Transcriptome Analysis Reveals Inhibitory Effects of Lentogenic Newcastle Disease Virus on Cell Survival and Immune Function in Spleen of Commercial Layer Chicks

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Total RNA Isolation
2.3. cDNA Library Construction and Sequencing
2.4. Sequence Reads Quality Control, Mapping, and Counting
2.5. Differential Expression, Pathway, and Co-Expression Analyses
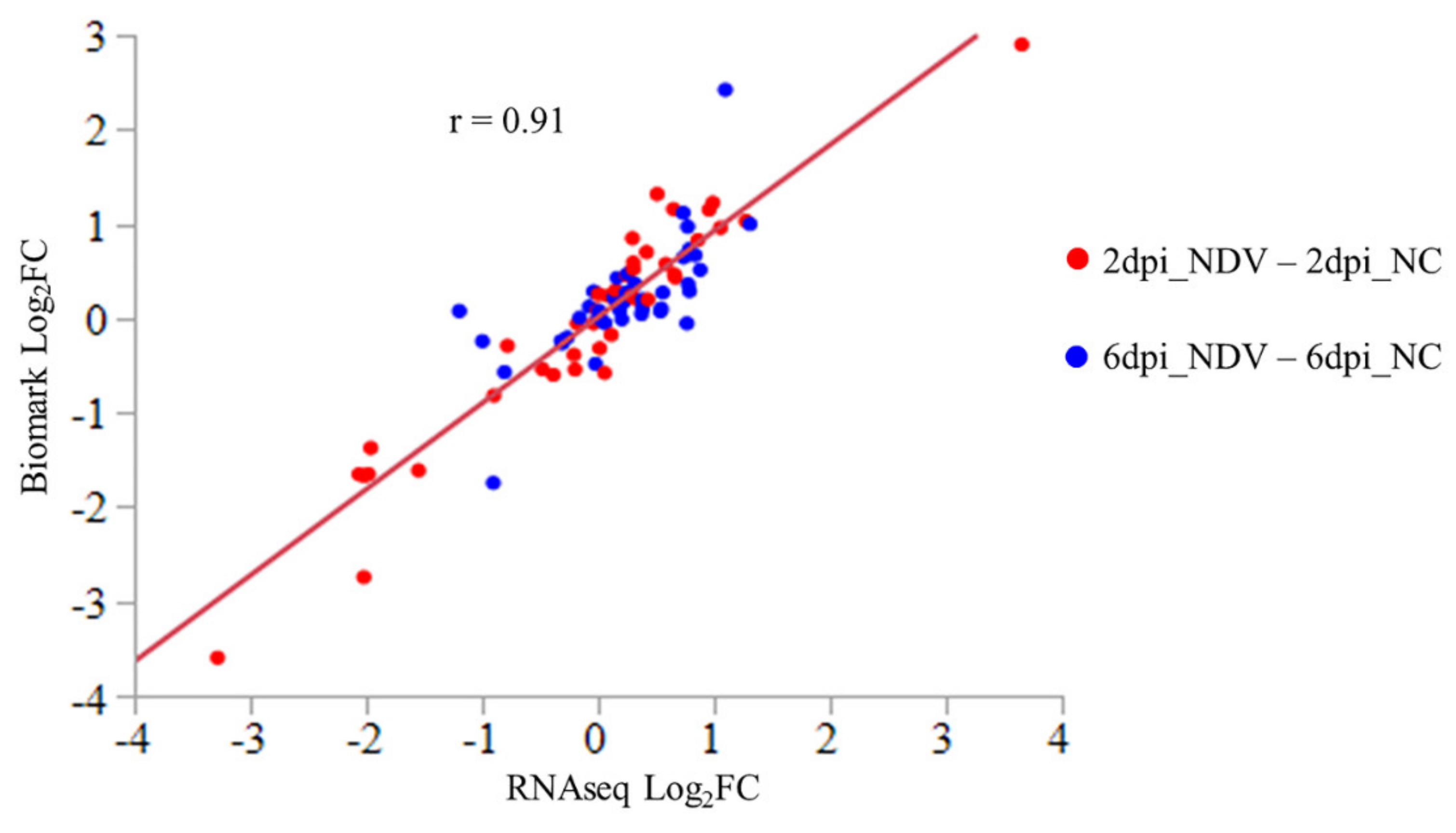
2.6. Fluidigm Biomark Assay of Gene Expression
2.7. Data Availability Statement
3. Results
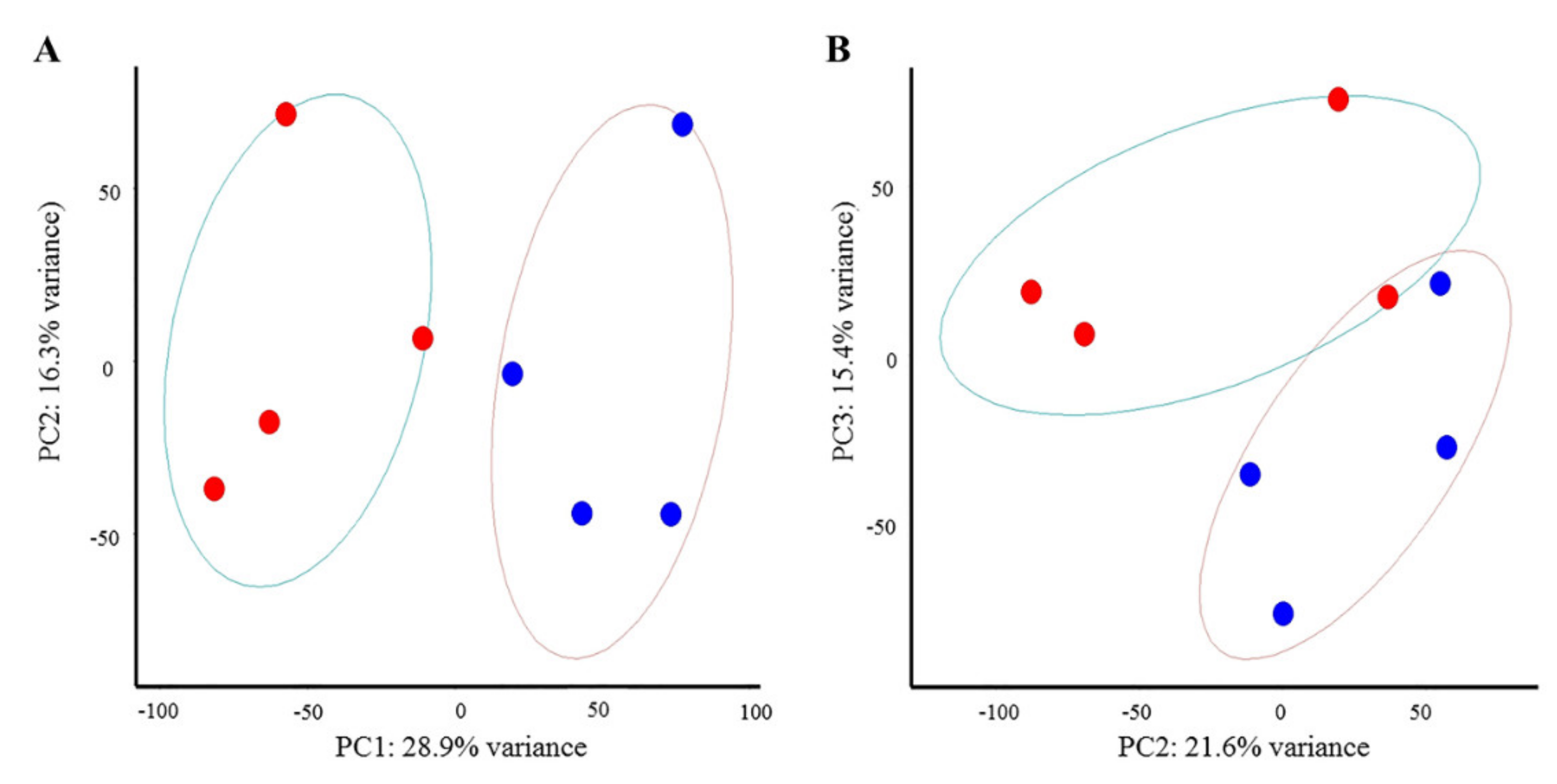
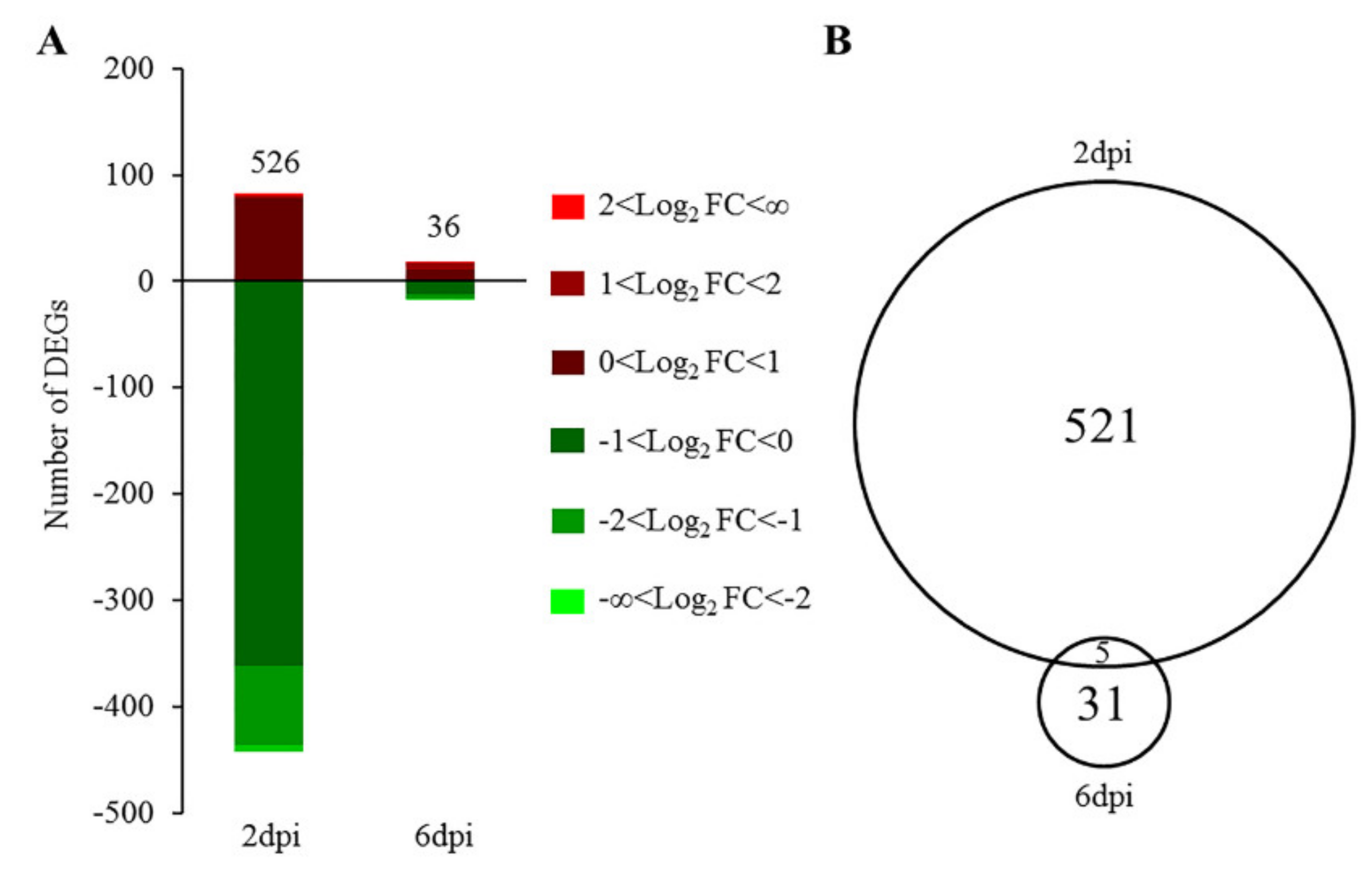
3.1. Gene Expression Changes Induced by NDV
3.2. Ingenuity Pathway Analysis of Differentially Expressed Genes
3.3. Comparison with Previous RNA-Seq and GWAS Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, C.; King, D.J.; Seal, B.S. Pathogenesis of Newcastle disease in chickens experimentally infected with viruses of different virulence. Vet. Pathol. 1999, 36, 125–132. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef]
- Gueye, E.F. The role of family poultry in poverty alleviation, food security and the promotion of gender equality in rural Africa. Outlook Agric. 2000, 29, 129–136. [Google Scholar] [CrossRef]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel Mechanisms Revealed in the Trachea Transcriptome of Resistant and Susceptible Chicken Lines following Infection with Newcastle Disease Virus. Clin. Vaccine Immunol. CVI 2017, 24. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.K.; Afify, M.A.; Aly, M.M. Genetic resistance of Egyptian chickens to infectious bursal disease and Newcastle disease. Trop. Anim. Health Prod. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- King, D.J. Influence of chicken breed on pathogenicity evaluation of velogenic neurotropic Newcastle disease virus isolates from cormorants and turkeys. Avian Dis. 1996, 40, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Rowland, K.; Wolc, A.; Gallardo, R.A.; Kelly, T.; Zhou, H.; Dekkers, J.C.M.; Lamont, S.J. Genetic Analysis of a Commercial Egg Laying Line Challenged With Newcastle Disease Virus. Front. Genet. 2018, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Kaiser, M.G.; Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.J.; Lamont, S.J. Transcriptome Analysis in Spleen Reveals Differential Regulation of Response to Newcastle Disease Virus in Two Chicken Lines. Sci. Rep. 2018, 8, 1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel analysis of the Harderian gland transcriptome response to Newcastle disease virus in two inbred chicken lines. Sci. Rep. 2018, 8, 6558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Dekkers, J.C.M.; Zhou, H.J.; Lamont, S.J. Resistant and susceptible chicken lines show distinctive responses to Newcastle disease virus infection in the lung transcriptome. BMC Genom. 2017, 18, 989. [Google Scholar] [CrossRef] [Green Version]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional Innate Immune Response of the Developing Chicken Embryo to Newcastle Disease Virus Infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Saelao, P.; Wang, Y.; Gallardo, R.A.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H.J. Novel insights into the host immune response of chicken Harderian gland tissue during Newcastle disease virus infection and heat treatment. BMC Vet. Res. 2018, 14, 280. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Gen. Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczynski, D.R.; Suarez, D.L.; King, D.J.; Miller, P.J.; Afonso, C.L. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef]
