Complex Structure of Lasiopodomys mandarinus vinogradovi Sex Chromosomes, Sex Determination, and Intraspecific Autosomal Polymorphism

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Specimens Sampled
2.3. Chromosome Preparation and Chromosome Staining
2.4. Crossbreeding Experiments
2.5. Female Reproductive Success in Relation to Karyomorphs
2.6. Microdissection and Probe Amplification
2.7. Fluorescence in Situ Hybridization (FISH)
2.8. Sequencing
2.9. Bioinformatic Analysis
3. Results
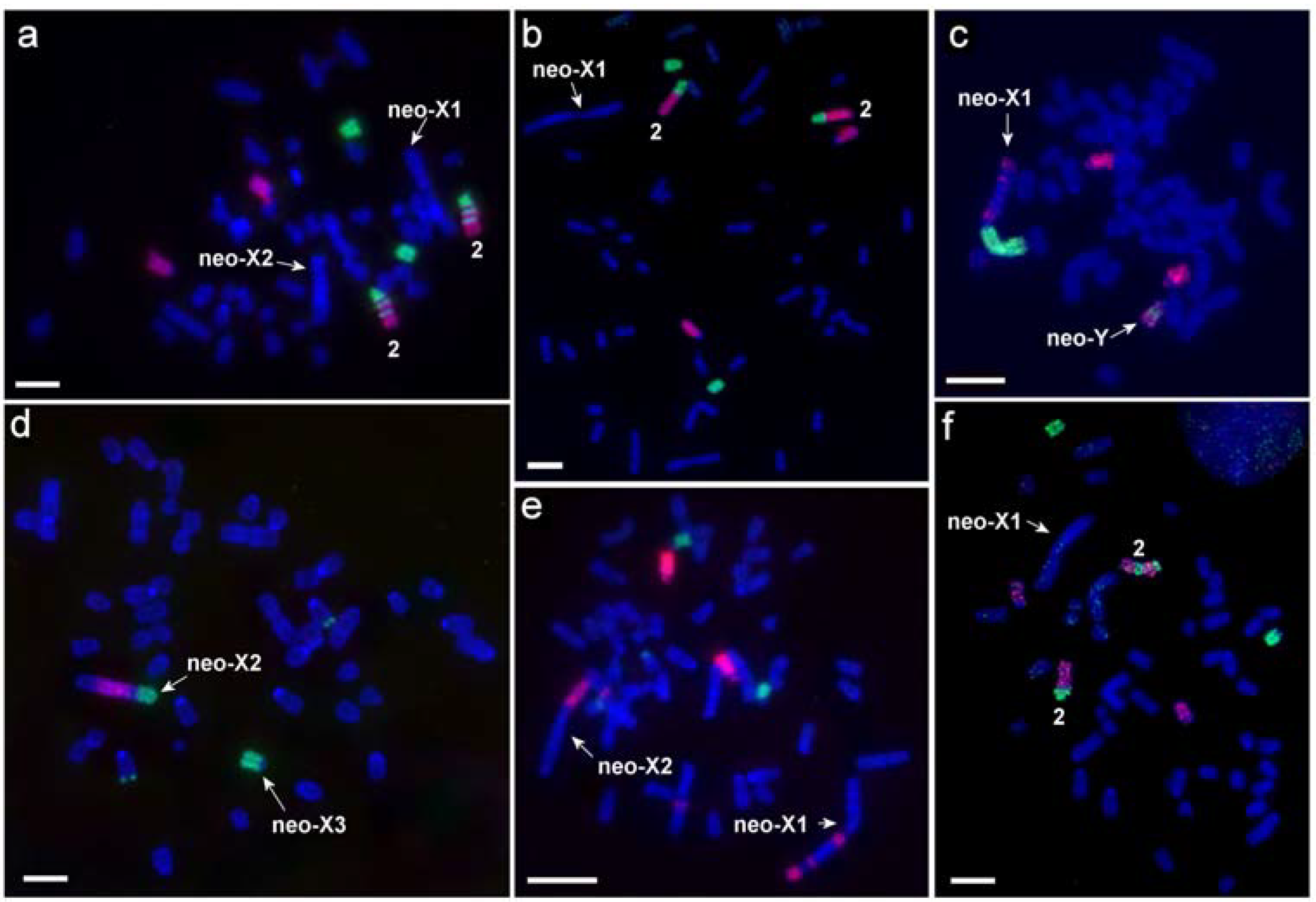
3.1. Sex Chromosome Combinations Revealed by Extensive Karyotyping
3.2. Hybridization Experiment
