Genome-Wide Identification and Expression Profiling Analysis of the Trihelix Gene Family Under Abiotic Stresses in Medicago truncatula


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Identification of the Trihelix Genes in M. truncatula
2.3. Chromosomal Distribution and Gene Duplication Events Analysis
2.4. Gene Structure and Conserved Motifs Analysis
2.5. Phylogenetic and Collinearity Analysis of MtGT genes
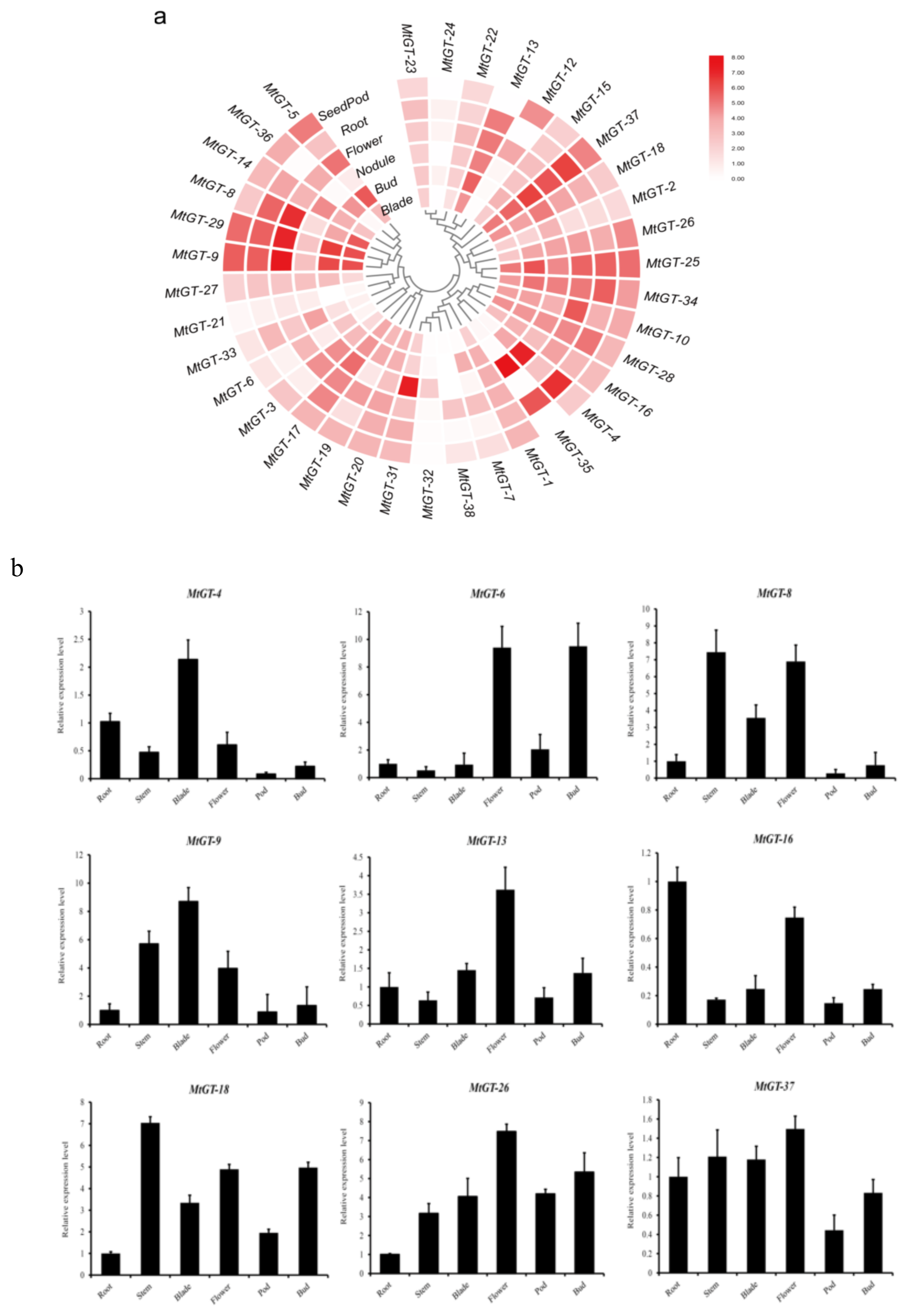
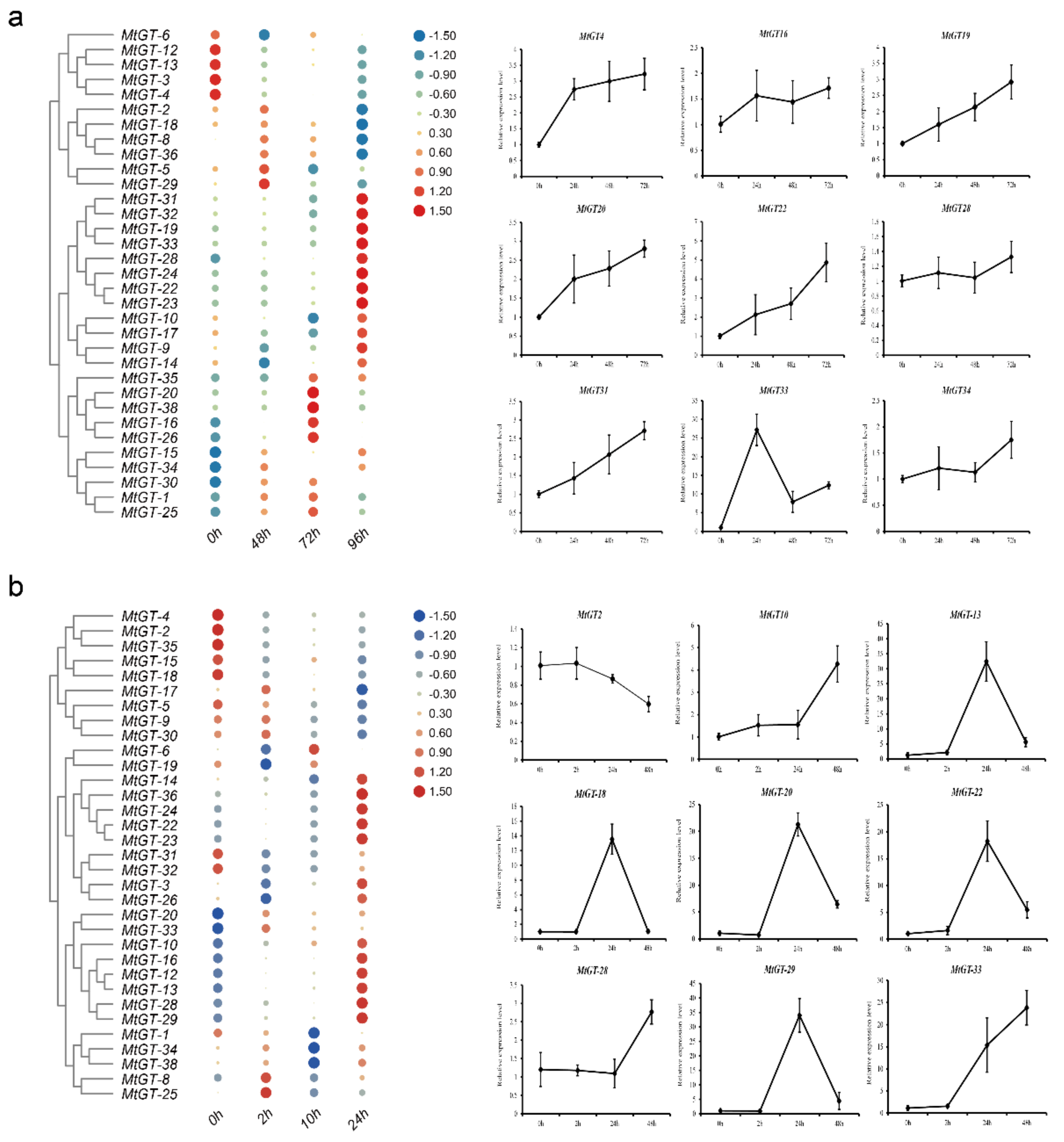
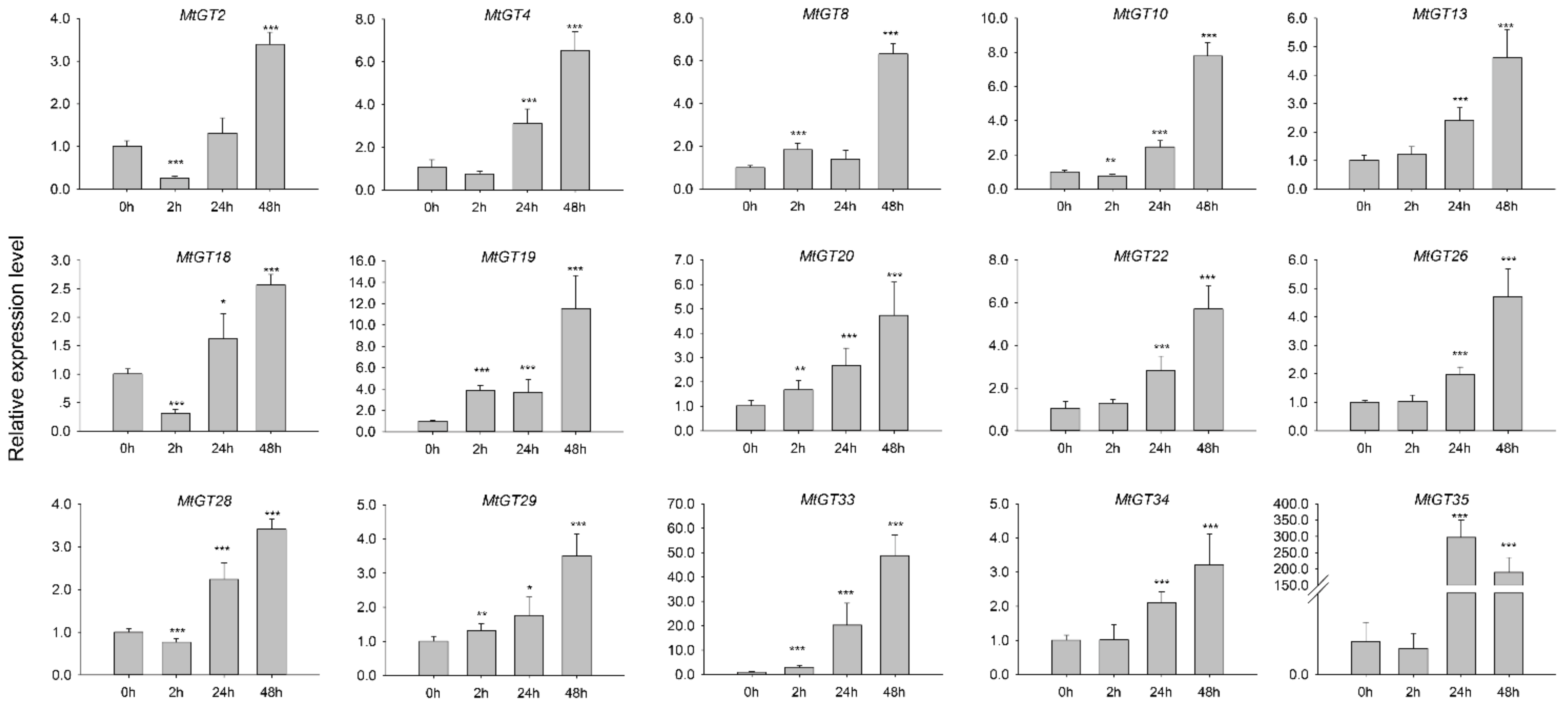
2.6. Tissue-Specific Expression and Abiotic Stress Expression Analysis
2.7. Expression Analysis of the MtGT Genes by Real-Time qPCR
3. Results
3.1. Identification of MtGT Genes in M. truncatula
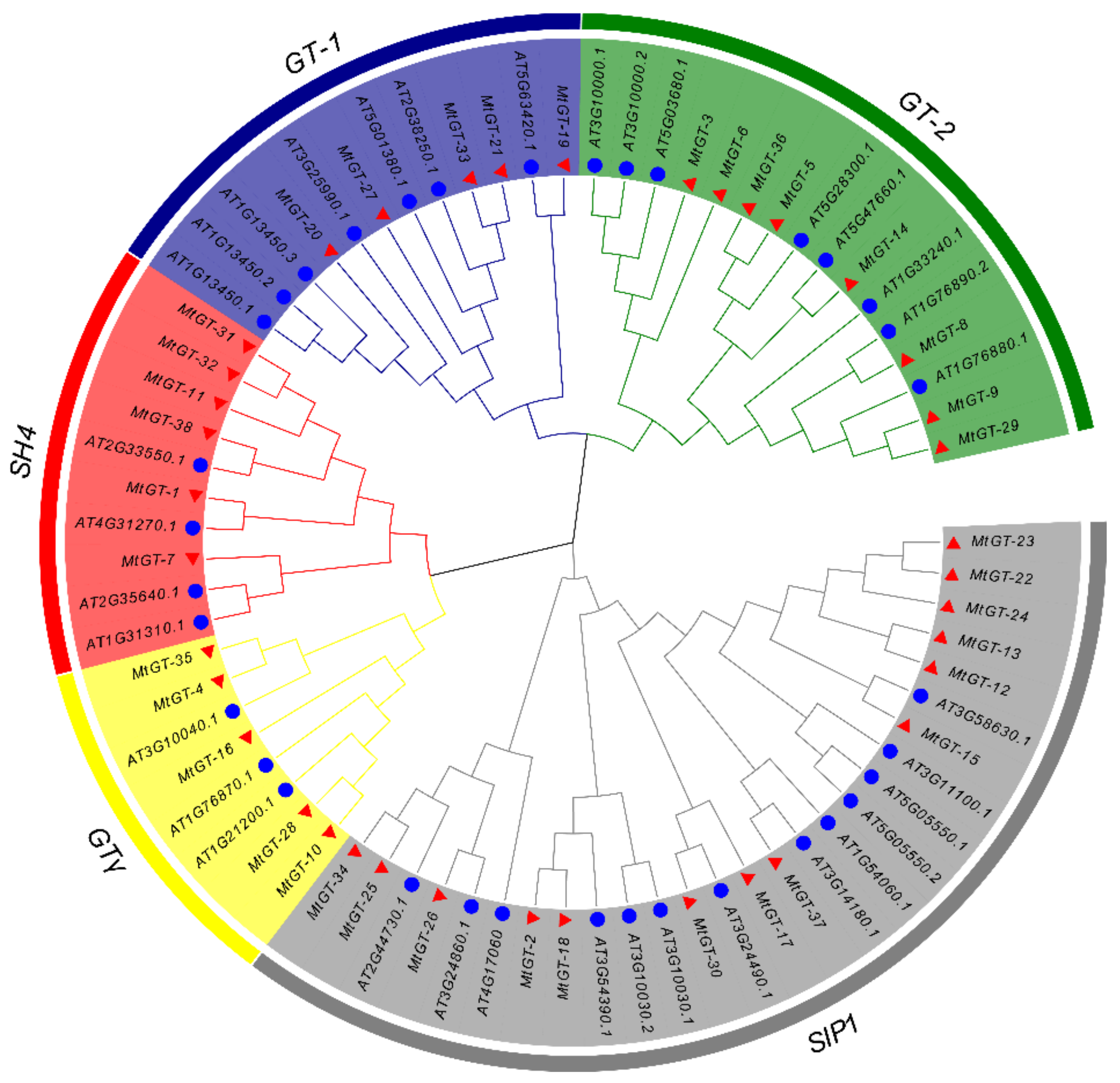
3.2. Phylogenetic Analysis and Classification of MtGT Genes
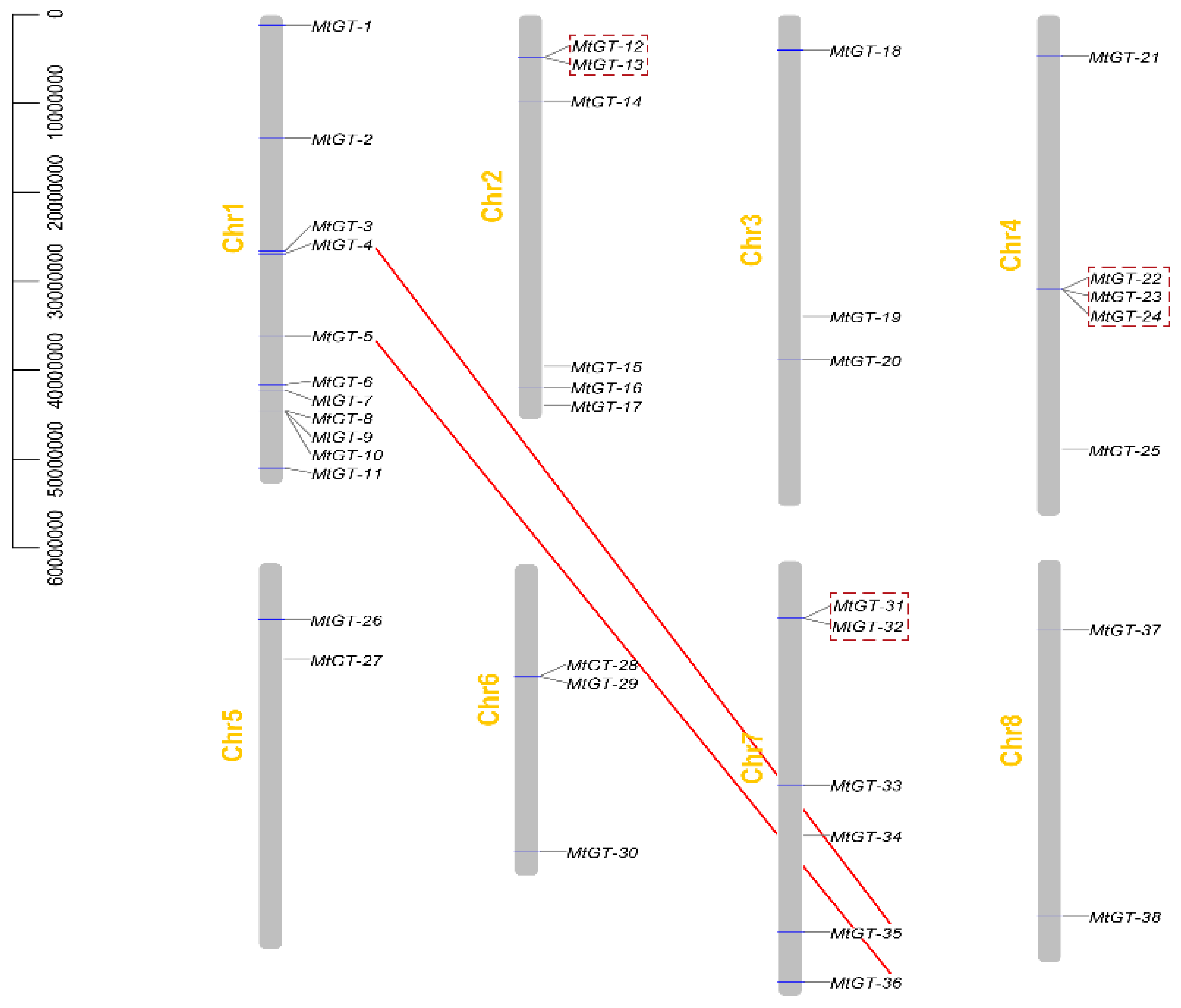
3.3. Chromosomal Distribution and Gene Duplication Events of MtGT Family
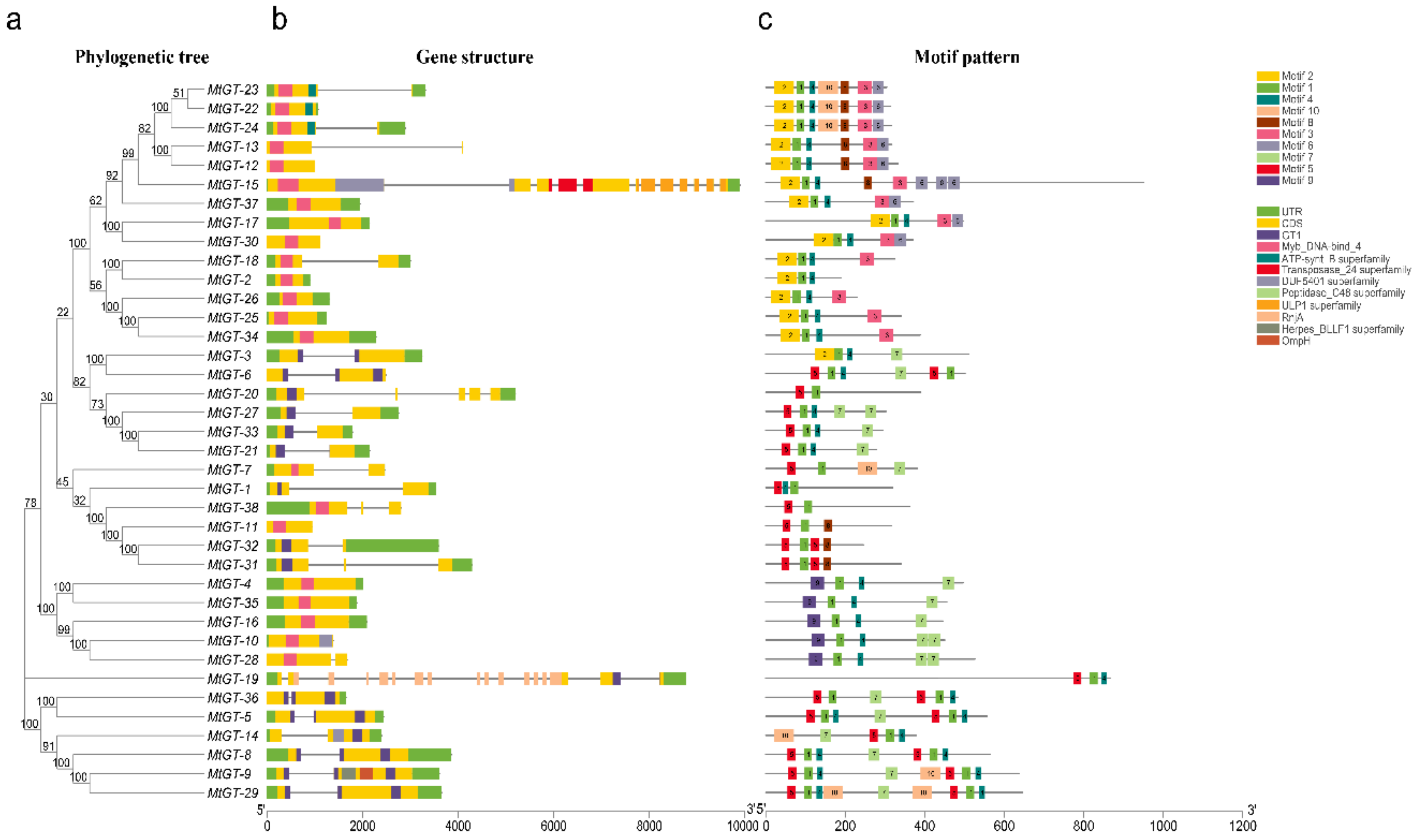
3.4. Gene Structural Characteristics and Conserved Motifs Compositions of MtGT Genes
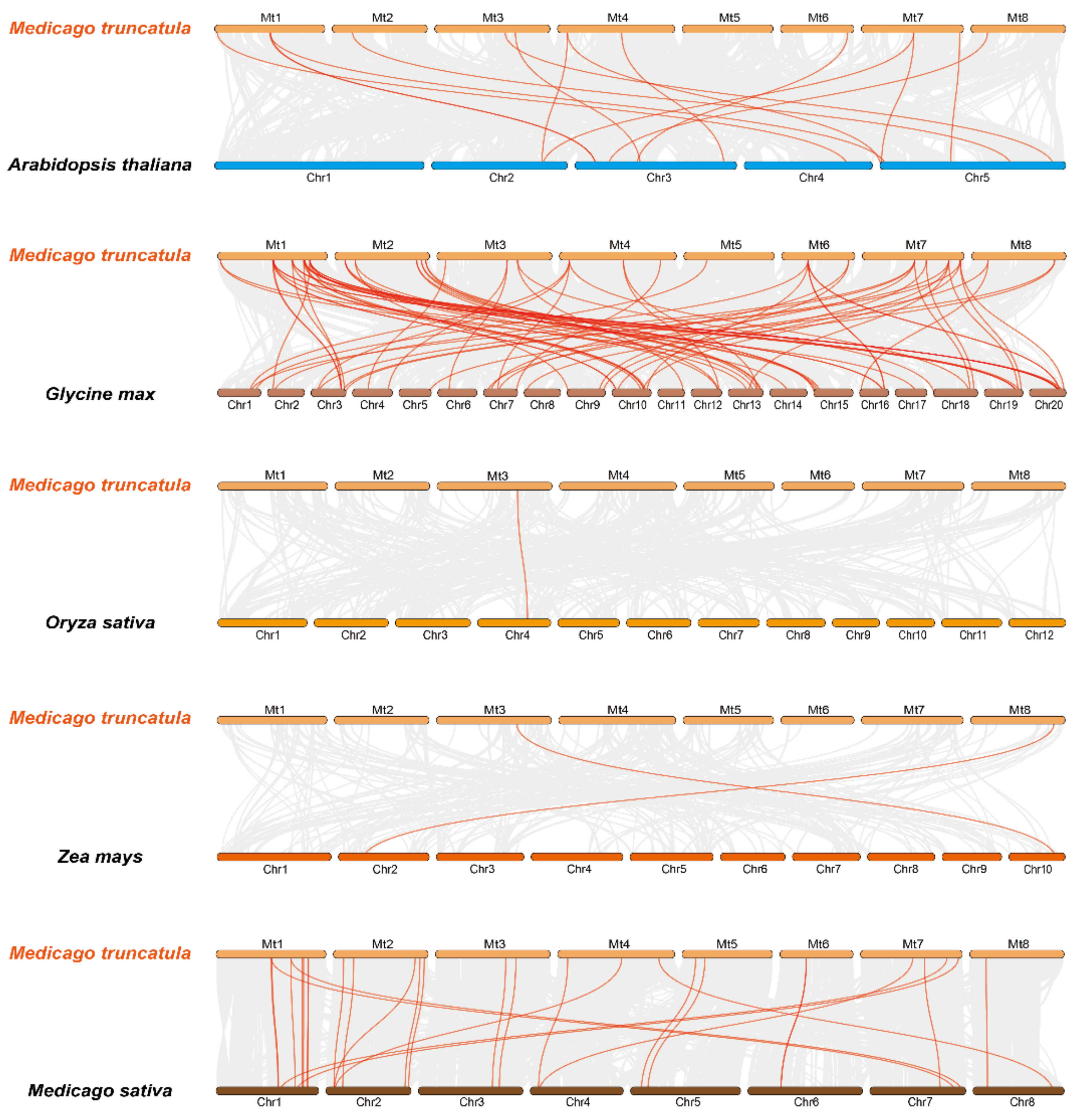
3.5. Evolutionary and Collinearity Analysis within MtGT Genes and Several Species
3.6. Expression Patterns of MtGT Genes in Different Tissues
3.7. Expression Profiling Analysis of MtGT Genes in Response to Abiotic Stress
3.8. Expression Profiling of MtGT Genes in Response to ABA Treatments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wray, G.A.; Hahn, M.W.; Abouheif, E.; Balhoff, J.P.; Pizer, M.; Rockman, M.V.; Romano, L.A. The evolution of transcriptional regulation in eukaryotes. Mol. Biol. Evol. 2003, 20, 1377–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Spitz, F.; Furlong, E.E.M. Transcription factors: From enhancer binding to developmental control. Nat. Rev. Genet. 2012, 13, 613–626. [Google Scholar] [CrossRef]
