Telomere Dynamics in the Diploid and Triploid Rainbow Trout (Oncorhynchus mykiss) Assessed by Q-FISH Analysis

Abstract
:1. Introduction
2. Materials and Method
2.1. Fish Origin
2.2. Egg Incubation and Fish Rearing
2.3. Preparation of Interphase Spreads
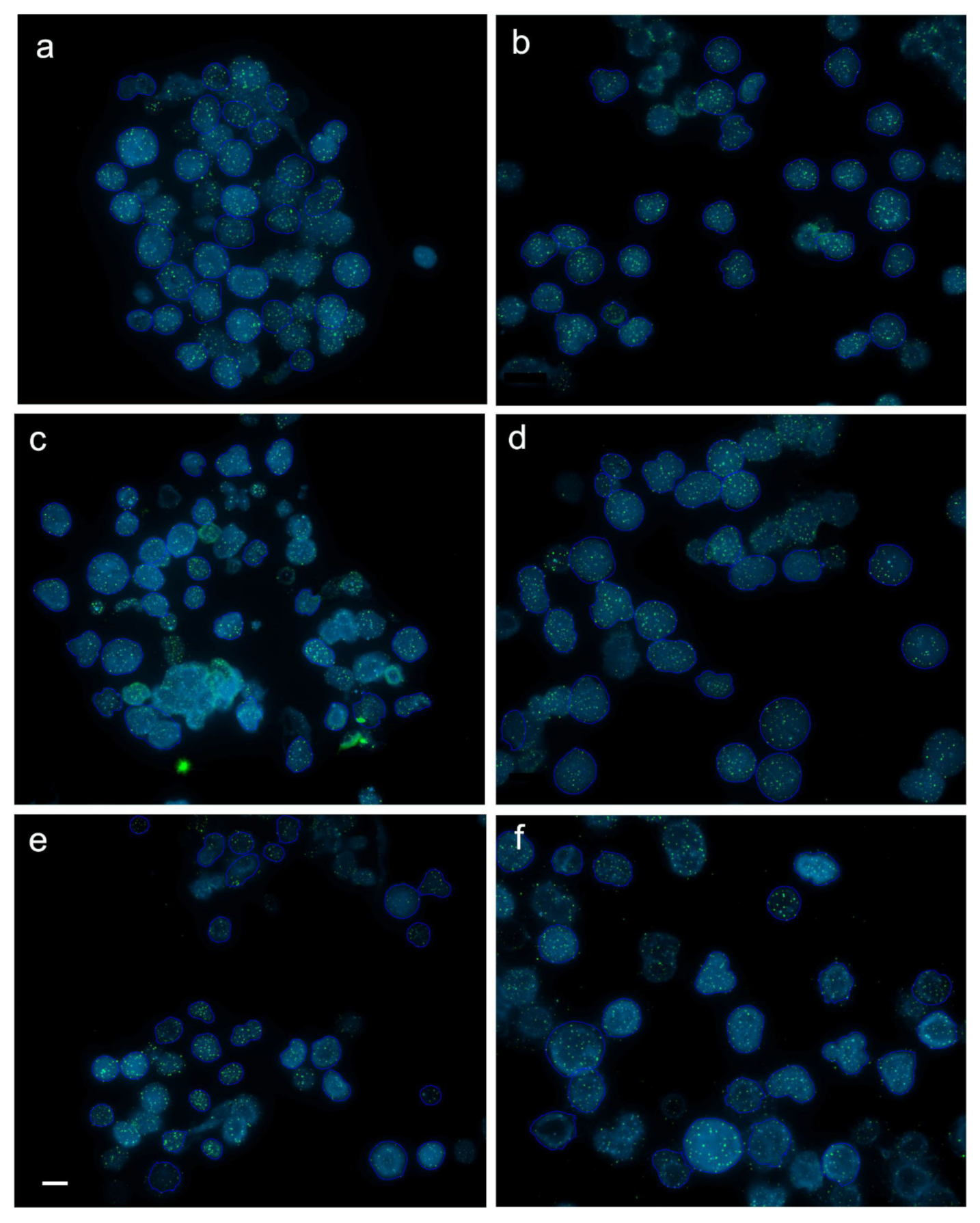
2.4. Interphase Quantitative Fluorescence In Situ Hybridization (Q-FISH)
2.5. Statistical Analysis
3. Results
3.1. Body Weight and Length and Gonadal Development
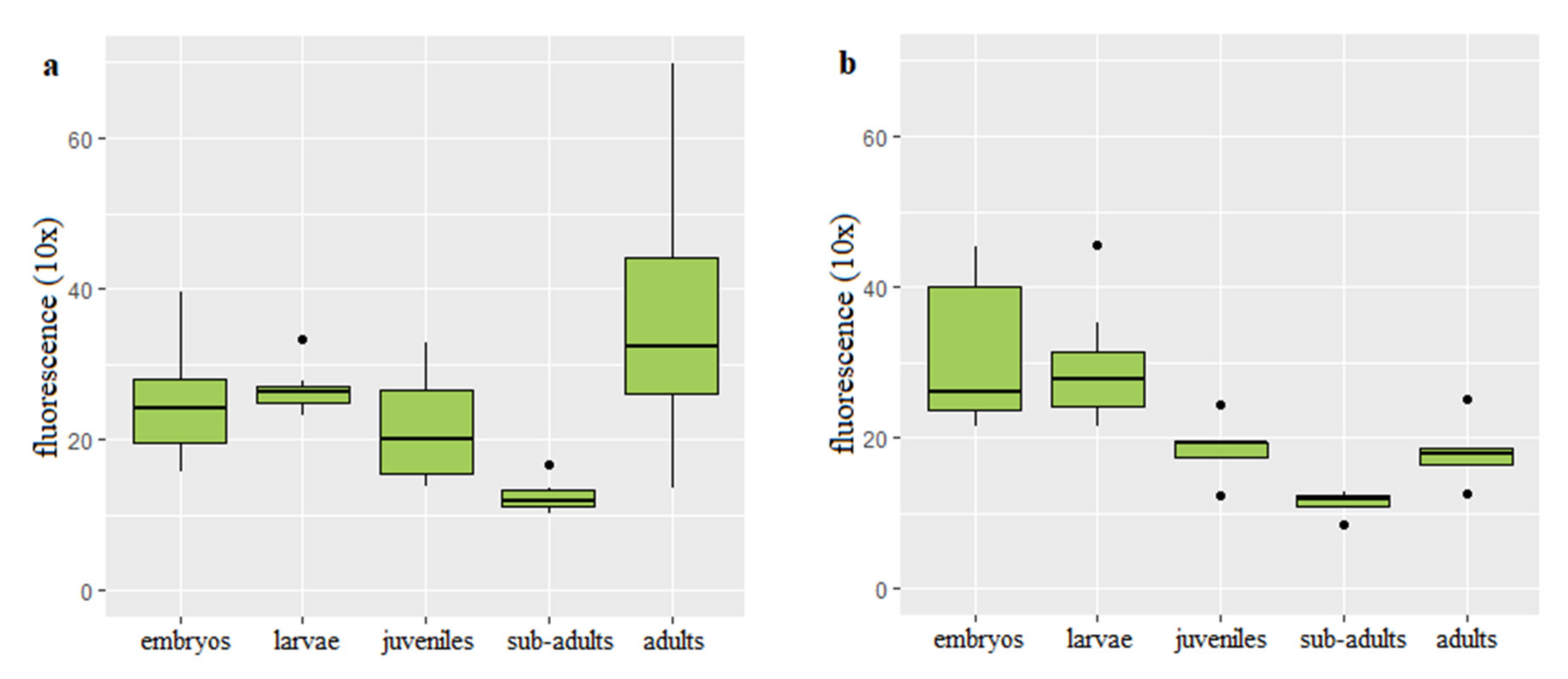
3.2. Dynamics of Telomere Length in Rainbow Trout
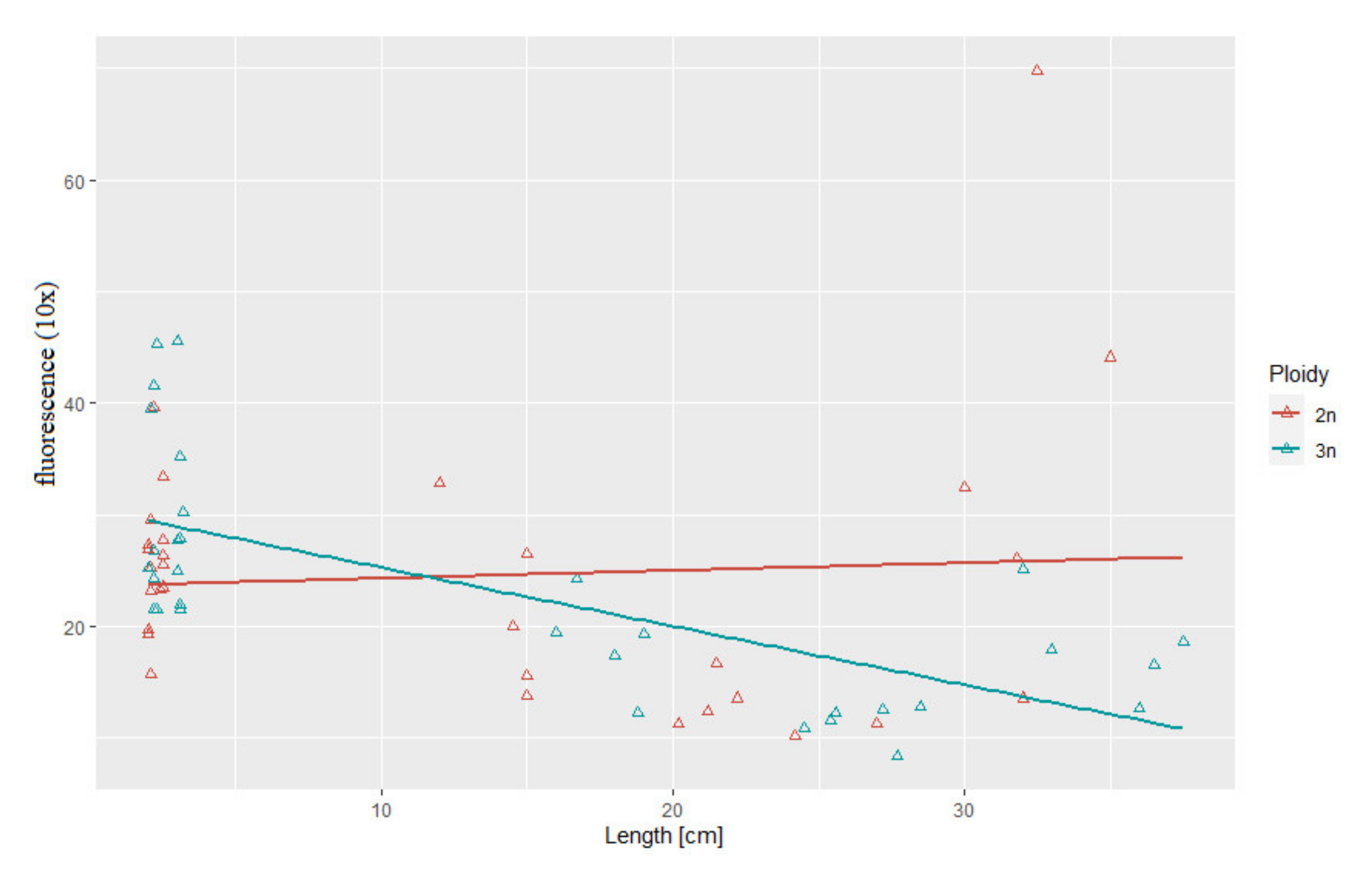
3.3. Correlation between Body Weight and Length and Telomere Length-Related Fluorescence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyne, J.; Ratliff, R.L.; Moyzis, R.K. Conservation of the human telomere sequence (TTAGGG) n among vertebrates. Proc. Natl. Acad. Sci. USA 1989, 86, 7049–7053. [Google Scholar] [CrossRef] [Green Version]
- Bolzán, A.D.; Bianchi, M.S. Telomeres, interstitial telomeric repeat sequences, and chromosomal aberrations. Mutat. Res. Rev. Mutat. Res. 2006, 612, 189–214. [Google Scholar] [CrossRef]
- De Lange, T. Protection of mammalian telomeres. Oncogene 2002, 21, 532–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olovnikov, A.M. A theory of marginotomy: The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J. Theor. Biol. 1973, 41, 181–190. [Google Scholar] [CrossRef]
- Von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemato, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef]
- Klapper, W.; Heidorn, K.; Kühne, K.; Parwaresch, R.; Krupp, G. Telomerase activity in ‘immortal’fish 1. FEBS Lett. 1998, 434, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Elmore, L.W.; Norris, M.W.; Sircar, S.; Bright, A.T.; McChesney, P.A.; Winn, R.N.; Holt, S.E. Upregulation of telomerase function during tissue regeneration. Exp. Biol. Med. 2008, 233, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.C.; Glass, T.J.; Tolar, J.; Blazar, B.R. Expression of telomerase and telomere length are unaffected by either age or limb regeneration in Danio rerio. PLoS ONE 2009, 4, e7688. