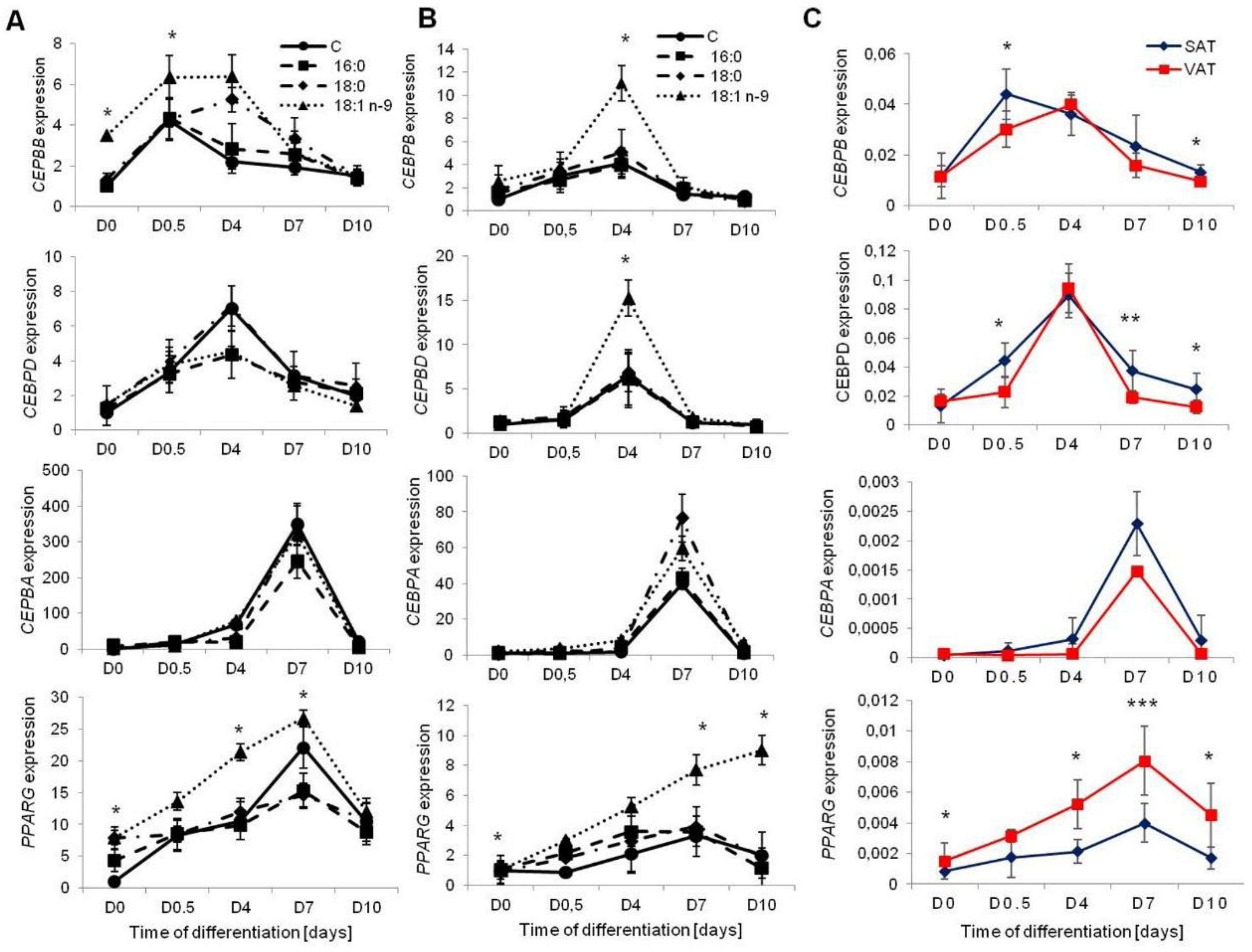
3.1.2. Transcription Factors Regulating Adipogenesis
3.2. The Phenotype of Mature Adipocytes
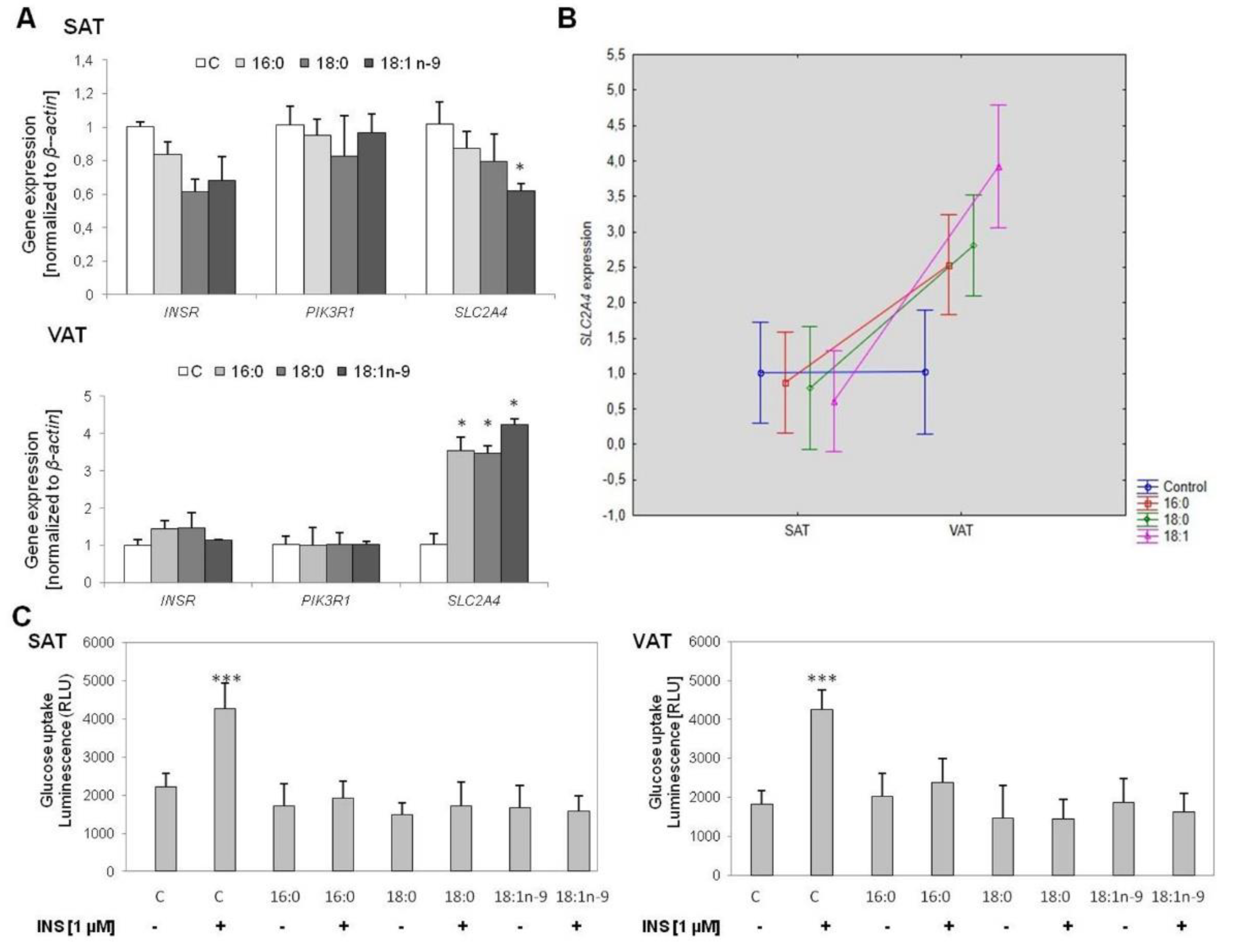
3.2.1. Insulin Signaling Pathway
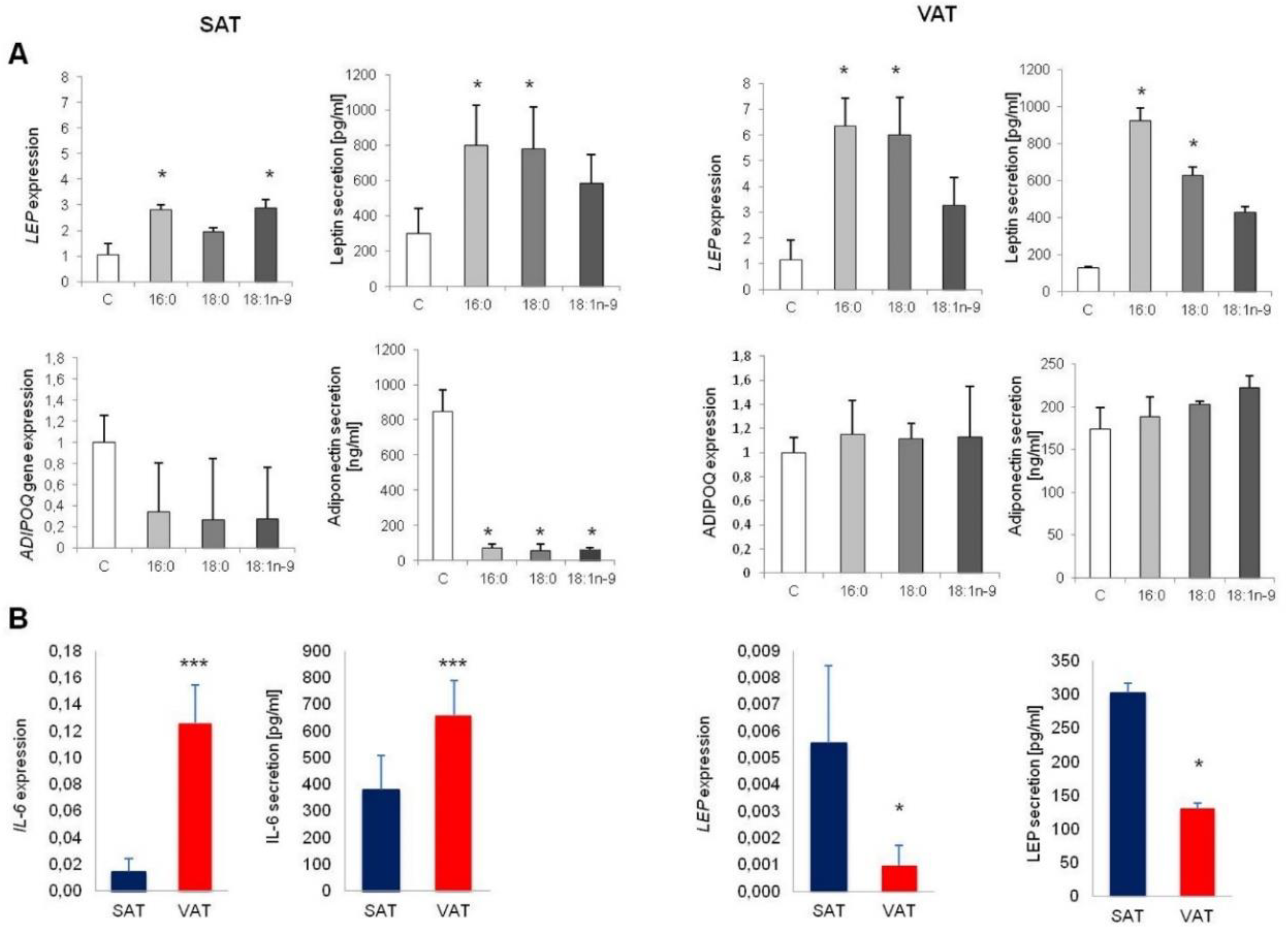
3.2.2. Cytokine and Adipokine Expression and Secretion
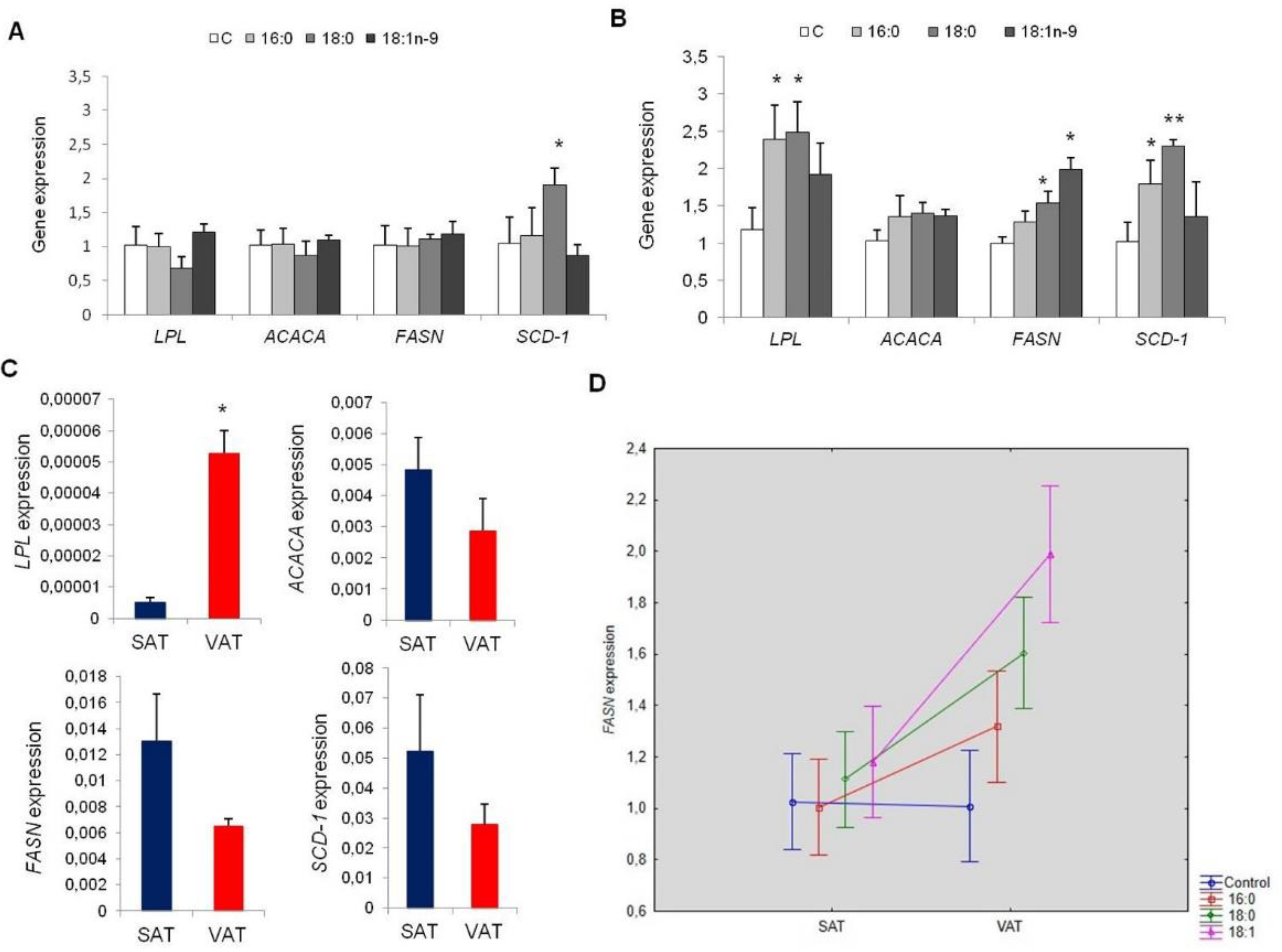
3.2.3. Lipid Metabolism
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Engin, A. The Definition and prevalence of obesity and metabolic syndrome. In Obesity and Lipotoxicity; Engin, A.B., Engin, A., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–17. ISBN 978-3-319-48382-5. [Google Scholar]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Joe, A.W.B.; Yi, L.; Even, Y.; Vogl, A.W.; Rossi, F.M.V. Depot-specific differences in adipogenic progenitor abundance and proliferative response to high-fat diet. Stem Cells 2009, 27, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Dam, V.; Sikder, T.; Santosa, S. From neutrophils to macrophages: Differences in regional adipose tissue depots. Obes. Rev. 2016, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Andersson, D.P.; Thörne, A.; Wirén, M.; Hoffstedt, J.; Näslund, E.; Thorell, A.; Rydén, M. Variations in the size of the major omentum are primarily determined by fat cell number. J. Clin. Endocrinol. Metab. 