- Lan, D.; Tang, C.; Li, M.; Yue, H. Screening and identification of differentially expressed genes from chickens infected with Newcastle disease virus by suppression subtractive hybridization. Avian Pathol. 2010, 39, 151–159. [Google Scholar] [CrossRef]
- Saelao, P.; Wang, Y.; Chanthavixay, G.; Gallardo, R.A.; Wolc, A.; Dekkers, J.C.M.; Lamont, S.J.; Kelly, T.; Zhou, H. Genetics and Genomic Regions Affecting Response to Newcastle Disease Virus Infection under Heat Stress in Layer Chickens. Genes 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Walugembe, M.; Mushi, J.R.; Amuzu-Aweh, E.N.; Chiwanga, G.H.; Msoffe, P.L.; Wang, Y.; Saelao, P.; Kelly, T.; Gallardo, R.A.; Zhou, H.; et al. Genetic Analyses of Tanzanian Local Chicken Ecotypes Challenged with Newcastle Disease Virus. Genes 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, N.; King, D.J.; Kapczynski, D.R.; Seal, B.S.; Brown, C.C. Experimental pathogenesis for chickens, turkeys, and pigeons of exotic Newcastle disease virus from an outbreak in California during 2002–2003. Vet. Pathol. 2006, 43, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Veits, J.; Wiesner, D.; Fuchs, W.; Hoffmann, B.; Granzow, H.; Starick, E.; Mundt, E.; Schirrmeier, H.; Mebatsion, T.; Mettenleiter, T.C.; et al. Newcastle disease virus expressing H5 hemagglutinin gene protects chickens against Newcastle disease and avian influenza. Proc. Natl. Acad. Sci. USA 2006, 103, 8197–8202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annapragada, A.; Borgerson, C.; Iams, S.; Ravelomanantsoa, M.A.; Crawford, G.C.; Helin, M.; Anjaranirina, E.J.G.; Randriamady, H.J.; Golden, C.D. Modeling the Impact of Newcastle Disease Virus Vaccinations on Chicken Production Systems in Northeastern Madagascar. Front. Vet. Sci. 2019, 6, 305. [Google Scholar] [CrossRef] [PubMed]
- Calderon, N.L.; Galundo-Muniz, F.; Ortiz, M.; Lomniczi, B.; Fehervari, T.; Paasch, L.H. Thrombocytopenia in Newcastle disease: Haematological evaluation and histological study of bone marrow. Acta Vet. Hung. 2005, 53, 507–513. [Google Scholar] [CrossRef]
- Ojok, L.; Brown, C. An immunohistochemical study of the pathogenesis of virulent viscerotropic Newcastle disease in chickens. J. Comp. Pathol. 1996, 115, 221–227. [Google Scholar] [CrossRef]
- Etriwati Ratih, D.; Handharyani, E.; Setiyaningsih, S. Pathology and immunohistochemistry study of Newcastle disease field case in chicken in Indonesia. Vet. World 2017, 10, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Lam, K.M.; Kabbur, M.B.; Eiserich, J.P. Newcastle disease virus-induced functional impairments and biochemical changes in chicken heterophils. Vet. Immunol. Immunopathol. 1996, 53, 313–327. [Google Scholar] [CrossRef]
- Lori, A.; Perrotta, M.; Lembo, G.; Carnevale, D. The Spleen: A Hub Connecting Nervous and Immune Systems in Cardiovascular and Metabolic Diseases. Int. J. Mol. Sci. 2017, 18, 1216. [Google Scholar] [CrossRef]