- Zhou, D.X. Regulatory mechanism of plant gene transcription by GT-elements and GT-factors. Trends Plant Sci. 1999, 4, 210–214. [Google Scholar] [CrossRef]
- Nagano, Y. Several features of the GT-factor trihelix domain resemble those of the Myb DNA-binding domain. Plant Physiol. 2000, 124, 491–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmartin, P.M.; Memelink, J.; Hiratsuka, K.; Kay, S.A.; Chua, N.H. Characterization of a gene encoding a DNA binding protein with specificity for a light-responsive element. Plant Cell 1992, 4, 839–849. [Google Scholar]
- Green, P.J.; Kay, S.A.; Chua, N.H. Sequence-specifc interactions of a pea nuclear factor with light-responsive elements upstream of the rbcS-3A gene. EMBO J. 1987, 6, 2543–2549. [Google Scholar] [CrossRef]
- Perisic, O.; Lam, E. A tobacco DNA binding protein that interacts with a light-responsive box II element. Plant Cell 1992, 4, 831–838. [Google Scholar]
- Hiratsuka, K.; Wu, X.; Fukuzawa, H.; Chua, N.H. Molecular dissection of GT-1 from Arabidopsis. Plant Cell 1994, 6, 1805–1813. [Google Scholar]
- Maréchal, E.; Hiratsuka, K.; Delgado, J.; Nairn, A.; Qin, J.; Chait, B.T.; Chua, N.H. Modulation of GT-1 DNA-binding activity by calcium-dependent phosphorylation. Plant Mol. Biol. 1999, 40, 373–386. [Google Scholar] [CrossRef]
- Dehesh, K.; Smith, L.G.; Tepperman, J.M.; Quail, P.H. Twin autonomous bipartite nuclear localization signals direct nuclear import of GT-2. Plant J. 1995, 8, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Levy, R.N.; Brewer, P.B.; Quon, T.; Smyth, D.R. The trihelix family of transcription factors - light, stress and development. Trends Plant Sci. 2012, 17, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Brewer, P.B.; Howles, P.A.; Dorian, K.; Griffith, M.E.; Ishida, T.; Kaplan-Levy, R.N.; Kilinc, A.; Smyth, D.R. PETAL LOSS, a trihelix transcription factor gene, regulates perianth architecture in the Arabidopsis flower. Development 2004, 