2013, 98, E897–E901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, R.; Metcalf, M.; White, A.; Madala, A.; Winters, B.; Maizlin, I.; Jobe, B.; Roberts, C.; Slifka, M.; Marks, D. Depot-specific differences in inflammatory mediators and a role for NK cells and IFN-γ in inflammation in human adipose tissue. Int. J. Obes. 2009, 33, 978–990. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [Green Version]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef]
- Regassa, A.; Kim, W.K. Effects of oleic acid and chicken serum on the expression of adipogenic transcription factors and adipogenic differentiation in hen preadipocytes. Cell Biol. Int. 2013, 37, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Hamilton, J.A.; Kirkland, J.L.; Corkey, B.E.; Guo, W. Oleate-induced formation of fat cells with impaired insulin sensivitity. Lipids 2006, 41, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Malodobra-Mazur, M.; Cierzniak, A.; Dobosz, T. Oleic acid influences the adipogenesis of 3T3-L1 cells via DNA Methylation and may predispose to obesity and obesity-related disorders. Lipids Health Dis. 2019, 18, 230. [Google Scholar] [CrossRef] [Green Version]
- Hilgendorf, K.I.; Johnson, C.T.; Mezger, A.; Rice, S.L.; Norris, A.M.; Demeter, J.; Greenleaf, W.J.; Reiter, J.F.; Kopinke, D.; Jackson, P.K. Omega-3 fatty acids activate ciliary FFAR4 to control adipogenesis. Cell 2019, 179, 1289–1305.e21. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Kyrou, I.; Tan, B.K.; Dimitriadis, G.K.; Ramanjaneya, M.; Tripathi, G.; Patel, V.; James, S.; Kawan, M.; Chen, J.; et al. Short-chain fatty acid acetate stimulates adipogenesis