- Wu, H.; Kumar, A.; Miao, H.; Holden-Wiltse, J.; Mosmann, T.R.; Livingstone, A.M.; Belz, G.T.; Perelson, A.S.; Zand, M.S.; Topham, D.J. Modeling of influenza-specific CD8+ T cells during the primary response indicates that the spleen is a major source of effectors. J. Immunol. 2011, 187, 4474–4482. [Google Scholar] [CrossRef] [Green Version]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef]
- Venable, J.D.; Ameriks, M.K.; Blevitt, J.M.; Thurmond, R.L.; Fung-Leung, W.P. Phosphoinositide 3-kinase gamma (PI3Kgamma) inhibitors for the treatment of inflammation and autoimmune disease. Recent Pat. Inflamm. Allergy Drug Discov. 2010, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Yuan, R.; Zhao, X.; Xiang, B.; Gao, S.; Gao, P.; Dai, X.; Feng, M.; Li, Y.; Xie, P.; et al. Transient activation of the PI3K/Akt pathway promotes Newcastle disease virus replication and enhances anti-apoptotic signaling responses. Oncotarget 2017, 8, 23551–23563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbist, K.C.; Klonowski, K.D. Functions of IL-15 in anti-viral immunity: Multiplicity and variety. Cytokine 2012, 59, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Fowell, D.J.; Shinkai, K.; Liao, X.C.; Beebe, A.M.; Coffman, R.L.; Littman, D.R.; Locksley, R.M. Impaired NFATc translocation and failure of Th2 development in Itk-deficient CD4+ T cells. Immunity 1999, 11, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Broussard, C.; Fleischecker, C.; Horai, R.; Chetana, M.; Venegas, A.M.; Sharp, L.L.; Hedrick, S.M.; Fowlkes, B.J.; Schwartzberg, P.L. Altered development of CD8(+) T cell lineages in mice deficient for the Tec kinases Itk and Rlk. Immunity 2006, 25, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atherly, L.O.; Brehm, M.A.; Welsh, R.M.; Berg, L.J. Tec kinases Itk and Rlk are required for CD8(+) T cell responses to virus infection independent of their role in CD4(+) T cell help. J. Immunol. 2006, 176, 3842. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Schorr, W. Role of interleukin-8 in neutrophil signaling. Curr. Opin. Hematol. 2000, 7, 178–182. [Google Scholar] [CrossRef]
- Nagaya, N.; Mori, H.; Murakami, S.; Kangawa, K.; Kitamura, S. Adrenomedullin: Angiogenesis and gene therapy. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 288, R1432–R1437. [Google Scholar] [CrossRef]
- Gros, A.; Syvannarath, V.; Lamrani, L.; Ollivier, V.; Loyau, S.; Goerge, T.; Nieswandt, B.; Jandrot-Perrus, M.; Ho-Tin-Noe, B. Single platelets seal neutrophil-induced vascular breaches via GPVI during immune-complex-mediated inflammation in mice. Blood 2015, 126, 1017–1026. [Google Scholar] [CrossRef]
- Bliss, S.P.; Navratil, A.M.; Xie, J.; Roberson, M.S. GnRH signaling, the gonadotrope and endocrine control of fertility. Front. Neuroendocrinol. 2010, 31, 322–340. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.; Alcantara, S.; Borrell, V.; Del Rio, J.A.; Blasi, J.; Otal, R.; Campos, N.; Boronat, A.; Barbacid, M.; Silos-Santiago, I.; et al. TrkB and TrkC signaling are required for maturation and synaptogenesis of hippocampal connections. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 7336–7350. [Google Scholar] [CrossRef] [Green Version]