131, 4035–4045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuer, C.; Kawamura, A.; Ichikawa, T.; Tominaga-Wada, R.; Wada, T.; Kondou, Y.; Muto, S.; Matsui, M.; Sugimoto, K. The trihelix transcription factor GTL1 regulates ploidy-dependent cell growth in the Arabidopsis trichome. Plant Cell 2009, 21, 2307–2322. [Google Scholar] [CrossRef] [Green Version]
- Yoo, C.Y.; Pence, H.E.; Jin, J.B.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 2010, 22, 4128–4141. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.J.; Lydiate, D.J.; Li, X.; Lui, H.; Gjetvaj, B.; Hegedus, D.D.; Rozwadowski, K. Repression of seed maturation genes by a trihelix transcriptional repressor in Arabidopsis seedlings. Plant Cell 2009, 21, 54–71. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Griffith, M.E.; Li, X.; Zhu, Z.; Tan, L.; Fu, Y.; Zhang, W.; Wang, X.; Xie, D.; Sun, C. Origin of seed shattering in rice (Oryza sativa L.). Planta 2007, 226, 11–20. [Google Scholar] [CrossRef]
- Park, H.C.; Kim, M.L.; Kang, Y.H.; Jeon, J.M.; Yoo, J.H.; Kim, M.C.; Park, C.Y.; Jeong, J.C.; Moon, B.C.; Lee, J.H.; et al. Pathogen-and NaCl-induced expression of the SCaM-4 promoter is mediated in part by a GT-1 box that interacts with a GT-1-like transcription factor. Plant Physiol. 2004, 135, 2150–2161. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Liu, H.; Ji, H.; Wang, Y.; Dong, B.; Qiao, Y.; Liu, M.; Li, X. The wheat GT factor TaGT2L1D negatively regulates drought tolerance and plant development. Sci. Rep. 2016, 6, 27042. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Xie, K.; Hou, X.; Hu, H.; Xiong, L. Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses. Mol. Genet. Genomics 2010, 283, 157–169. [Google Scholar] [CrossRef]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.D.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.X.; Gouzy, J.; Schoof, H.; et al. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cañas, L.A.; Beltrán, J.P. Model legumes: Functional genomics tools in Medicago truncatula. Methods Mol. Biol. 2018, 1822, 11–37. [Google Scholar] [PubMed]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; et al. Genome-wide characterization and identification of trihelix transcription factor and expression profiling in response to abiotic stresses in rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Cai, X.; Ye, Z.; Li, H. Genome-wide identification and expression profiling analysis of trihelix gene family in tomato. Biochem. Biophys. Res. Commun. 2015, 468, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, P.; Liu, T.K.; Ren, H.; Li, Y.; Hou, X. Genome-wide analysis and expression divergence of the trihelix family in Brassica Rapa: Insight into the evolutionary patterns in plants. Sci. Rep. 2017, 7, 6463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, H.; Wang, L.; Ma, S.; Yu, D.; Lu, L.; Yang, Z.; Yang, Z.; Li, F. Transcriptome profiling of Gossypium arboreum during fiber initiation and the genome-wide identification of trihelix transcription factors. Gene 2019, 709, 36–47. [Google Scholar] [CrossRef]
- Osorio, M.B.; Bücker-Neto, L.; Castilhos, G.; Turchetto-Zolet, A.C.; Wiebke-Strohm, B.; Bodanese-Zanettini, M.H.; Margis-Pinheiro, M. Identification and in silico characterization of soybean trihelix-GT and bHLH transcription factors involved in stress responses. Genet. Mol. Biol. 2012, 35, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Song, A.; Wu, D.; Fan, Q.; Tian, C.; Chen, S.; Guan, Z.; Xin, J.; Zhao, K.; Chen, F. Transcriptome-wide identification and expression profiling analysis of chrysanthemum trihelix transcription factors. Int. J. Mol. Sci. 2016, 17, 198. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Hu, R.; Gu, T.; Han, J.; Qiu, D.; Su, P.; Feng, J.; Chang, J.; Yang, G.; He, G. Genome-wide identification and expression profiling of trihelix gene family under abiotic stresses in wheat. BMC Genomics 2019, 20, 287. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Q.; Wang, H.; Zhang, H.; Xu, X.; Li, C.; Yang, C. Comprehensive analysis of trihelix genes and their expression under biotic and abiotic stresses in Populus trichocarpa. Sci. Rep. 2016, 6, 36274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Jin, J.; Tang, L.; Zhao, Y.; Gu, X.; Gao, G.; Luo, J. PlantTFDB 2.0: Update and improvement of the comprehensive plant transcription factor database. Nucleic Acids Res. 2011, 39, D1114–D1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Benedito, V.A.; Wang, M.; Murray, J.D.; Zhao, P.X.; Tang, Y.; Udvardi, M.K. The Medicago truncatula gene expression atlas web server. BMC Bioinform. 2009, 10, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Innan, H.; Kondrashov, F. The evolution of gene duplications: Classifying and distinguishing between models. Nat. Rev. Genet. 2010, 11, 97–108. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, M.; Sun, W.; Huang, L.; Wu, Q.; Bu, T.; Li, C.; Chen, H. Genome-wide identification and expression analysis of the trihelix transcription factor family in tartary buckwheat (Fagopyrum tataricum). BMC Plant Biol. 2019, 19, 344. [Google Scholar] [CrossRef]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, T.; Nakashima, K.; Miyakawa, T.; Kuromori, T.; Tanokura, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular basis of the core regulatory network in ABA responses: Sensing, signaling and transport. Plant Cell Physiol. 2010, 51, 1821–1839. [Google Scholar] [CrossRef]
- Willmann, M.R.; Mehalick, A.J.; Packer, R.L.; Jenik, P.D. MicroRNAs regulate the timing of embryo maturation in Arabidopsis. Plant Physiol. 2011, 155, 1871–1884. [Google Scholar] [CrossRef] [Green Version]