and mitochondrial biogenesis via GPR43 in brown adipocytes. Endocrinology 2016, 157, 1881–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krey, G.; Braissant, O.; L’Horset, F.; Kalkhoven, E.; Perroud, M.; Parker, M.G.; Wahli, W. Fatty acids, eicosanoids, and hypolipidemic agents identified as ligands of peroxisome proliferator-activated receptors by coactivator-dependent receptor ligand assay. Mol. Endocrinol. 1997, 11, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Legrand-Poels, S.; Esser, N.; L’homme, L.; Scheen, A.; Paquot, N.; Piette, J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem. Pharmacol. 2014, 92, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.; Martinez, K.; Chuang, C.-C.; LaPoint, K.; McIntosh, M. Saturated fatty acid-mediated inflammation and insulin resistance in adipose tissue: Mechanisms of action and implications. J. Nutr. 2009, 139, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Koska, J.; Ozias, M.K.; Deer, J.; Kurtz, J.; Salbe, A.D.; Harman, S.M.; Reaven, P.D. A human model of dietary saturated fatty acid induced insulin resistance. Metabolism 2016, 65, 1621–1628. [Google Scholar] [CrossRef]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; McAllan, L.; Nilaweera, K.N.; et al. Monounsaturated fatty acid–enriched high-fat diets impede adipose NLRP3 inflammasome–mediated IL-1β secretion and insulin resistance despite obesity. Diabetes 2015, 64, 2116–2128. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamaguchi, S.; Shimoyama, T.; Seki, H.; Miyokawa, K.; Katsuta, H.; Tanaka, T.; Yoshimoto, K.; Ohno, H.; Nagamatsu, S.; et al. JNK- and IκB-dependent pathways regulate MCP-1 but not adiponectin release from artificially hypertrophied 3T3-L1 adipocytes preloaded with palmitate in vitro. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E898–E909. [Google Scholar] [CrossRef]
- Hemmrich, K.; von Heimburg, D.; Cierpka, K.; Haydarlioglu, S.; Pallua, N. Optimization of the differentiation of human preadipocytes in vitro. Differentiation 2005, 73, 28–35. [Google Scholar] [CrossRef]
- Yanting, C.; Yang, Q.Y.; Ma, G.L.; Du, M.; Harrison, J.H.; Block, E. Dose- and type-dependent effects of long-chain fatty acids on adipogenesis and lipogenesis of bovine adipocytes. J. Dairy Sci. 2018, 101, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, T.; Yonekura, S.; Kobayashi, Y.; Hagino, A.; Katoh, K.; Obara, Y. Effects of long-chain fatty acids on cytosolic triacylglycerol accumulation and lipid droplet formation in primary cultured bovine mammary epithelial cells. J. Dairy Sci. 2004, 87, 2527–2534. [Google Scholar] [CrossRef] [Green Version]
- Lauvai, J.; Becker, A.-K.; Lehnert, K.; Schumacher, M.; Hieronimus, B.; Vetter, W.; Graeve, L. The Furan Fatty Acid 9M5 Acts as a partial ligand to peroxisome proliferator-activated receptor γ and enhances adipogenesis in 3T3-L1 preadipocytes. Lipids 2019, 54, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Terauchi, Y.; Miki, H.; Tamemoto, H.; Yamauchi, T.; Komeda, K.; Satoh, S.; Nakano, R.; Ishii, C.; Sugiyama, T.; et al. PPAR γ mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol. Cell 1999, 4, 597–609. [Google Scholar] [CrossRef]