- Lussier, A.L.; Weeber, E.J.; Rebeck, G.W. Reelin Proteolysis Affects Signaling Related to Normal Synapse Function and Neurodegeneration. Front. Cell. Neurosci. 2016, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, R.M.; Wurth, M.A.; Aguilar, H.C.; Lee, B.; Moncman, C.L.; McCann, R.O.; Dutch, R.E. Rho GTPase activity modulates paramyxovirus fusion protein-mediated cell-cell fusion. Virology 2006, 350, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Gower, T.L.; Pastey, M.K.; Peeples, M.E.; Collins, P.L.; McCurdy, L.H.; Hart, T.K.; Guth, A.; Johnson, T.R.; Graham, B.S. RhoA signaling is required for respiratory syncytial virus-induced syncytium formation and filamentous virion morphology. J. Virol. 2005, 79, 5326–5336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tybulewicz, V.L.J.; Henderson, R.B. Rho family GTPases and their regulators in lymphocytes. Nat. Rev. Immunol. 2009, 9, 630–644. [Google Scholar] [CrossRef]
- Deist, M.S.; Gallardo, R.A.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel Combined Tissue Transcriptome Analysis after Lentogenic Newcastle Disease Virus Challenge in Inbred Chicken Lines of Differential Resistance. Front. Genet. 2020, 11, 11. [Google Scholar] [CrossRef]
- Schilling, M.A.; Memari, S.; Cattadori, I.M.; Katani, R.; Muhairwa, A.P.; Buza, J.J.; Kapur, V. Innate Immune Genes Associated with Newcastle Disease Virus Load in Chick Embryos From Inbred and Outbred Lines. Front. Microbiol. 2019, 10, 1432. [Google Scholar] [CrossRef] [Green Version]
- Richey, D.J.; Schmittle, S.C. The effect of congenital passive immunity levels on the response of chicks to Newcastle disease vaccination. J. Immunol. 1962, 89, 344–347. [Google Scholar]
- Eidson, C.S.; Kleven, S.H.; Villegas, P. Efficacy of intratracheal administration of Newcastle disease vaccine in day-old chicks. Poult. Sci. 1976, 55, 1252–1267. [Google Scholar] [CrossRef]
- Staszewski, V.; Gasparini, J.; McCoy, K.D.; Tveraa, T.; Boulinier, T. Evidence of an interannual effect of maternal immunization on the immune response of juveniles in a long-lived colonial bird. J. Anim. Ecol. 2007, 76, 1215–1223. [Google Scholar] [CrossRef]
- Hasselquist, D.; Nilsson, J.A. Maternal transfer of antibodies in vertebrates: Trans-generational effects on offspring immunity. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2009, 364, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.H.; Liu, X.Y.; Chen, W.; Chen, L. IFIT5 potentiates anti-viral response through enhancing innate immune signaling pathways. Acta Biochim. Biophys. Sin. 2013, 45, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, Y.M.; Pichmair, A.; Gorna, M.W.; Superti-Furga, G.; Nagar, B. Structural basis for viral 5′-PPP-RNA recognition by human IFIT proteins. Nature 2013, 494, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zheng, Z.; Zhang, Z.; Meng, J.; Liu, Y.; Ke, X.; Hu, Q.; Wang, H. IFIT5 positively regulates NF-kappaB signaling through synergizing the recruitment of IkappaB kinase (IKK) to TGF-beta-activated kinase 1 (TAK1). Cell. Signal. 2015, 27, 2343–2354. [Google Scholar] [CrossRef]
- Rohaim, M.A.; Santhakumar, D.; Naggar, R.F.E.; Iqbal, M.; Hussein, H.A.; Munir, M. Chickens Expressing IFIT5 Ameliorate Clinical Outcome and Pathology of Highly Pathogenic Avian Influenza and Velogenic Newcastle Disease Viruses. Front. Immunol. 2018, 9, 2025. [Google Scholar] [CrossRef]
