Genome-Wide Transcriptome Profiling Provides Insight on Cholesterol and Lithocholate Degradation Mechanisms in Nocardioides simplex VKM Ac-2033D

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Microorganism, Cultivation, and Gene Expression Induction
2.3. Growth Estimations
2.4. Steroid Bioconversion
2.5. Steroid Metabolite Isolation
2.6. Thin Layer Chromatography (TLC)
2.7. High-Performance Liquid Chromatography (HPLC)
2.8. Mass-Spectrometry (MS), 1H-NMR and 13C-NMR Spectroscopy
2.9. Isolation of mRNA and High-Throughput Sequencing
2.10. Computational Analyses
2.11. Search of Transcription Factor Binding Motifs
3. Results
3.1. Growth on Cholesterol and LCA
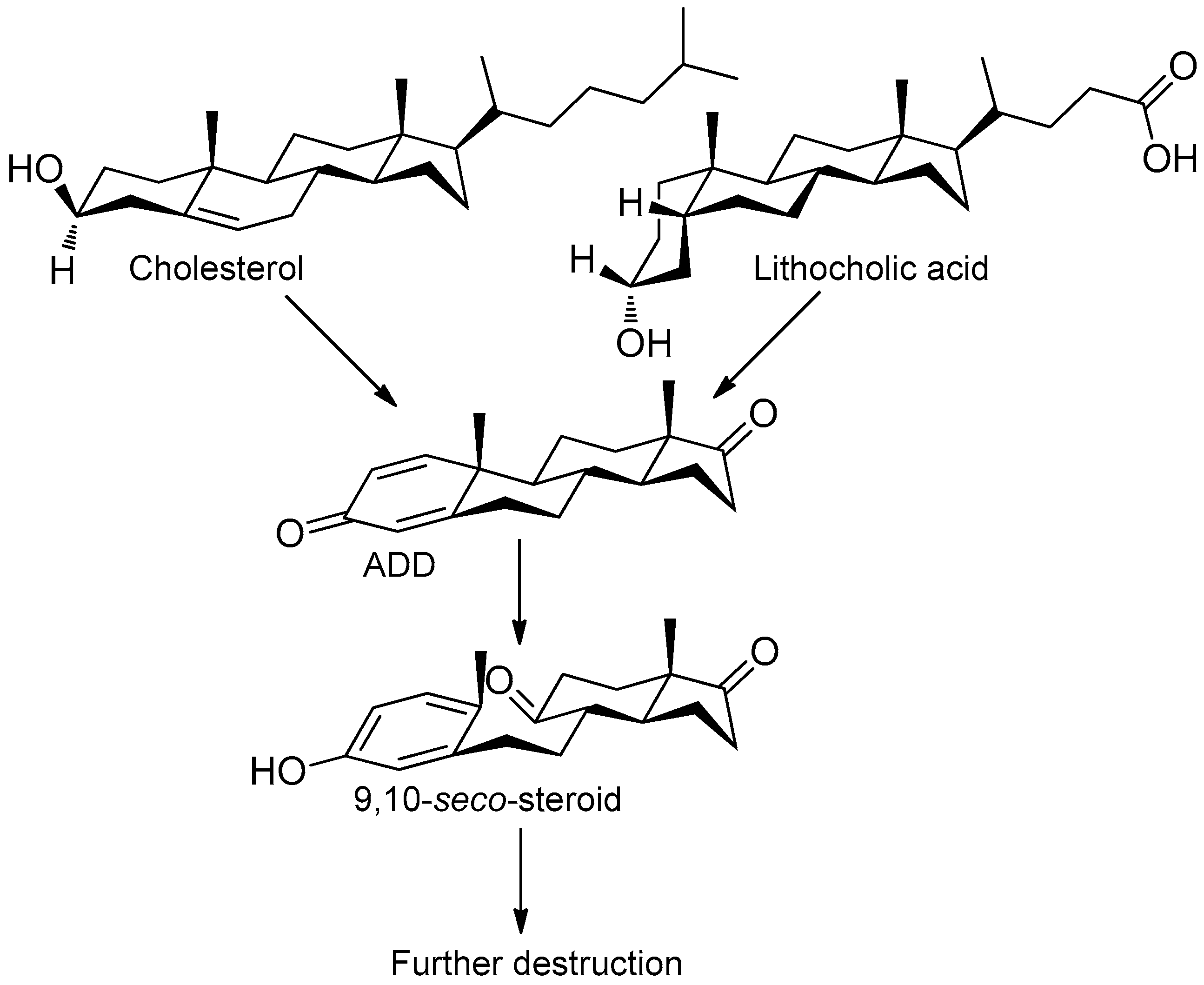
3.2. Cholesterol and LCA Bioconversion
3.3. Transcriptome Sequencing Differently Expressed Genes
3.4. Candidate Motifs for Transcriptional Regulators of LCA Catabolism in N. Simplex
4. Discussion
4.1. Clusters and Regulons
4.2. Initial Steps of Cholesterol Oxidation
4.3. Main Steps of LCA Oxidative Destruction and Cholic Acid Interconversion Genes
4.4. Regulation and Candidate Motifs for LCA Catabolism Transcriptional Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bergstrand, L.H.; Cardenas, E.; Holert, J.; van Hamme, J.D.; Mohn, W.W. Delineation of steroid-degrading microorganisms through comparative genomic analysis. mBio 2016, 7, e00166. [Google Scholar] [CrossRef] [Green Version]
- Philipp, B. Bacterial degradation of bile salts. Appl. Microbiol. Biotechnol. 2011, 89, 903–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef] [PubMed]
- Olivera, E.R.; Luengo, J.M. Steroids as environmental compounds recalcitrant to degradation: Genetic mechanisms of bacterial biodegradation pathways. Genes 2019, 10, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgi, V.; Menéndez, P.; García-Carnelli, C. Microbial transformation of cholesterol: Reactions and practical aspects—An update. World J. Microbiol. Biotechnol. 2019, 35. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xiong, L.-B.; Tao, X.; Liu, Q.-H.; Wang, F.-Q.; Wei, D.-Z. Integrated transcriptome and proteome studies reveal the underlying mechanisms for sterol catabolism and steroid production in Mycobacterium neoaurum. J. Agric. Food Chem. 2018, 66, 9147–9157. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-H.; Xu, L.-Q.; Yao, K.; Xiong, L.-B.; Tao, X.-Y.; Liu, M.; Wang, F.-Q.; Wei, D.-Z. Engineered 3-ketosteroid 9α-hydroxylases in Mycobacterium neoaurum: An efficient platform for production of steroid drugs. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef] [Green Version]
- Van der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef] [Green Version]