- Malodobra-Mazur, M.; Dziewulska, A.; Kozinski, K.; Dobrzyn, P.; Kolczynska, K.; Janikiewicz, J.; Dobrzyn, A. Stearoyl-CoA desaturase regulates inflammatory gene expression by changing DNA methylation level in 3T3 adipocytes. Int. J. Biochem. Cell Biol. 2014, 55, 40–50. [Google Scholar] [CrossRef]
- Peirce, V.; Carobbio, S.; Vidal-Puig, A. The different shades of fat. Nature 2014, 510, 76–83. [Google Scholar] [CrossRef]
- Drolet, R.; Richard, C.; Sniderman, A.D.; Mailloux, J.; Fortier, M.; Huot, C.; Rhéaume, C.; Tchernof, A. Hypertrophy and hyperplasia of abdominal adipose tissues in women. Int. J. Obes. 2008, 32, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Hernandez-Morante, J.J.; Lujan, J.; Tebar, F.J.; Zamora, S. Relationship between fat cell size and number and fatty acid composition in adipose tissue from different fat depots in overweight/obese humans. Int. J. Obes. 2006, 30, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Arner, P. Differences in lipolysis between human subcutaneous and omental adipose tissues. Ann. Med. 1995, 27, 435–438. [Google Scholar] [CrossRef]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chen, Y.; Xiong, Z.; Yu, S.; Zhou, B.; Ling, Y.; Zheng, Z.; Shi, G.; Wu, Y.; Qian, X. Ginsenoside Rb1 inhibits free fatty