- Koledova, Z.; Zhang, X.; Streuli, C.; Clarke, R.B.; Klein, O.D.; Werb, Z.; Lu, P. SPRY1 regulates mammary epithelial morphogenesis by modulating EGFR-dependent stromal paracrine signaling and ECM remodeling. Proc. Natl. Acad. Sci. USA 2016, 113, E5731–E5740. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, X.; Cao, W.; Hua, Z.C. Tumor-specifically hypoxia-induced therapy of SPRY1/2 displayed differential therapeutic efficacy for melanoma. Am. J. Cancer Res. 2015, 5, 792–801. [Google Scholar]
- Lee, S.; Bui Nguyen, T.M.; Kovalenko, D.; Adhikari, N.; Grindle, S.; Polster, S.P.; Friesel, R.; Ramakrishnan, S.; Hall, J.L. Sprouty1 inhibits angiogenesis in association with up-regulation of p21 and p27. Mol. Cell. Biochem. 2010, 338, 255–261. [Google Scholar] [CrossRef]
- Yang, X.; Gong, Y.; Tang, Y.; Li, H.; He, Q.; Gower, L.; Liaw, L.; Friesel, R.E. Spry1 and Spry4 differentially regulate human aortic smooth muscle cell phenotype via Akt/FoxO/myocardin signaling. PLoS ONE 2013, 8, e58746. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.H.; Gong, Y.; Friesel, R. Spry1 Is Expressed in Hemangioblasts and Negatively Regulates Primitive Hematopoiesis and Endothelial Cell Function. PLoS ONE 2011, 6, e18374. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.; Waickman, A.; Basson, A.; Kupfer, A.; Licht, J.D.; Horton, M.R.; Powell, J.D. Regulation of CD4(+) and CD8(+) effector responses by Sprouty-1. PLoS ONE 2012, 7, e49801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyanarayana, P.; Dev, A.; Pradeep, A.; Ufkin, M.; Licht, J.D.; Wojchowski, D.M. Spry1 as a novel regulator of erythropoiesis, EPO/EPOR target, and suppressor of JAK2. Blood 2012, 119, 5522–5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basson, M.A.; Watson-Johnson, J.; Shakya, R.; Akbulut, S.; Hyink, D.; Costantini, F.D.; Wilson, P.D.; Mason, I.J.; Licht, J.D. Branching morphogenesis of the ureteric epithelium during kidney development is coordinated by the opposing functions of GDNF and Sprouty1. Dev. Biol. 2006, 299, 466–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehata, H.M.; Khan, S.; Chen, E.; Fields, P.E.; Flavell, R.A.; Sanjabi, S. Lack of Sprouty 1 and 2 enhances survival of effector CD8(+) T cells and yields more protective memory cells. Proc. Natl. Acad. Sci. USA 2018, 115, E8939–E8947. [Google Scholar] [CrossRef] [Green Version]
- Lawir, D.F.; Iwanami, N.; Schorpp, M.; Boehm, T. A missense mutation in zbtb17 blocks the earliest steps of T cell differentiation in zebrafish. Sci. Rep. 2017, 7, 44145. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, J.; Armulik, A.; Senti, K.A.; Edoff, K.; Spalding, K.; Momma, S.; Cassidy, R.; Flanagan, J.G.; Frisen, J. Ephrin-A2 reverse signaling negatively regulates neural progenitor proliferation and neurogenesis. Gene Dev. 2005, 19, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Lennon, F.E.; Mirzapoiazova, T.; Mambetsariev, N.; Mambetsariev, B.; Salgia, R.; Singleton, P.A. Transactivation of the Receptor-tyrosine Kinase Ephrin Receptor A2 Is Required for the Low Molecular Weight Hyaluronan-mediated Angiogenesis That Is implicated in Tumor Progression. J. Biol. Chem. 2014, 289, 24043–24058. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.; Burnett, E.; Kinch, M.; Simon, E.; Wang, B. Activation of EphA2 kinase suppresses integrin function and causes focal-adhesion-kinase dephosphorylation. Nat. Cell Biol. 2000, 2, 62–69. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ensembl ID | Gene Name | 2 Dpi | 6 Dpi | ||