- McLeod, M.P.; Warren, R.L.; Hsiao, W.W.L.; Araki, N.; Myhre, M.; Fernandes, C.; Miyazawa, D.; Wong, W.; Lillquist, A.L.; Wang, D.; et al. The complete genome of Rhodococcus sp. RHA1 provides insights into a catabolic powerhouse. Proc. Natl. Acad. Sci. USA 2006, 103, 15582–15587. [Google Scholar] [CrossRef] [Green Version]
- Uhía, I.; Galán, B.; Kendall, S.L.; Stoker, N.G.; García, J.L. Cholesterol metabolism in Mycobacterium smegmatis: Cholesterol pathway. Environ. Microbiol. Rep. 2012, 4, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Drzyzga, O.; Fernández de las Heras, L.; Morales, V.; Navarro Llorens, J.M.; Perera, J. Cholesterol degradation by Gordonia cholesterolivorans. Appl. Environ. Microbiol. 2011, 77, 4802–4810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, M.; Koshino, H.; Malon, M.; Hirota, H.; Hayashi, T. Steroid degradation in Comamonas testosteroni TA441: Identification of the entire β-oxidation cycle of the cleaved B ring. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Mohn, W.W.; Wilbrink, M.H.; Casabon, I.; Stewart, G.R.; Liu, J.; van der Geize, R.; Eltis, L.D. Gene cluster encoding cholate catabolism in Rhodococcus spp. J. Bacteriol. 2012, 194, 6712–6719. [Google Scholar] [CrossRef] [Green Version]
- Philipp, B.; Erdbrink, H.; Suter, M.J.-F.; Schink, B. Degradation of and sensitivity to cholate in Pseudomonas sp. strain Chol1. Arch. Microbiol. 2006, 185, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Mendelski, M.N.; Dölling, R.; Feller, F.M.; Hoffmann, D.; Ramos Fangmeier, L.; Ludwig, K.C.; Yücel, O.; Mährlein, A.; Paul, R.J.; Philipp, B. Steroids originating from bacterial bile acid degradation affect Caenorhabditis elegans and indicate potential risks for the fauna of manured soils. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Luengo, J.M.; Olivera, E.R. Identification and characterization of the genes and enzymes belonging to the bile acid catabolic pathway in Pseudomonas. In Microbial Steroids; Barredo, J.-L., Herráiz, I., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1645, pp. 109–142. ISBN 978-1-4939-7182-4. [Google Scholar]
- Fokina, V.V.; Sukhodolskaya, G.V.; Baskunov, B.P.; Turchin, K.F.; Grinenko, G.S.; Donova, M.V. Microbial conversion of pregna-4,9(11)-diene-17α,21-diol-3,20-dione acetates by Nocardioides simplex VKM Ac-2033D. Steroids 2003, 68, 415–421. [Google Scholar] [CrossRef]
- Fokina, V.V.; Donova, M.V. 21-Acetoxy-pregna-4(5),9(11),16(17)-triene-21-ol-3,20-dione conversion by Nocardioides simplex VKM Ac-2033D. J. Steroid Biochem. Mol. Biol. 2003, 87, 319–325. [Google Scholar] [CrossRef]
- Fokina, V.V.; Sukhodol’skaya, G.V.; Gulevskaya, S.A.; Gavrish, E.Y.; Evtushenko, L.I.; Donova, M.V. The 1(2)-dehydrogenation of steroid substrates by Nocardioides simplex VKM Ac-2033D. Microbiology 2003, 72, 24–29. [Google Scholar] [CrossRef]