- Ayadi, M.; Delaporte, V.; Li, Y.F.; Zhou, D.X. Analysis of GT-3a identifies a distinct subgroup of trihelix DNA-binding transcription factors in Arabidopsis. FEBS Lett. 2004, 562, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Tzafrir, I.; Pena-Muralla, R.; Dickerman, A.; Berg, M.; Rogers, R.; Hutchens, S.; Sweeney, T.C.; McElver, J.; Aux, G.; Patton, D.; et al. Identification of genes required for embryo development in Arabidopsis. Plant Physiol. 2004, 135, 1206–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagnussat, G.C.; Yu, H.J.; Ngo, Q.A.; Rajani, S.; Mayalagu, S.; Johnson, C.S.; Capron, A.; Xie, L.F.; Ye, D.; Sundaresan, V. Genetic and molecular identification of genes required for female gametophyte development and function in Arabidopsis. Development 2005, 132, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Song, L.; Song, J.; Ouyang, B.; Guo, L.; Shang, L.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. ShCIGT, a trihelix family gene, mediates cold and drought tolerance by interacting with SnRK1 in tomato. Plant Sci. 2018, 270, 140–149. [Google Scholar] [CrossRef]
- Xi, J.; Qiu, Y.; Du, L.; Poovaiah, B.W. Plant-specific trihelix transcription factor AtGT2L interacts with calcium/calmodulin and responds to cold and salt stresses. Plant Sci. 2012, 185–186, 274–280. [Google Scholar] [CrossRef]
- Xu, H.; Shi, X.; He, L.; Guo, Y.; Zang, D.; Li, H.; Zhang, W.; Wang, Y. Arabidopsis thaliana trihelix transcription factor AST1 mediates abiotic stress tolerance by binding to a novel AGAG-box and some GT motifs. Plant Cell Physiol. 2018, 59, 946–965. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Length (aa) | Signature Domain | MW (KDa) | Subcellular Localization | pI | GRAVY |
|---|---|---|---|---|---|---|---|
| MtGT-1 | Medtr1g009220.1 | 319 | 19–104 | 37,270.73 | Nucleus | 5.36 | −1.039 |
| MtGT-2 | Medtr1g037650.1 | 189 | 35–122 | 21,313.82 | Nucleus | 9.14 | −0.917 |
| MtGT-3 | Medtr1g060990.1 | 511 | 129–217 | 58,779.54 | Nucleus | 6.57 | −1.173 |