|---|---|---|---|---|---|
| Log2FC | FDR | Log2FC | FDR | ||
| ENSGALG00000008747 | ITGA8 | −0.36 | 0.04896 | −0.41 | 0.04412 |
| ENSGALG00000009100 | FSHR | −0.36 | 0.04813 | −0.45 | 0.04412 |
| ENSGALG00000009626 | THBS1 | −0.67 | 0.01366 | −0.42 | 0.03912 |
| ENSGALG00000027198 | TMEM47 | −0.43 | 0.01800 | −0.52 | 0.00341 |
| ENSGALG00000036566 | ARHGAP20 | −0.41 | 0.00242 | −0.67 | 0.00002 |
| Time | Pathways | Top DEGs Contributing to Prediction | Ratio | z-Score |
|---|---|---|---|---|
| 2 dpi | IL-15 production | EPH(A2/A5/B1), ROR1, MATK, SRC, DDR2, FES, HCK, AATK | 15/121 | −3.357 |
| Signaling by Rho family GTPases | FOS, ARHGEF(11/16/17), ACTA1, ITGA3, JUN, LIMK1, PIK3R6 | 17/244 | −3.153 | |
| GNRH signaling | HBEGF, FOS, EGR1, MAP3K(6/14), CAMK2B, CACNA2D2, SRC | 14/173 | −3.317 | |
| Synaptogenesis signaling pathway | EPH(A2/A5/B1), GRIN3B, CAMK2B, THBS1, SRC, HCK, PIK3R6 | 19/312 | −4.000 | |
| Integrin signaling | ACT(A1/N4), BCAR1, ITGA(3/8), SRC, VCL, MPRIP, RHOB, PIK3R6 | 15/213 | −3.606 | |
| Reelin signaling in neurons | GRIN3B, ARHGEF(11/16), ITGA3, CAMK2B, SRC, HCK, PIK3R6 | 10/129 | −3.162 | |
| GP6 signaling pathway | LAM(B2/A5), COL(5A1/6A3/24A1), PIK3R6 | 9/119 | −3.000 | |
| Tec kinase signaling | FOS, ACTA1, ITGA3, TNFRSF25, SRC, HCK, RHOB, FGR, PIK3R6 | 9/164 | −3.000 | |
| Adrenomedullin signaling pathway | FOS, IL1RN, PLCD3, KCNH2, MATK, NPR2, ITPR3, PIK3R6 | 10/197 | −3.000 | |
| IL-8 signaling | HBEGF, FOS, SRC, JUN, RHOB, LIMK1, PLD1, FLT4, PIK3R6 | 9/200 | −3.000 | |
| 6 dpi | Hematopoiesis from multipotent stem cells | KITLG | 1/12 | - |
| Granulocyte adhesion and diapedesis | HRH1, CCL21 | 2/180 | - | |
| Chondrotin sulfate degradation | HYAL2 | 1/16 | - | |
| Agranulocyte adhesion and diapedesis | HRH1, CCL21 | 2/193 | - | |
| Dermatan sulfate degradation | HYAL2 | 1/17 | - | |
| G-Protein coupled receptor signaling | FSHR, HRH1 | 2/272 | - | |
| Inhibition of angiogenesis by TSP1 | THBS1 | 1/34 | - |
| Time | Disease and Biofunctions | Top DEGs Contributing to Prediction | z-Score | No. |
|---|---|---|---|---|
| 2 dpi | Organismal death | STAR, NR4A3, KLF4, SIK1, HBEGF, CCN1, ADAMTS4, FOS, IL1RN, DUSP1 | 9.770 | 159 |
| Necrosis | NRTN,STAR, NR4A3, KLF4, CCN1, IL1RN, DUSP1, ASTN1, EPHA2, ADAM15 | 2.841 | 162 | |
| Viral Infection | EGR1, CROCC, MAP3K12, FBLIM1, ARTN, DPP4, FAM167B, BCAR1, BMP1 | −5.663 | 90 | |
| Cell viability | NR4A(1/3), HBEGF, CCN(1/2), FOS, DUSP1, CACNB3, WNT5A, EGR1, EPHA2 | −5.568 | 98 | |
| Migration of cells | CCL24, PDCH15, HBEGF, CCN1, FOS, ITGA2B, WNT5A, FOSL2, EGR1, EPHA2 | −5.554 | 142 | |
| Vasculogenesis | CCL24, NR4A1, HBEGF, ATF3, CCN(1/2), FOS, WNT5A, EGR1, EPHA2, ADAM15 | −5.154 | 66 | |
| Microtubule dynamics | ATF3, CCN(1/2), FOS, WNT5A, EGR1, EPHA2, CROCC, METRN, GEM, BCAR1 | −4.553 | 110 | |