acids-induced oxidative stress and inflammation in 3T3-L1 adipocytes. Mol. Med. Rep. 2017, 16, 9165–9172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drolet, R.; Bélanger, C.; Fortier, M.; Huot, C.; Mailloux, J.; Légaré, D.; Tchernof, A. Fat depot-specific impact of visceral obesity on adipocyte adiponectin release in women. Obesity 2009, 17, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Van Harmelen, V.; Reynisdottir, S.; Eriksson, P.; Thörne, A.; Hoffstedt, J.; Lönnqvist, F.; Arner, P. Leptin secretion from subcutaneous and visceral adipose tissue in women. Diabetes 1998, 47, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Dong, L.Q. Adiponectin signaling and function in insulin target tissues. J. Mol. Cell Biol. 2016, 8, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence 5′→3′ | Exon * | Length [bp] | R2 ** |
|---|---|---|---|---|
| CEBPB | F: AGCACCACGACTTCCTCTC | 1 | 54 | 0.99 |
| R: AGTTCTTGCCCCCGTAGTC | ||||
| C/EBPδ | F: ATGTACGACGACGAGAGCG | 1 | 108 | 0.98 |
| R: GTTGAAGAGGTCGGCGAAG | ||||
| C/EBPα | F: GCCAAGAAGTCGGTGGACA | 1 | 150 | 0.98 |
| R: GCGGTCATTGTCACTGGTC | ||||
| PPARG | F: TAATGCCATCAGGTTTGGGC | 4/5 | 101 | 0.96 |
| R: GGTCAGCGGACTCTGGATT | ||||
| INSR | F: TTCGAGGAGGCAACAATCTG | 4/5 | 93 | 0.95 |
| R: CGTAGGATCGGCGGATTTTT | ||||
| PIK3R1 | F: TGCCTGCTCTGTAGTGGTG | 14/15 | 70 | 0.98 |
| R: GCCATAGCCAGTTGCTGTTT | ||||
| SLC2A4 | F: AGCAGCTCTCTGGCATCAAT | 8/9 | 139 | 0.99 |
| R: ACCAACAACACCGAGACCAA | ||||