- Sukhodolskaya, G.; Fokina, V.; Shutov, A.; Nikolayeva, V.; Savinova, T.; Grishin, Y.; Kazantsev, A.; Lukashev, N.; Donova, M. Bioconversion of 6-(N-methyl-N-phenyl)aminomethyl androstane steroids by Nocardioides simplex. Steroids 2017, 118, 9–16. [Google Scholar] [CrossRef]
- Deshcherevskaya, N.O.; Lobastova, T.G.; Kollerov, V.V.; Kazantsev, A.V.; Donova, M.V. Search and discovery of actinobacteria capable of transforming deoxycholic and cholic acids. J. Mol. Catal. B Enzym. 2016, 133, S157–S165. [Google Scholar] [CrossRef]
- Shtratnikova, V.Y.; Schelkunov, M.I.; Pekov, Y.A.; Fokina, V.V.; Logacheva, M.D.; Sokolov, S.L.; Bragin, E.Y.; Ashapkin, V.V.; Donova, M.V. Complete genome sequence of steroid-transforming Nocardioides simplex VKM Ac-2033D. Genome Announc. 2015, 3, e01406–e01414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shtratnikova, V.Y.; Schelkunov, M.I.; Fokina, V.V.; Pekov, Y.A.; Ivashina, T.; Donova, M.V. Genome-wide bioinformatics analysis of steroid metabolism-associated genes in Nocardioides simplex VKM Ac-2033D. Curr. Genet. 2016, 62, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Park, N.S.; Myeong, J.S.; Park, H.J.; Han, K.B.; Kim, S.N.; Kim, E.S. Characterization and culture optimization of regiospecific cyclosporin hydroxylation in rare actinomycetes species. J. Microbiol. Biotechnol. 2005, 15, 188–191. [Google Scholar]
- Lobastova, T.G.; Khomutov, S.M.; Shutov, A.A.; Donova, M.V. Microbiological synthesis of stereoisomeric 7(α/β)-hydroxytestololactones and 7(α/β)-hydroxytestolactones. Appl. Microbiol. Biotechnol. 2019, 103, 4967–4976. [Google Scholar] [CrossRef]
- Bragin, E.Y.; Shtratnikova, V.Y.; Schelkunov, M.I.; Dovbnya, D.V.; Donova, M.V. Genome-wide response on phytosterol in 9-hydroxyandrostenedione-producing strain of Mycobacterium sp. VKM Ac-1817D. BMC Biotechnol. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- McClure, R.; Balasubramanian, D.; Sun, Y.; Bobrovskyy, M.; Sumby, P.; Genco, C.A.; Vanderpool, C.K.; Tjaden, B. Computational analysis of bacterial RNA-Seq data. Nucleic Acids Res. 2013, 41, e140. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreit, J. Aerobic catabolism of sterols by microorganisms: Key enzymes that open the 3-ketosteroid nucleus. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Fernández de las Heras, L. Estudio Genético y Bioquímico de la Ruta de Degradación del Colesterol en Rhodococcus spp; Universidad Complutense de Madrid: Madrid, Spain, 2013. [Google Scholar]