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.M.; Shay, J.W.; Wright, W.E. Telomere biology in Metazoa. FEBS Lett. 2010, 584, 3741–3751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, M.; Wapstra, E.; Friesen, C. Ectothermic telomeres: It’s time they came in from the cold. In Philosophical Transactions of the Royal Society B: Biological Sciences 373; The Royal Society: Lodon, UK, 2018. [Google Scholar]
- Scott, N.M.; Haussmann, M.F.; Elsey, R.M.; Trosclair, P.L.; Vleck, C.M. Telomere length shortens with body length in Alligator mississippiensis. Southeast Nat. 2006, 5, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Bronikowski, A.M. The evolution of aging phenotypes in snakes: A review and synthesis with new data. Age 2008, 30, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plot, V.; Criscuolo, F.; Zahn, S.; Georges, J.J.-Y. Telomeres, Age and Reproduction in a long-lived reptile. PLoS ONE 2012, 7, e40855. [Google Scholar] [CrossRef]
- Hatakeyama, H.; Nakamura, K.I.; Izumiyama-Shimomura, N.; Ishii, A.; Tsuchida, S.; Takubo, K.; Ishikawa, N. The teleost Oryzias latipes shows telomere shortening with age despite considerable telomerase activity throughout life. Mech. Ageing Dev. 2008, 129, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, N.; Reichwald, K.; Lechel, A.; Graf, M.; Kirschner, J.; Dorn, A.; Terzbasi, E.; Wellner, J.; Platzer, M.; Rudolph, K.; et al. Telomeres shorten while Tert expression increases during ageing of the short-lived fish Nothobranchius furzeri. Mech. Ageing Dev. 2009, 130, 290–296. [Google Scholar] [CrossRef]
- Horn, T.; Gemmell, N.J.; Robertson, B.C.; Bridges, C.R. Telomere length change in European sea bass (Dicentrarchus labrax). Aust J. Zool. 2008, 56, 207–210. [Google Scholar] [CrossRef]
- Gao, J.; Munch, S.B. Does reproductive investment decrease telomere length in Menidia menidia? PLoS ONE 2015, 10, e0125674. [Google Scholar] [CrossRef]
- Anchelin, M.; Murcia, L.; Alcaraz-Pérez, F.; García-Navarro, E.M.; Cayuela, M.L. Behaviour of telomere and telomerase during aging and regeneration in zebrafish. PLoS ONE 2011, 6, e16955. [Google Scholar] [CrossRef]