3.3. Female Reproductive Success Related to Their Karyomorphs
3.4. Comparative Molecular Cytogenetic Investigation of Different L. m. vinogradovi Karyomorphs
3.5. Sequencing and Bioinformatic Analysis
4. Discussion
4.1. How Is Such an Unusual System Maintained in Populations of L. m. vinogradovi?
4.2. Complex Systems of Sex Chromosomes in L. m. vinogradovi and Their Origin
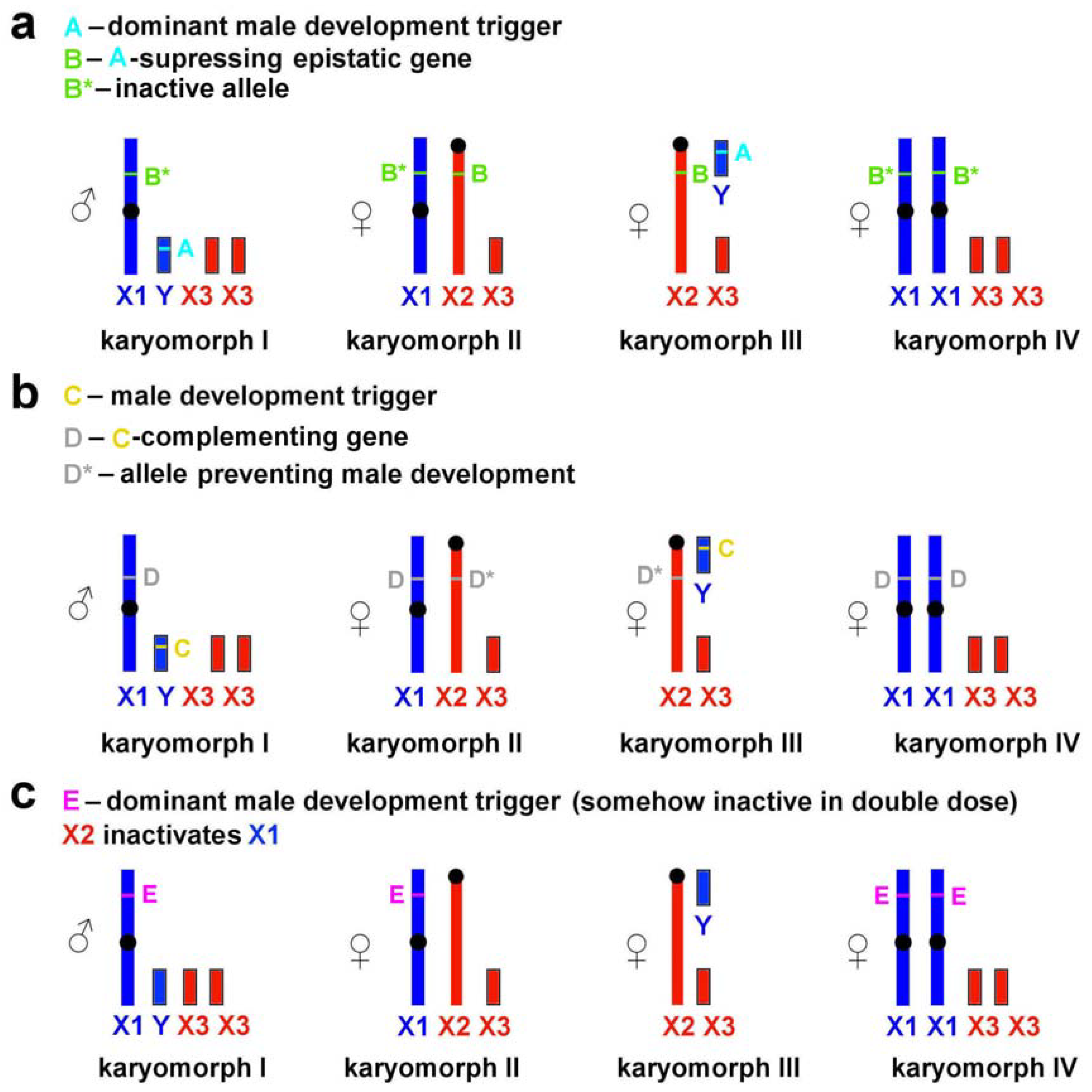
4.3. Possible Mechanisms of Sex Determination in L. m. vinogradovi
- (1)
- The neo-X2 chromosome contains an epistatic locus (B) suppressing the dominant male development trigger (A) located on the neo-Y (Figure 4a).