| Endocytosis | HBEGF, WNT5A, EGR1, PEAR1, FLNA, CSF1, RASA4, THBS1, SRC, HIP1, TGM2 | −4.147 | 43 | |
| Development of neurons | CCN1, ADAMTS4, FOS, WNT5A, ARTN, METRN, GEM, ITGA3, NR2F1, EGFL7 | −3.640 | 65 | |
| Leukocyte migration | CLL24, HBEGF, CCN(1/2), FOS, ITGA2B, WNT5A, FOSL2, EPHA2, ADAM15 | −2.976 | 69 | |
| Phagocytosis of cells | WNT5A, PEAR1, FLNA, RASA4, THBS1, SRC, SCARF1, TGM2, LRP1, HCK, LIMK1 | −2.398 | 26 | |
| 6 dpi | Proliferation of lymphocytes | HRH1, KITLG, ZEB2, THBS1, CCL21 | −1.938 | 5 |
| Development of neurons | FSHR, KITLG, ZEB2, THBS1, CCL21 | −1.706 | 5 | |
| Recruitment of cells | HRH1, KITLG, THBS1, CCL21 | −1.253 | 4 | |
| Activation of phagocytes | HRH1, KITLG, THBS1, CCL21, HYAL2 | −1.188 | 5 | |
| Microtubule dynamics | KITLG, ZEB2, FSHR, SPATA13, THBS1, ITGA8, CCL21 | −1.083 | 7 |
| Gene Name | Time | LFC | Consistent Response in Other NDV Studies |
|---|---|---|---|
| IFIT5 | 2 dpi | 1.27 | Spleen of Leghorn at 1 [17], 2 [8,17] and 6 dpi [8], and of Fayoumi at 2 dpi [8] |
| CCL19 | 6 dpi | 0.84 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| AICDA | 2 dpi | 0.80 | Trachea of Leghorn at 6 dpi [4] |
| SLBP | 2 dpi | 0.68 | Spleen [8] and trachea [4] of Leghorn at 2 dpi |
| TRIM24 | 2 dpi | 0.52 | Trachea of Leghorn at 6 dpi [4] |
| PLCXD1 | 2 dpi | 0.50 | Harderian gland of Leghorn at 6 dpi [9] |
| SNX10 | 2 dpi | 0.50 | Spleen [8] and trachea [4] of Leghorn at 2 dpi |
| DRAM1 | 2 dpi | 0.49 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| FGL2 | 2 dpi | 0.44 | Trachea of Leghorn at 2 dpi [4] |
| ASNS | 2 dpi | 0.40 | Harderian gland of Leghorn at 6 dpi [9] |
| PARP12 | 2 dpi | 0.40 | Harderian gland at 2 and 6 dpi [9], in spleen at 2 dpi [8] in Leghorn |
| SEPT2 | 2 dpi | 0.38 | Trachea of Fayoumi at 2 and 6 dpi and Leghorn at 6 dpi [4] |
| BFAR | 2 dpi | 0.37 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| OSTM1 | 2 dpi | 0.34 | Trachea of Leghorn at 2 dpi [4] |
| ARHGAP15 | 2 dpi | 0.33 | Trachea of Fayoumi at 2 dpi and Leghorn at 2 and 6 dpi [4] |
| CDC42SE2 | 2 dpi | 0.33 | Trachea of Fayoumi at 2 dpi [4] |
| P2RY8 | 2 dpi | 0.33 | Harderian gland at 6 dpi [9] and trachea at 2 and 6 dpi [4] in Leghorn |
| MYH10 | 2 dpi | −0.37 | Trachea of Leghorn at 6 dpi [4] |
| FSHR | 2 and 6 dpi | −0.40 | Trachea of Leghorn at 6 dpi [4] |
| SEPT11 | 2 dpi | −0.41 | Trachea of Leghorn at 6 dpi [4] |
| OGN | 6 dpi | −0.44 | Trachea of Leghorn at 6 dpi [4] |
| KAZALD1 | 2 dpi | −0.47 | Trachea of Leghorn at 6 dpi [4] |
| SRSF5 | 2 dpi | −0.50 | Trachea of Leghorn at 2 dpi [4] |
| P2RX1 | 2 dpi | −0.51 | Lung of Fayoumi at 10 dpi [10] |
| HPSE2 | 2 dpi | −0.52 | Trachea of Leghorn at 2 dpi [4] |
| JUN | 2 dpi | −0.55 | Leghorn embryo [11] |
| EPHB1 | 2 dpi | −0.57 | Trachea of Fayoumi at 2 dpi [4] |
| LMOD1 | 2 dpi | −0.65 | Lung of Fayoumi at 10 dpi [10] |
| ROR1 | 2 dpi | −0.73 | Lung of Fayoumi at 2 dpi [10] |
| ST3GAL4 | 2 dpi | −0.77 | Harderian gland of Leghorn at 6 dpi under heat stress [12] |
| UROC1 | 2 dpi | −1.01 | Harderian gland of Leghorn at 6 dpi [9] |
| Gene Name | Time | LFC | Trait Associated with Nearby SNPs Identified in Previous Study |
|---|---|---|---|
| EPHA2 | 2 dpi | −1.30 | Antibody titer at 10 dpi in GWAS with the same population [7] |