- Monaghan, P.; Ozanne, S.E. Somatic growth and telomeric dynamics in vertebrates: Relationships, mechanisms and consequences. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160446. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.L.B.; Richardson, D.S. Sex differences in telomeres and lifespan. Aging Cell 2011, 10, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Viña, J.; Borrás, C.; Gambini, J.; Sastre, J.; Pallardó, F.V. Why females live longer than males? Importance of the upregulation of longevity-associated genes by oestrogenic compounds. FEBS Lett. 2005, 12, 2541–2545. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, W.R.; Friesen, C.R.; Sihlbom, C.; Bergström, J.; Berger, E.; Wilson, M.R.; Olsson, M. Vitellogenin offsets oxidative costs of reproduction in female painted dragon lizards. J. Exp. Biol. 2020, 223, jeb221630. [Google Scholar] [CrossRef]
- Piferrer, F.; Beaumont, A.; Falguière, J.C.; Flajšhans, M.; Haffray, P.; Colombo, L. Polyploid fish and shellfish: Production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture 2009, 293, 125–156. [Google Scholar] [CrossRef] [Green Version]
- Tivary, B.K.; Kirubagaran, R.; Ray, A.K. The biology of triploid fish. Rev. Fish Biol. Fish. 2004, 14, 391–402. [Google Scholar]
- Cormier, S.M.; Neiheisel, T.W.; Williams, D.E.; Tiersch, T.R. Natural occurrence of triploidy in a wild brown bullhead. Trans. Am. Fish. Soc. 1993, 122, 390–392. [Google Scholar] [CrossRef]
- Fernandes-Matioli, F.M.C.; Almeida-Toled, L.F.; Toledo-Filho, S.A. Natural triploidy in the Neotropical species Gymnotus carapo (Pisces: Gymnotiformes). Caryologia 1998, 51, 319–322. [Google Scholar] [CrossRef]
- Ocalewicz, K.; Dobosz, S. Karyotype variation in the albino rainbow trout (Oncorhynchus mykiss (Walbaum)). Genome 2009, 52, 347–352. [Google Scholar] [CrossRef]
- Pandian, T.J.; Koteeswaran, R. Ploidy induction and sex control in fish. Hydrobiologia 1998, 384, 167–243. [Google Scholar] [CrossRef]
- Poon, S.S.; Lansdorp, P.M. Quantitative fluorescence in situ hybridization (Q-FISH). Curr. Protoc. Cell Biol. 2001, 12, 18.4.1–18.4.21. [Google Scholar] [CrossRef]