- Qin, H.-M.; Wang, J.-W.; Guo, Q.; Li, S.; Xu, P.; Zhu, Z.; Sun, D.; Lu, F. Refolding of a novel cholesterol oxidase from Pimelobacter simplex reveals dehydrogenation activity. Protein Expr. Purif. 2017, 139, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fernández de las Heras, L.; Perera, J.; Navarro Llorens, J.M. Cholesterol to cholestenone oxidation by ChoG, the main extracellular cholesterol oxidase of Rhodococcus ruber strain Chol-4. J. Steroid Biochem. Mol. Biol. 2014, 139, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.; Kanwar, S.S. Cholesterol oxidase: Source, properties and applications. Insights Enzym. Res. 2017, 1, 5–17. [Google Scholar] [CrossRef]
- Jang, D.S.; Choi, G.; Cha, H.J.; Shin, S.; Hong, B.H.; Lee, H.J.; Lee, H.C.; Choi, K.Y. Contribution of a low-barrier hydrogen bond to catalysis Is not significant in ketosteroid isomerase. Mol. Cells 2015, 38, 409–415. [Google Scholar] [CrossRef]
- Horinouchi, M.; Kurita, T.; Yamamoto, T.; Hatori, E.; Hayashi, T.; Kudo, T. Steroid degradation gene cluster of Comamonas testosteroni consisting of 18 putative genes from meta-cleavage enzyme gene tesB to regulator gene tesR. Biochem. Biophys. Res. Commun. 2004, 324, 597–604. [Google Scholar] [CrossRef]
- Knol, J.; Bodewits, K.; Hessels, G.I.; Dijkhuizen, L.; van der Geize, R. 3-Keto-5α-steroid Δ1-dehydrogenase from Rhodococcus erythropolis SQ1 and its orthologue in Mycobacterium tuberculosis H37Rv are highly specific enzymes that function in cholesterol catabolism. Biochem. J. 2008, 410, 339. [Google Scholar] [CrossRef] [Green Version]
- Guevara, G.; Fernández de las Heras, L.; Perera, J.; Navarro Llorens, J.M. Functional differentiation of 3-ketosteroid Δ1-dehydrogenase isozymes in Rhodococcus ruber strain Chol-4. Microb. Cell Factories 2017, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capyk, J.K.; Kalscheuer, R.; Stewart, G.R.; Liu, J.; Kwon, H.; Zhao, R.; Okamoto, S.; Jacobs, W.R.; Eltis, L.D.; Mohn, W.W. Mycobacterial cytochrome p450 125 (cyp125) catalyzes the terminal hydroxylation of c27 steroids. J. Biol. Chem. 2009, 284, 35534–35542. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.-Q.; Liu, Y.-J.; Yao, K.; Liu, H.-H.; Tao, X.-Y.; Wang, F.-Q.; Wei, D.-Z. Unraveling and engineering the production of 23,24-bisnorcholenic steroids in sterol metabolism. Sci. Rep. 2016, 6, 21928. [Google Scholar] [CrossRef] [Green Version]
- Capyk, J.K.; D’Angelo, I.; Strynadka, N.C.; Eltis, L.D. Characterization of 3-ketosteroid 9α-hydroxylase, a Rieske oxygenase in the cholesterol degradation pathway of Mycobacterium tuberculosis. J. Biol. Chem. 2009, 284, 9937–9946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oosterwijk, N.; Knol, J.; Dijkhuizen, L.; van der Geize, R.; Dijkstra, B.W. Structure and catalytic mechanism of 3-ketosteroid-Δ4-(5α)-dehydrogenase from Rhodococcus jostii RHA1 genome. J. Biol. Chem. 2012, 287, 30975–30983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatta, T.; Wakabayashi, T.; Itagaki, E. 3-Keto-5α-steroid-Δ4-dehydrogenase from Nocardia corallina: Purification and characterization. J. Biochem. 