- (2)
- The neo-X1 chromosome contains a locus (D) complementing the male development trigger (C), whereas this locus is absent from or is inactive on the homologous neo-X2 (Figure 4b). In this case, the sex-determination system is similar to that described for Myopus schisticolor [41] where a deletion differentiates two types of X chromosomes [43]; the same has been suggested for Dicrostonyx torquatus [42]. Zhu et al. [8] proposed a possible role of deletions in the formation of sex chromosomes in L. m. faeceus.
- (3)
- Taking into account the uncertainty of the location of male development gene(s), it cannot be ruled out that they are associated not with neo-Y but with neo-X1 chromosome only (Figure 4c). According to this scenario, the neo-X2 chromosome either is capable of inactivating neo-X1 or carries a trigger-suppressing gene resulting in female phenotype of neo-X1/neo-X2 carriers Although nonrandom inactivation of the X chromosome has not been described for the mandarin voles, it has been identified in experiments on interspecific crosses of several arvicoline species [45]. This scenario, however, appears to be the least plausible as it requires the dominant male development trigger to be somehow inactive in double doses to produce neo-X1/neo-X1 females.
4.4. Chromosomal Differences of L. mandarinus from Different Populations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bakloushinskaya, I.Y. Evolution of sex determination in mammals. Biol. Bull. 2009, 36, 167–174. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Volobouev, V.T. Non-sciuromorph rodent karyotypes in evolution. Cytogenet. Genome Res. 2012, 137, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Kovalskaya, Y.M.; Orlov, V.N. Unusual sex chromosomes and intrapopulation chromosomal polymorphism in the Chinese vole. Tsitologiia 1974, 16, 497. [Google Scholar]
- Zhu, B.C.; Wang, H.Y.; Qu, A. Studies on fertility of XO females in the Microtus mandarinus. Acta Zool. Sin. 1998, 44, 209–212. [Google Scholar]
- Zhu, B.; Gao, H.; Wang, H.; Gao, J.; Zhang, Y.; Dong, Y.; Hou, J.; Nan, X. The origin of the genetical diversity of Microtus mandarinus chromosomes. Hereditas 2003, 139, 90–95. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, J.; Xu, Y.; Zhang, Y.; Wang, T. Cytogenetic studies of brown field-mouse. Yi Chuan Xue Bao 1993, 20, 135–140. [Google Scholar]
- Wang, J.X.; Zhao, X.F.; Deng, Y.; Qi, H.Y.; Wang, Z.J. Chromosomal polymorphism of mandarin vole, Microtus mandarinus (Rodentia). Hereditas 2003, 138, 47–53. [Google Scholar] [CrossRef]
- Zhu, B.; Dong, Y.; Gao, J.; Li, P.; Pang, Y.; Liu, H.; Chen, H. Numerical and structural variations of the X chromosomes and no. 2 autosomes in mandarin vole, Microtus mandarinus (Rodentia). Hereditas 2006, 143, 130–137. [Google Scholar] [CrossRef]
- Kovalskaya, Y.M.; Smorkatcheva, A.V. Overview of geographical variability of karyotype of Chinese vole Lasiopodomys mandarinus Milne-Edwards 1871 (Rodentia, Arvicolinae)–Another group of close species? In Proceedings of the Teriofauna of Russia and adjacent territories; Nauka: Moscow, 2011; p. 227. [Google Scholar]
- Gladkikh, O.L.; Romanenko, S.A.; Lemskaya, N.A.; Serdyukova, N.A.; O’Brien, P.C.; Kovalskaya, J.M.; Smorkatcheva, A.V.; Golenishchev, F.N.; Perelman, P.L.; Trifonov, V.A. Rapid karyotype evolution in Lasiopodomys involved at least two autosome–sex chromosome translocations. PLoS ONE 2016, 11, e0167653. [Google Scholar] [CrossRef]