- Montpetit, A.J.; Alhareeri, A.A.; Montpetit, M.; Starkweather, A.R.; Elmore, L.W.; Filler, K.; Mohanraj, L.; Burton, C.W.; Menzies, V.S.; Lyon, D.E.; et al. Telomere length: A review of methods for measurement. Nurs. Res. 2014, 63, 289. [Google Scholar] [CrossRef] [Green Version]
- Polonis, M.; Fujimoto, T.; Dobosz, S.; Zalewski, T.; Ocalewicz, K. Genome incompatibility between rainbow trout (Oncorhynchus mykiss) and sea trout (Salmo trutta) and induction of the interspecies gynogenesis. J. Appl. Genet. 2018, 59, 91–97. [Google Scholar] [CrossRef]
- Ocalewicz, K.; Furgala-Slezniow, G.; Szmyt, M.; Lisboa, R.; Kuciński, M.; Lejk, A.M.; Jankun, M. Pericentromeric location of the telomeric DNA sequences on the European grayling chromosomem. Genetica 2013, 141, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocalewicz, K.; Woznicki, P.; Jankun, M. Mapping of rRNA genes and telomeric sequences in Danube salmon (Hucho hucho) chromosomes using primed in situ labeling technique (PRINS). Genetica 2008, 134, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Ocalewicz, K.; Sliwinska, A.; Jankun, M. Autosomal localization of Interstitial Telomeric Site (ITS) in brook trout, Salvelinus fontinalis (Pisces, Salmonidae). Cytogenet. Genome Res. 2004, 105, 79–82. [Google Scholar] [CrossRef]
- Jankun, M.; Woznicki, P.; Ocalewicz, K.; Furgala-Selezniow, G. Chromosomal evolution in the three species of Holarctic fish of the genus Coregonus (Salmoniformes). Adv. Limnol. 2007, 60, 25–37. [Google Scholar]
- Pauliny, A.; Devlin, R.H.; Johnsson, J.I.; Blomqvist, D. Rapid growth accelerates telomere attrition in a transgenic fish. BMC Evol. Biol. 2015, 15, 159. [Google Scholar] [CrossRef] [Green Version]
- McLennan, D.; Armstrong, J.D.; Stewart, D.C.; Mckelvey, S.; Boner, W.; Monaghan, P.; Metcalfe, N.B. Shorter juvenile telomere length is associated with higher survival to spawning in migratory Atlantic salmon. Funct. Ecol. 2017, 31, 2070–2079. [Google Scholar] [CrossRef] [Green Version]