1991, 109, 581–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plesiat, P.; Grandguillot, M.; Harayama, S.; Vragar, S.; Michel-Briand, Y. Cloning, sequencing, and expression of the Pseudomonas testosteroni gene encoding 3-oxosteroid delta 1-dehydrogenase. J. Bacteriol. 1991, 173, 7219–7227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florin, C.; Köhler, T.; Grandguillot, M.; Plesiat, P. Comamonas testosteroni 3-ketosteroid-delta 4(5α)-dehydrogenase: Gene and protein characterization. J. Bacteriol. 1996, 178, 3322–3330. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Feng, J.; Zhang, D.; Wu, Q.; Zhu, D.; Ma, Y. Characterization of new recombinant 3-ketosteroid-Δ1-dehydrogenases for the biotransformation of steroids. Appl. Microbiol. Biotechnol. 2017, 101, 6049–6060. [Google Scholar] [CrossRef]
- García-Fernández, E.; Medrano, F.J.; Galán, B.; García, J.L. Deciphering the transcriptional regulation of cholesterol catabolic pathway in Mycobacteria: Identification of the inducer of KstR repressor. J. Biol. Chem. 2014, 289, 17576–17588. [Google Scholar] [CrossRef] [Green Version]
- Petrusma, M.; Hessels, G.; Dijkhuizen, L.; van der Geize, R. Multiplicity of 3-ketosteroid-9α-hydroxylase enzymes in Rhodococcus rhodochrous DSM43269 for specific degradation of different classes of steroids. J. Bacteriol. 2011, 193, 3931–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, S.; Kanematsu, Y.; Fujiwara, T. New dehydroxylation reaction observed in the microbiological degradation pathway of cholic acid. Nature 1967, 214, 520–521. [Google Scholar] [CrossRef] [PubMed]
- Mohn, W.W.; van der Geize, R.; Stewart, G.R.; Okamoto, S.; Liu, J.; Dijkhuizen, L.; Eltis, L.D. The actinobacterial mce4 locus encodes a steroid transporter. J. Biol. Chem. 2008, 283, 35368–35374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallonee, D.H.; Adams, J.L.; Hylemon, P.B. The bile acid-inducible baiB gene from Eubacterium sp. strain VPI 12708 encodes a bile acid-coenzyme A ligase. J. Bacteriol. 1992, 174, 2065–2071. [Google Scholar] [CrossRef] [Green Version]
- Mallonee, D.H.; Lijewski, M.A.; Hylemon, P.B. Expression in Escherichia coli and characterization of a bile acid-inducible 3 α-hydroxysteroid dehydrogenase from Eubacterium sp. strain VPI 12708. Curr. Microbiol. 1995, 30, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, M.; Hayashi, T.; Yamamoto, T.; Kudo, T. A new bacterial steroid degradation gene cluster in Comamonas testosteroni TA441 which consists of aromatic-compound degradation genes for seco-steroids and 3-ketosteroid dehydrogenase genes. Appl. Environ. Microbiol. 2003, 69, 4421–4430. [Google Scholar] [CrossRef] [Green Version]