| IL-6 | F: TCAATGAGGAGACTTGCCTG | 2/3 | 114 | 0.98 |
| R: GCACAGCTCTGGCTTGTTC | ||||
| IL-10 | F: GGACTTTAAGGGTTACCTGG | 2/3 | 95 | 0.99 |
| R: CTGGGTCTTGGTTCTCAGC | ||||
| ADIPOQ | F: GGAGATCCAGGTCTTATTGG | 1/2 | 184 | 0.99 |
| R: TGGGCATGTTGGGGATAGTA | ||||
| LEP | F: TTCACACACGCAGTCAGTCT | 1/2 | 134 | 0.98 |
| R: GCATACTGGTGAGGATCTGT | ||||
| LPL | F: TCACTCTGCCTGAAGTTTCC | 7/8 | 135 | 0.99 |
| R: TGCTCCACCAGTCTGACCA | ||||
| ACACA | F: CGTCCTCACCCAACCCAAA | 47/48 | 105 | 0.98 |
| R: TCTACCAACCACCACAGTCT | ||||
| FASN | F: GGCATCAATGTCCTGCTGAA | 4/5 | 107 | 0.99 |
| R: TACCCATTCCCCGCTGTGT | ||||
| SCD-1 | F: CCAGAGGAGGTACTACAAAC | 3/4 | 72 | 0.99 |
| R: AAATACCAGGGCACAAGCGT | ||||
| Β-actin | F: GAGAAGATGACCCAGATCA | 2/3 | 72 | 0.99 |
| R: TAGCACAGCCTGGATAGCAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Małodobra-Mazur, M.; Cierzniak, A.; Pawełka, D.; Kaliszewski, K.; Rudnicki, J.; Dobosz, T. Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids. Genes 2020, 11, 1092. https://doi.org/10.3390/genes11091092
Małodobra-Mazur M, Cierzniak A, Pawełka D, Kaliszewski K, Rudnicki J, Dobosz T. Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids. Genes. 2020; 11(9):1092. https://doi.org/10.3390/genes11091092
Chicago/Turabian StyleMałodobra-Mazur, Małgorzata, Aneta Cierzniak, Dorota Pawełka, Krzysztof Kaliszewski, Jerzy Rudnicki, and Tadeusz Dobosz. 2020. "Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids" Genes 11, no. 9: 1092. https://doi.org/10.3390/genes11091092