- McLennan, D.; Armstrong, J.D.; Stewart, D.C.; Mckelvey, S.; Boner, W.; Monaghan, P.; Metcalfe, N.B. Telomere elongation during early development is independent of environmental temperatures in Atlantic salmon. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Fauconneau, B.; Kaushik, S.J.; Blanc, J.M. Uptake and metabolization of dissolved compounds in rainbow trout (Salmo gairdneri R.) fry. Comp. Biochem. Physiol. 1989, 93, 839–843. [Google Scholar] [CrossRef] [Green Version]
- Oliva-Teles, A.; Kaushik, S.J. Nitrogen and energy metabolism during the early ontogeny of diploid and triploid rainbow trout (Salmo gairdneri R.). Comp. Biochem. Physiol. 1987, 87, 157–160. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Kaushik, S.J. Effect of temperature on utilization of endogenous energy reserves during embryonic development of diploid and triploid rainbow trout (Salmo gairdneri R.). Aquaculture 1990, 84, 373–382. [Google Scholar] [CrossRef]
- Shrimpton, J.M.; Heath, J.W.; Devlin, R.H.; Heath, D.D. Effect of triploidy on growth and ionoregulatory performance in ocean-type Chinook salmon: A quantitative genetics approach. Aquaculture 2012, 362, 248–254. [Google Scholar] [CrossRef]
- Leggatt, R.A.; Scheer, K.W.; Afonso, L.O.; Iwama, G.K. Triploid and diploid rainbow trout do not differ in their stress response to transportation. N. Am. J. Aquac. 2006, 68, 1–8. [Google Scholar] [CrossRef]
- Chalmers, L.; Vera, L.M.; Taylor, J.F.; Adams, A.; Migaud, H. Comparative ploidy response to experimental hydrogen peroxide exposure in Atlantic salmon (Salmo salar). Fish Shelfish Immun. 2018, 81, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Risques, R.A.; Promislow, D.E. All’s well that ends well: Why large species have short telomeres. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160448. [Google Scholar] [CrossRef] [Green Version]
- Pauliny, A.; Larsson, K.; Blomqvist, D. Telomere dynamics in a long-lived bird, the barnacle goose. BMC Evol. Biol. 2012, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rollings, N.; Uhrig, E.J.; Krohmer, R.W.; Waye, H.L.; Mason, R.T.; Olsson, M.; Whittington, C.M.; Friesen, C.R. Age-related sex differences in body condition and telomere dynamics of red-sided garter snakes. Proc. R. Soc. B 2017, 284, 61. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, H.; Yamazaki, H.; Nakamura, K.I.; Izumiyama-Shimomura, N.; Aida, J.; Suzuki, H.; Tsuchida, S.; Matsuura, M.; Takubo, K.; Ishikawa, N. Telomere attrition and restoration in the normal teleost Oryzias latipes are linked to growth rate and telomerase activity at each life stage. Aging 2016, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Biessmann, H.; Mason, J.M. Telomere maintenance without telomerase. Chromosoma 1997, 106, 63–69. [Google Scholar] [CrossRef]
- Henson, J.D.; Neumann, A.A.; Yeager, T.R.; Reddel, R.R. Alternative lengthening of telomeres in mammalian cells. Oncogene 2002, 21, 598–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of Development | Ploidy | Length (cm) | Weight (g) | ||
|---|---|---|---|---|---|
| Mean | ±SD | Mean | ±SD | ||
| Embryos | 2n | 2.08 * | 0.07 | 0.12 | 0.009 |
| 3n | 2.19 * | 0.10 | 0.12 | 0.010 | |
| Larvae | 2n | 2.43 * | 3.14 | 0.12 * | 0.017 |
| 3n | 3.08 * | 0.07 | 0.21 * | 0.021 | |
| Juveniles (one year old) | 2n | 14.30 * | 1.30 | 26.12 * | 5.01 |
| 3n | 17.70 * | 1.31 | 60.28 * | 11.80 | |
| Subadults (two years old) | 2n | 22.72 * | 2.49 | 126.83 * | 47.10 |
| 3n | 26.48 * | 1.55 | 204.00 * | 46.08 | |
| Adults (three years old) | 2n | 32.26 | 1.80 | 425.20 | 109.20 |
| 3n | 35.00 | 2.37 | 442.40 | 151.76 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panasiak, L.; Dobosz, S.; Ocalewicz, K. Telomere Dynamics in the Diploid and Triploid Rainbow Trout (Oncorhynchus mykiss) Assessed by Q-FISH Analysis. Genes 2020, 11, 786. https://doi.org/10.3390/genes11070786
Panasiak L, Dobosz S, Ocalewicz K. Telomere Dynamics in the Diploid and Triploid Rainbow Trout (Oncorhynchus mykiss) Assessed by Q-FISH Analysis. Genes. 2020; 11(7):786. https://doi.org/10.3390/genes11070786
Chicago/Turabian StylePanasiak, Ligia, Stefan Dobosz, and Konrad Ocalewicz. 2020. "Telomere Dynamics in the Diploid and Triploid Rainbow Trout (Oncorhynchus mykiss) Assessed by Q-FISH Analysis" Genes 11, no. 7: 786. https://doi.org/10.3390/genes11070786