- Mallonee, D.H.; Hylemon, P.B. Sequencing and expression of a gene encoding a bile acid transporter from Eubacterium sp. strain VPI 12708. J. Bacteriol. 1996, 178, 7053–7058. [Google Scholar] [CrossRef] [Green Version]
- Thanassi, D.G.; Cheng, L.W.; Nikaido, H. Active efflux of bile salts by Escherichia coli. J. Bacteriol. 1997, 179, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Elkins, C.A.; Savage, D.C. Identification of genes encoding conjugated bile salt hydrolase and transport in Lactobacillus johnsonii 100-100. J. Bacteriol. 1998, 180, 4344–4349. [Google Scholar] [CrossRef] [Green Version]
- Haußmann, U.; Wolters, D.A.; Fränzel, B.; Eltis, L.D.; Poetsch, A. Physiological adaptation of the Rhodococcus jostii RHA1 membrane proteome to steroids as growth substrates. J. Proteome Res. 2013, 12, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Somalinga, V.; Mohn, W.W. Rhodococcus jostii porin A (RjpA) functions in cholate uptake. Appl. Environ. Microbiol. 2013, 79, 6191–6193. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Kurita, T.; Hayashi, T.; Kudo, T. Steroid degradation genes in Comamonas testosteroni TA441: Isolation of genes encoding a Δ4(5)-isomerase and 3α- and 3β-dehydrogenases and evidence for a 100kb steroid degradation gene hot spot. J. Steroid Biochem. Mol. Biol. 2010, 122, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweet, F.; Samant, B.R. Bifunctional enzyme activity at the same active site: Study of 3-α and 20-β activity by affinity alkylation of 3-α,20-β-hydroxysteroid dehydrogenase with 17-(bromoacetoxy)steroids. Biochemistry 1980, 19, 978–986. [Google Scholar] [CrossRef]
- Franklund, C.V.; Baron, S.F.; Hylemon, P.B. Characterization of the baiH gene encoding a bile acid-inducible NADH:flavin oxidoreductase from Eubacterium sp. strain VPI 12708. J. Bacteriol. 1993, 175, 3002–3012. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.A.; Mallonee, D.H.; Björkhem, I.; Hylemon, P.B. Expression and characterization of a C24 bile acid 7-α-dehydratase from Eubacterium sp. strain VPI 12708 in Escherichia coli. J. Lipid Res. 1996, 37, 1258–1267. [Google Scholar]
- Hayakawa, S.; Kanematsu, Y.; Fujiwara, T. Microbiological degradation of bile acids. Ring a cleavage and 7α,12α-dehydroxylation of cholic acid by Arthrobacter simplex. Biochem. J. 1969, 115, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, S.; Kurokawa, K. Microbiological dehydroxylation of cholic acid. Nature 1963, 199, 490. [Google Scholar] [CrossRef]
- Hayakawa, S.; Samuelsson, B. Transformation of cholic acid in vitro by Corynebacterium simplex. Bile acids and steroids. 132. J. Biol. Chem. 1964, 239, 94–97. [Google Scholar]