| MtGT-4 | Medtr1g061640.1 | 496 | 121–247 | 56,941.93 | Nucleus | 6.46 | −1.242 |
| MtGT-5 | Medtr1g081180.1 | 557 | 101–181 | 64,373.39 | Nucleus | 5.71 | −1.104 |
| MtGT-6 | Medtr1g492790.1 | 501 | 112–200 | 57,848.24 | Nucleus | 5.99 | −0.921 |
| MtGT-7 | Medtr1g094045.1 | 381 | 53–183 | 42,978.72 | Extracell/Nucleus | 9.09 | −1.072 |
| MtGT-8 | Medtr1g098870.1 | 565 | 53–140 | 64,378.61 | Nucleus | 5.91 | −1.088 |
| MtGT-9 | Medtr1g098900.1 | 637 | 55–142 | 70,838.09 | Nucleus | 5.77 | −0.900 |
| MtGT-10 | Medtr1g098920.1 | 450 | 123–250 | 51,386.17 | Nucleus | 6.16 | −1.049 |
| MtGT-11 | Medtr1g112650.1 | 316 | 40–131 | 35,249.58 | Chloroplast/Nucleus | 4.97 | −0.653 |
| MtGT-12 | Medtr2g016140.1 | 332 | 20–114 | 37,437.82 | Chloroplast | 5.37 | −0.703 |
| MtGT-13 | Medtr2g016140.2 | 317 | 20–114 | 35,816.88 | Chloroplast/Nucleus | 5.29 | −0.805 |
| MtGT-14 | Medtr2g026725.1 | 379 | 260–348 | 42,411.51 | Nucleus | 7.74 | −0.924 |
| MtGT-15 | Medtr2g092960.1 | 1223 | 43–136 | 140,855.87 | Nucleus | 8.51 | −0.845 |
| MtGT-16 | Medtr2g098080.1 | 445 | 112–238 | 50,604.51 | Nucleus | 6.21 | −0.991 |
| MtGT-17 | Medtr2g102227.1 | 496 | 272–360 | 57,303.69 | Nucleus | 4.48 | −1.395 |
| MtGT-18 | Medtr3g014280.1 | 325 | 35–122 | 37,084.54 | Nucleus | 9.03 | −1.057 |
| MtGT-19 | Medtr3g089020.1 | 867 | 773–857 | 96,355.08 | Chloroplast | 7.97 | −0.400 |
| MtGT-20 | Medtr3g085960.1 | 390 | 74–163 | 44,613.12 | Chloroplast/Nucleus/Peroxisome | 5.90 | −0.834 |
| MtGT-21 | Medtr4g015680.1 | 279 | 40–125 | 33,950.09 | Nucleus | 6.55 | −1.283 |
| MtGT-22 | Medtr4g079960.1 | 313 | 28–123 | 35,566.23 | Nucleus | 9.53 | −0.778 |
| MtGT-23 | Medtr4g079960.2 | 304 | 28–123 | 34,456.87 | Nucleus | 9.60 | −0.818 |
| MtGT-24 | Medtr4g079960.3 | 315 | 28–123 | 35,784.64 | Nucleus | 9.73 | −0.774 |
| MtGT-25 | Medtr4g117990.1 | 340 | 41–135 | 37,889.76 | Nucleus | 9.07 | −0.915 |