- Liu, H.; Yan, N.; Zhu, B. Two new karyotypes and bandings in Microtus mandarinus faeceus (Rodentia): Two new karyotypes and bandings in M. m. faeceus. Hereditas 2010, 147, 123–126. [Google Scholar] [CrossRef]
- Zhang, Z.; Gu, W.; Zhu, B.; Wang, T. Study on synaptonemal complexes in spermatocytes of Microtus mandarinus. J. Shaanxi Norm. Univ. Nat. Sci. Ed. 1999, 27, 93–96. [Google Scholar]
- Yan, N.; Zhu, B.C.; Wang, Y.F. Recent advances on sex determining mechanisms of Microtus mandarinus. Yi Chuan Hered. 2009, 31, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Borodin, P.M.; Basheva, E.A.; Torgasheva, A.A.; Dashkevich, O.A.; Golenishchev, F.N.; Kartavtseva, I.V.; Mekada, K.; Dumont, B.L. Multiple independent evolutionary losses of XY pairing at meiosis in the grey voles. Chromosome Res. 2012, 20, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ming, Q.; Zhu, B. Exclusion of Sall 4 as the sex-determining gene in the Mandarin vole Microtus mandarinus mandarinus. Hereditas 2011, 148, 93–97. [Google Scholar] [CrossRef]
- Chen, Y.; Dong, Y.; Xiang, X.; Zhang, X.; Zhu, B. Sex determination of Microtus mandarinus mandarinus is independent of Sry gene. Mamm. Genome 2008, 19, 61–68. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Radjabli, S.I. Chromosomes of Agricultural and Laboratory Mammals; An atlas; Nauka: Novosibirsk, 1989; p. 128. [Google Scholar]
- Stanyon, R.; Galleni, L. A rapid fibroblast culture technique for high resolution karyotypes. Ital. J. Zool. 1991, 58, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, S.A.; Perelman, P.L.; Trifonov, V.A.; Serdyukova, N.A.; Li, T.; Fu, B.; O’Brien, P.C.; Ng, B.L.; Nie, W.; Liehr, T. A first generation comparative chromosome map between Guinea pig (Cavia porcellus) and humans. PLoS ONE 2015, 10, e0127937. [Google Scholar] [CrossRef]
- Seabright, M. A rapid banding technique for human chromosomes. Lancet 1971, 298, 971–972. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Smorkatcheva, A.V. Parental care in the captive mandarin vole, Lasiopodomys mandarinus. Can. J. Zool. 2003, 81, 1339–1345. [Google Scholar] [CrossRef]
- Yang, F.; Trifonov, V.A.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of paint probes from flow-sorted and microdissected chromosomes. In Fluorescence in Situ Hybridization (FISH)-Application Guide; Springer: Berlin, Heidelberg, 2017; pp. 67–79. [Google Scholar]
- Telenius, H.; Ponder, B.A.; Tunnacliffe, A.; Pelmear, A.H.; Carter, N.P.; Ferguson-Smith, M.A.; Behmel, A.; Nordenskjöld, M.; Pfragner, R. Cytogenetic analysis by chromosome painting using DOP-PCR amplified flow-sorted chromosomes. Genes. Chromosomes Cancer 1992, 4, 257–263. [Google Scholar] [CrossRef]
- Yang, F.; Carter, N.P.; Shi, L.; Ferguson-Smith, M.A. A comparative study of karyotypes of muntjacs by chromosome painting. Chromosoma 1995, 103, 642–652. [Google Scholar] [CrossRef]
- Sitnikova, N.A.; Romanenko, S.A.; O’Brien, P.C.M.; Perelman, P.L.; Fu, B.; Rubtsova, N.V.; Serdukova, N.A.; Golenishchev, F.N.; Trifonov, V.A.; Ferguson-Smith, M.A.; et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). I. The genome homology of tundra vole, field vole, mouse and golden hamster revealed by comparative chromosome painting. Chromosome Res. 2007, 15, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S.A.; Lemskaya, N.A.; Trifonov, V.A.; Serdukova, N.; O’Brien, P.C.M.; Bulatova, N.S.; Golenishchev, F.N.; Ferguson-Smith, M.A.; Yang, F.; Graphodatsky, A.S. Genome-wide comparative chromosome maps of Arvicola amphibius, Dicrostonyx torquatus, and Myodes rutilus. Chromosome Res. 2016, 24, 145–159. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maden, B.E.; Dent, C.L.; Farrell, T.E.; Garde, J.; McCallum, F.S.; Wakeman, J.A. Clones of human ribosomal DNA containing the complete 18 S-rRNA and 28 S-rRNA genes. Characterization, a detailed map of the human ribosomal transcription unit and diversity among clones. Biochem. J. 1987, 246, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; O’Brien, P.C.M.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A Complete comparative chromosome map for the dog, red fox, and human and its integration with canine genetic maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Graphodatsky, A.S.; Sablina, O.V.; Meyer, M.N.; Malikov, V.G.; Isakova, E.A.; Trifonov, V.A.; Polyakov, A.V.; Lushnikova, T.P.; Vorobieva, N.V.; Serdyukova, N.A.; et al. Comparative cytogenetics of hamsters of the genus Calomyscus. Cytogenet. Genome Res. 2000, 88, 296–304. [Google Scholar] [CrossRef]
- Makunin, A.I.; Kichigin, I.G.; Larkin, D.M.; O’Brien, P.C.; Ferguson-Smith, M.A.; Yang, F.; Proskuryakova, A.A.; Vorobieva, N.V.; Chernyaeva, E.N.; O’Brien, S.J. Contrasting origin of B chromosomes in two cervids (Siberian roe deer and grey brocket deer) unravelled by chromosome-specific DNA sequencing. BMC Genomics 2016, 17, 618. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–251760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olshen, A.B.; Venkatraman, E.S.; Lucito, R.; Wigler, M. Circular binary segmentation for the analysis of array-based DNA copy number data. Biostatistics 2004, 5, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Bulmer, M.G. The evolution of XY females in mammals. Heredity 1981, 47, 347–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredga, K. Aberrant sex chromosome mechanisms in mammals. Diffentiation 1983, 23, S23–S30. [Google Scholar]
- Bianchi, N.O. Akodon sex reversed females: The never ending story. Cytogenet. Genome Res. 2002, 96, 60–65. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection; Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Zhao, S.; Shetty, J.; Hou, L.; Delcher, A.; Zhu, B.; Osoegawa, K.; de Jong, P.; Nierman, W.C.; Strausberg, R.L.; Fraser, C.M. Human, mouse, and rat genome large-scale rearrangements: Stability versus speciation. Genome Res. 2004, 14, 1851–1860. [Google Scholar] [CrossRef] [Green Version]
- Fredga, K.; Groop, A.; Winking, H.; Frank, F. A hypothesis explaining the exceptional sex ratio in the wood lemming (Myopus schisticolor). Hereditas 1977, 85, 101–104. [Google Scholar] [CrossRef]
- Gileva, E.A.; Chebotar, N.A. Fertile XO males and females in the varying lemming, Dicrostonyx torquatus Pall.(1779). Heredity 1979, 42, 67. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-S.; Eriksson, L.; Fredga, K. XY sex reversal in the wood lemming is associated with deletion of Xp21-23 as revealed by chromosome microdissection and fluorescence in situ hybridization. Chromosome Res. 1998, 6, 379–383. [Google Scholar] [CrossRef]