- Mukherjee, E.; Banerjee, S.; Mahato, S.B. Transformation of cholic acid by Arthrobacter simplex. Steroids 1993, 58, 484–490. [Google Scholar] [CrossRef]
- Carere, J.; McKenna, S.E.; Kimber, M.S.; Seah, S.Y.K. Characterization of an aldolase-dehydrogenase complex from the cholesterol degradation pathway of Mycobacterium tuberculosis. Biochemistry 2013, 52, 3502–3511. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Kurita, T.; Kudo, T. Identification of 9,17-dioxo-1,2,3,4,10,19-hexanorandrostan-5-oic acid, 4-hydroxy-2-oxohexanoic acid, and 2-hydroxyhexa-2,4-dienoic acid and related enzymes involved in testosterone degradation in Comamonas testosteroni TA441. Appl. Environ. Microbiol. 2005, 71, 5275–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | Molecular Weight | Chemical Structure | Number | Name | Molecular Weight | Chemical Structure |
|---|---|---|---|---|---|---|---|
| V | Cholest-4-en-3-one | 384 |  | VI | Cholesta-1,4-dien-3-one | 382 |  |
| VII | 26-Hydroxy-cholest-4-en-3-one | 400 |  | VIII | 3-Oxo-cholest-4-en-26-oic acid | 414 |  |
| IX | 3-Oxo-cholesta-1,4-dien-26-oic acid | 412 |  | X | 3β-Hydroxy-cholest-5-en-26-oic acid | 416 |  |
| XI | Androst-4-en-17β-ol-3-one, Testosterone | 288 |  | XII | Androsta-1,4-dien-17β-ol-3-one, 1-Dehydrotestosterone | 286 |  |
| XIII | Androst-4-ene-9α,17β-diol-3-one, 9α-Hydroxy-testosterone(preliminarily determined) | 304 |  |
| Cluster | Locus Tags | Cholesterol | LCA | LCA/Cholesterol | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | Ave | Max | D | I | Ave | Max | D | I | Ave | Max | D | ||
| A | KR76_14160-KR76_14505 | 23 | 50 | 226 | 0 | 28 | 40 | 108 | 0 | 0 | 0 | ||
| B | KR76_12190-KR76_12345 | 16 | 14 | 38 | 0 | 19 | 17 | 34 | 0 | 0 | 0 | ||
| C | KR76_25015- KR76_25200 | 2 | 9 | 13 | 0 | 9 | 13 | 65 | 0 | 2 | 5 | 5 | 0 |
| D | KR76_27035-KR76_27130 | 0 | 0 | 1 | 8 | 8 | 1 | 1 | 8 | 8 | 0 | ||
| E | KR76_17985-KR76_18075 | 0 | 1 | 15 | 25 | 44 | 0 | 17 | 31 | 58 | 0 | ||
| Out of clusters | 86 | 19 | 84 | 20 | 24 | 19 | |||||||
| Total | 127 | 20 | 156 | 21 | 44 | 19 | |||||||
| Step | Gene Name | Annotation | Locus Tag | Cluster | Regulon | LCA | Chol | References |
|---|---|---|---|---|---|---|---|---|
| 1 | cho | cholesterol oxidase | KR76_09550 | KstR | + | + | [39] | |
| ksdI, ksi | ∆5-3-ketosteroid isomerase | KR76_23530 | KstR | + | [40] | |||
| KR76_24125 | KstR | + | + | |||||
| 2 | kstD3, tesH | ∆1-3-ketosteroid dehydrogenase with preference for the 1,2-saturated steroid substrates | KR76_14500 | A | KstR | + | + | [41,42,43] |
| 3, 4 | cyp125 | cytochrome P450 125 | KR76_14335 | A | KstR | + | + | [44] |
| 6 | hsd4A | 17β-hydroxysteroid dehydrogenase or β-hydroxyacyl-CoA dehydrogenase | KR76_12260 | B | KstR | + | [45] | |