| MtGT-26 | Medtr5g017500.1 | 229 | 20–115 | 26,637.29 | Nucleus | 9.22 | −0.834 |
| MtGT-27 | Medtr5g026540.1 | 302 | 43–128 | 35,678.99 | Nucleus | 8.34 | −1.208 |
| MtGT-28 | Medtr6g035315.1 | 526 | 117–243 | 59,762.71 | Nucleus | 6.05 | −0.988 |
| MtGT-29 | Medtr6g035370.1 | 646 | 53–140 | 72,115.32 | Nucleus | 5.74 | −1.018 |
| MtGT-30 | Medtr6g486270.1 | 370 | 127–218 | 42,750.57 | Nucleus | 4.59 | −1.147 |
| MtGT-31 | Medtr7g020870.1 | 341 | 38–130 | 37,377.48 | Nucleus | 5.32 | −0.740 |
| MtGT-32 | Medtr7g020870.2 | 245 | 38–131 | 26,785.93 | Nucleus | 5.77 | −0.636 |
| MtGT-33 | Medtr7g068770.1 | 293 | 51–136 | 35,338.67 | Nucleus | 6.62 | −1.346 |
| MtGT-34 | Medtr7g081190.1 | 388 | 43–140 | 43,393.75 | Nucleus | 9.64 | −0.969 |
| MtGT-35 | Medtr7g103390.1 | 455 | 101–227 | 52,293.03 | Nucleus | 6.02 | −1.204 |
| MtGT-36 | Medtr7g114860.1 | 483 | 379–476 | 55,781.34 | Nucleus | 6.34 | −0.832 |
| MtGT-37 | Medtr8g022290.1 | 371 | 65–161 | 40,319.8 | Nucleus | 9.53 | −0.803 |
| MtGT-38 | Medtr8g100130.1 | 361 | 45–139 | 40,525.42 | Nucleus | 5.36 | −0.803 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhang, H.; Ma, L.; Wang, Z.; Wang, K. Genome-Wide Identification and Expression Profiling Analysis of the Trihelix Gene Family Under Abiotic Stresses in Medicago truncatula. Genes 2020, 11, 1389. https://doi.org/10.3390/genes11111389
Liu X, Zhang H, Ma L, Wang Z, Wang K. Genome-Wide Identification and Expression Profiling Analysis of the Trihelix Gene Family Under Abiotic Stresses in Medicago truncatula. Genes. 2020; 11(11):1389. https://doi.org/10.3390/genes11111389
Chicago/Turabian StyleLiu, Xiqiang, Han Zhang, Lin Ma, Zan Wang, and Kun Wang. 2020. "Genome-Wide Identification and Expression Profiling Analysis of the Trihelix Gene Family Under Abiotic Stresses in Medicago truncatula" Genes 11, no. 11: 1389. https://doi.org/10.3390/genes11111389