- Veyrunes, F.; Chevret, P.; Catalan, J.; Castiglia, R.; Watson, J.; Dobigny, G.; Robinson, T.J.; Britton-Davidian, J. A novel sex determination system in a close relative of the house mouse. Proc. R. Soc. Lond. B Biol. Sci. 2009, 277, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- Zakian, S.M.; Kulbakina, N.A.; Meyer, M.N.; Semenova, L.A.; Bochkarev, M.N.; Radjabli, S.I.; Serov, O.L. Non-random inactivation of the X-chromosome in interspecific hirbid voles. Genet Res. 1987, 50, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, B. Relation between polymorphism of chromosome No. 1 and chromosome number in Microtus mandarinus. J. Shaanxi Norm. Univ. Nat. Sci. Ed. 2000, 28, 83–86. [Google Scholar]
- Hsu, T.C.; Spirito, S.E.; Pardue, M.L. Distribution of 18+28S ribosomal genes in mammalian genomes. Chromosoma 1975, 53, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Proskuryakova, A.; Kulemzina, A.; Perelman, P.; Serdukova, N.; Ryder, O.; Graphodatsky, A. The Case of X and Y Localization of Nucleolus Organizer Regions (NORs) in Tragulus javanicus (Cetartiodactyla, Mammalia). Genes 2018, 9, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazaux, B.; Catalan, J.; Veyrunes, F.; Douzery, E.J.; Britton-Davidian, J. Are ribosomal DNA clusters rearrangement hotspots? A case study in the genus Mus (Rodentia, Muridae). BMC Evol. Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dam’s Karyomorph (number of dams) | Number of Sons/Number of Progeny (%) | Deviation from the Expected = 0.5 |
|---|---|---|
| II (17) | 54/149 (36.2) | χ2 = 11.2; p < 0.001 |
| III (18) | 70/171 (40.9) | χ2 = 5.66; p = 0.017 |
| IV (3) | 7/7 (100) | Fisher’s exact test: p = 0.070 |
| Abbreviation | 2n | Origin | Complex of Sex Chromosomes | Type of Autosome LMAN2 |
|---|---|---|---|---|
| LMAN0f | 47 | laboratory colony | neo-X1/neo-X2/neo-X3 | 2b2b |
| LMAN1f | 47 | laboratory colony | neo-X1/neo-X2/neo-X3 | 2b2b |
| LMAN2f | 47 | laboratory colony | neo-X1/neo-X2/neo-X3 | 2a2b |
| LMAN3f | 47 | laboratory colony | neo-X1/neo-X2/neo-X3 | 2a2b |
| LMAN5f | 47 | laboratory colony | neo-X2/neo-X3/neo-Y | 2a2b |
| LMAN6f | 47 | laboratory colony | neo-X2/neo-X3/neo-Y4 | 2a2a |
| LMAN10m | 48 | laboratory colony | neo-X1/neo-Y/neo-X3/neo-X3 | 2a2a |
| LMAN14f | 47 | wild | neo-X2/neo-X3/neo-Y | 2a2c |
| LMAN15m | 48 | wild | neo-X1/neo-Y/neo-X3/neo-X3 | 2a2c |
| LMAN16f | 47 | wild | neo-X1/neo-X2/neo-X3 | 2a2a |
| LMAN17f | 47 | wild | neo-X1/neo-X2/neo-X3 | 2a2c |
| LMAN19f | 47 | wild | neo-X1/neo-X2/neo-X3 | 2c2c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanenko, S.A.; Smorkatcheva, A.V.; Kovalskaya, Y.M.; Prokopov, D.Y.; Lemskaya, N.A.; Gladkikh, O.L.; Polikarpov, I.A.; Serdyukova, N.A.; Trifonov, V.A.; Molodtseva, A.S.; et al. Complex Structure of Lasiopodomys mandarinus vinogradovi Sex Chromosomes, Sex Determination, and Intraspecific Autosomal Polymorphism. Genes 2020, 11, 374. https://doi.org/10.3390/genes11040374
Romanenko SA, Smorkatcheva AV, Kovalskaya YM, Prokopov DY, Lemskaya NA, Gladkikh OL, Polikarpov IA, Serdyukova NA, Trifonov VA, Molodtseva AS, et al. Complex Structure of Lasiopodomys mandarinus vinogradovi Sex Chromosomes, Sex Determination, and Intraspecific Autosomal Polymorphism. Genes. 2020; 11(4):374. https://doi.org/10.3390/genes11040374
Chicago/Turabian StyleRomanenko, Svetlana A., Antonina V. Smorkatcheva, Yulia M. Kovalskaya, Dmitry Yu. Prokopov, Natalya A. Lemskaya, Olga L. Gladkikh, Ivan A. Polikarpov, Natalia A. Serdyukova, Vladimir A. Trifonov, Anna S. Molodtseva, and et al. 2020. "Complex Structure of Lasiopodomys mandarinus vinogradovi Sex Chromosomes, Sex Determination, and Intraspecific Autosomal Polymorphism" Genes 11, no. 4: 374. https://doi.org/10.3390/genes11040374