| 7 | kshA | 3-ketosteroid 9α-hydroxylase, oxygenase subunit | KR76_14170 | A | KstR | + | + | [46] |
| kshB | 3-ketosteroid 9α-hydroxylase, reductase subunit | KR76_14160 | A | KstR | [46] |
| Step | Enzyme Annotation | Gene Name | C. scindens | R. jostii | N. simplex | Cluster | Regulon | LCA | Chol | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| transporter | mce | KR76_12195- KR76_12230 | B | KstR | + | + | [56] | |||
| 1 | bile acid CoA ligase | baiB | HDCHBGLK_01430 | casG | KR76_10030 | KstR | + | + | [57] | |
| KR76_18025 | E | + | ||||||||
| 2 | 3α-hydroxysteroid dehydrogenase | baiA | HDCHBGLK_01433 | KR76_18030 | E | + | [58] | |||
| 3α-(or 20β)-hydroxysteroid dehydrogenase | KR76_12265 | B | KstR | + | + | |||||
| 3α-hydroxysteroid dehydrogenase | KR76_09915 | out | KstR | + | + | |||||
| short-chain alcohol dehydrogenase | KR76_18055 | E | + | [59] | ||||||
| 4 | Δ4-(5α)-3-ketosteroid dehydrogenase | kstD4, tesI | KR76_24505 | out | KstR | + | + | [47,60] | ||
| 5 | Δ1-3-ketosteroid dehydrogenases | kstD3, tesH | KR76_01140 | + | [41,43] | |||||
| KR76_14500 | A | KstR | + | + | ||||||
| 6 | 3-ketosteroid 9α-hydroxylase, oxygenase subunit | kshA | KR76_14170 | A | KstR | + | + | [46] | ||
| KR76_18045 | E | + | ||||||||
| 3-ketosteroid 9α-hydroxylase, reductase subunit | kshB | KR76_14160 | A | KstR | ||||||
| KR76_27080 | D | + | ||||||||
| oxidoreductase | baiCD | casH | KR76_18020 | + | [59] | |||||
| bile acid 7α-dehydratase | baiE | HDCHBGLK_01432 | KR76_16145 | KstR | + | + | [59] | |||
| KR76_18075 | E | + | ||||||||
| flavin-dependent oxidoreductase | ro05794 | KR76_18065 | E | + | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shtratnikova, V.Y.; Schelkunov, M.I.; Fokina, V.V.; Bragin, E.Y.; Lobastova, T.G.; Shutov, A.A.; Kazantsev, A.V.; Donova, M.V. Genome-Wide Transcriptome Profiling Provides Insight on Cholesterol and Lithocholate Degradation Mechanisms in Nocardioides simplex VKM Ac-2033D. Genes 2020, 11, 1229. https://doi.org/10.3390/genes11101229
Shtratnikova VY, Schelkunov MI, Fokina VV, Bragin EY, Lobastova TG, Shutov AA, Kazantsev AV, Donova MV. Genome-Wide Transcriptome Profiling Provides Insight on Cholesterol and Lithocholate Degradation Mechanisms in Nocardioides simplex VKM Ac-2033D. Genes. 2020; 11(10):1229. https://doi.org/10.3390/genes11101229
Chicago/Turabian StyleShtratnikova, Victoria Y., Mikhail I. Schelkunov, Victoria V. Fokina, Eugeny Y. Bragin, Tatyana G. Lobastova, Andrey A. Shutov, Alexey V. Kazantsev, and Marina V. Donova. 2020. "Genome-Wide Transcriptome Profiling Provides Insight on Cholesterol and Lithocholate Degradation Mechanisms in Nocardioides simplex VKM Ac-2033D" Genes 11, no. 10: 1229. https://doi.org/10.3390/genes11101229