| FBLIM1 | 2 dpi | −1.09 | Antibody titer at 10 dpi in GWAS with the same population [7] |
| PANX3 | 2 dpi | −0.88 | NDV titer at 6 dpi in GWAS with Hy-Line Brown chickens under heat stress [19] and Tanzanian chicken ecotypes [20] |
| ST3GAL4 | 2 dpi | −0.77 | NDV titer at 6 dpi in GWAS with Hy-Line Brown chickens under heat stress [19] and Tanzanian chicken ecotypes [20] |
| SPRY1 | 2 dpi | −0.69 | Viral load at 6 dpi in GWAS with the same population [7] |
| CPN2 | 2 dpi | −0.65 | Antibody to NDV at 10 dpi in GWAS with Tanzanian chicken ecotypes [20] |
| SRSF5 | 2 dpi | −0.50 | NDV titer at 2 dpi in GWAS with Tanzanian chicken ecotypes [20] |
| ZBTB17 | 2 dpi | −0.33 | Antibody titer at 10 dpi in GWAS with the same population [7] |
| Pathway | z-Score | Activation Predicted by IPA in Other Studies |
|---|---|---|
| Integrin signaling | −3.61 | Lung of Fayoumi at 10 dpi [10] |
| IL-8 signaling | −3.00 | Lung of Fayoumi at 10 dpi [10] and trachea of Fayoumi and Leghorn at 2 dpi [4] |
| Tec kinase signaling | −3.00 | Lung of Fayoumi at 10 dpi [10] and trachea of Fayoumi and Leghorn at 2 dpi [4] |
| GP6 signaling pathway | −3.00 | Harderian gland of Leghorn at 6 dpi under heat stress [12] |
| B cell receptor signaling | −2.83 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| IL-6 signaling | −2.83 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| Production of nitric oxide and reactive oxygen species in macrophage | −2.83 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| P2γ purigenic receptor signaling pathway | −2.45 | Lung of Fayoumi at 10 dpi [10] |
| Fcγ receptor-mediated phagocytosis in macrophages and monocytes | −2.45 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| Thrombin signaling | −2.33 | Lung of Fayoumi at 10 dpi [10] |
| Ephrin receptor signaling | −2.24 | Lung of Fayoumi at 10 dpi [10] |
| Relaxin signaling | −2.24 | Lung of Fayoumi at 10 dpi [10] |
| CD40 signaling | −2.24 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| Leukocyte extravasation signaling | −2.12 | Trachea of Fayoumi and Leghorn at 2, 6 and 10 dpi [4] |
| TNFR1 signaling | −2.00 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
| TNFR2 signaling | −2.00 | Trachea of Fayoumi and Leghorn at 2 dpi [4] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Kaiser, M.G.; Gallardo, R.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Transcriptome Analysis Reveals Inhibitory Effects of Lentogenic Newcastle Disease Virus on Cell Survival and Immune Function in Spleen of Commercial Layer Chicks. Genes 2020, 11, 1003. https://doi.org/10.3390/genes11091003
Zhang J, Kaiser MG, Gallardo RA, Kelly TR, Dekkers JCM, Zhou H, Lamont SJ. Transcriptome Analysis Reveals Inhibitory Effects of Lentogenic Newcastle Disease Virus on Cell Survival and Immune Function in Spleen of Commercial Layer Chicks. Genes. 2020; 11(9):1003. https://doi.org/10.3390/genes11091003
Chicago/Turabian StyleZhang, Jibin, Michael G. Kaiser, Rodrigo A. Gallardo, Terra R. Kelly, Jack C. M. Dekkers, Huaijun Zhou, and Susan J. Lamont. 2020. "Transcriptome Analysis Reveals Inhibitory Effects of Lentogenic Newcastle Disease Virus on Cell Survival and Immune Function in Spleen of Commercial Layer Chicks" Genes 11, no. 9: 1003. https://doi.org/10.3390/genes11091003