Population Genetic Analysis of the Estonian Native Horse Suggests Diverse and Distinct Genetics, Ancient Origin and Contribution from Unique Patrilines

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals for Comparative Analysis
2.3. DNA Isolation and STR Genotyping
2.4. Population Diversity and Structure Analysis
2.5. Y Chromosome and Mitochondrial Variant Screening and Haplotype Analysis
2.6. DMRT3 Gait Genotyping
3. Results
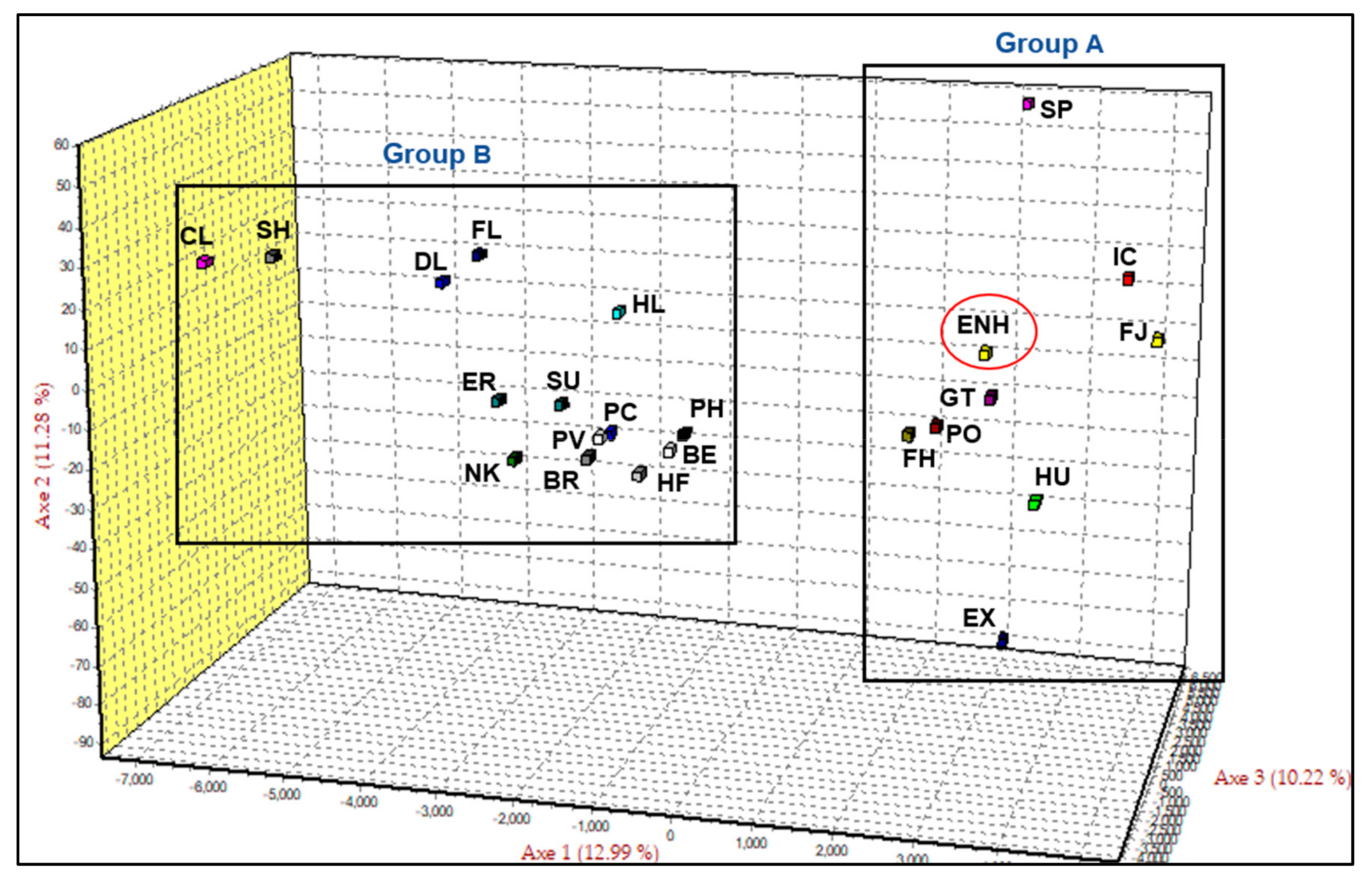
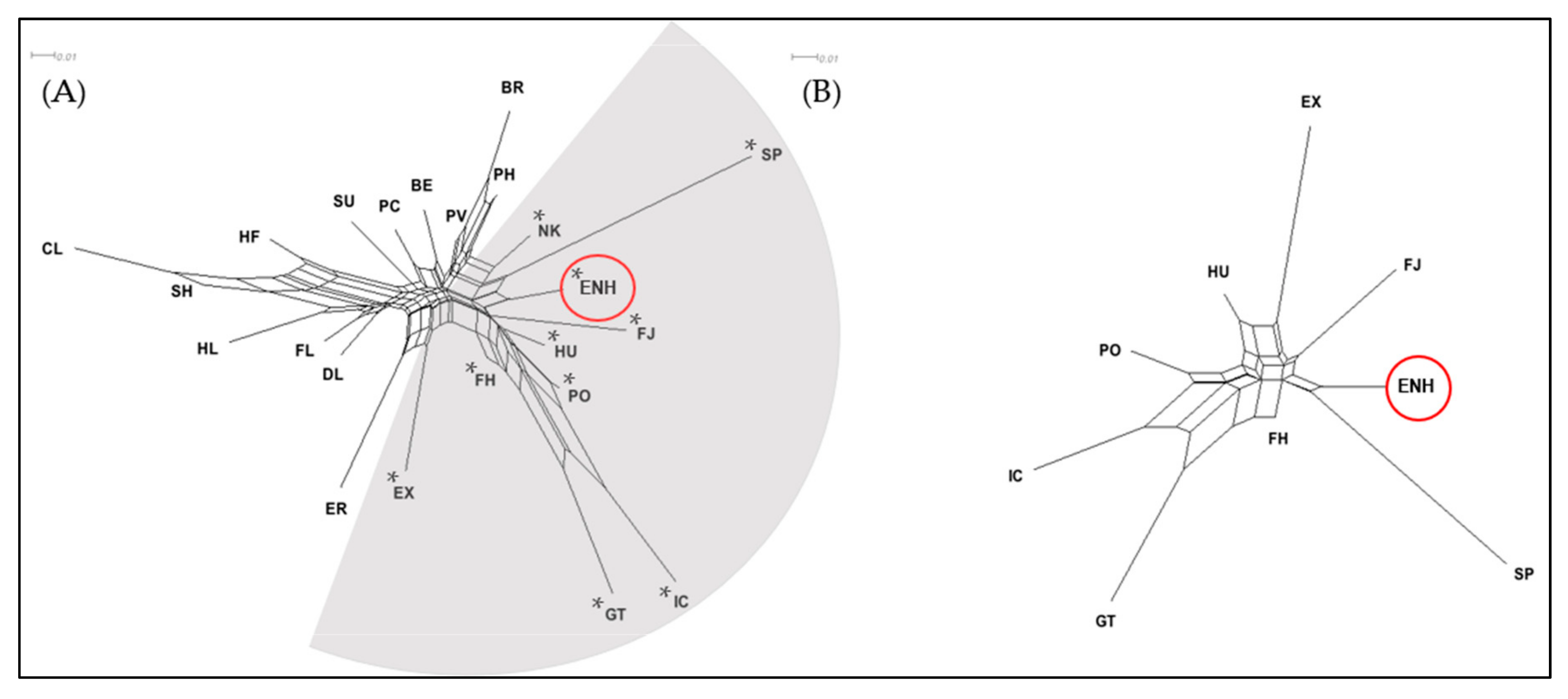
3.1. Genetic Diversity and Relationships of the Estonian Native Horse
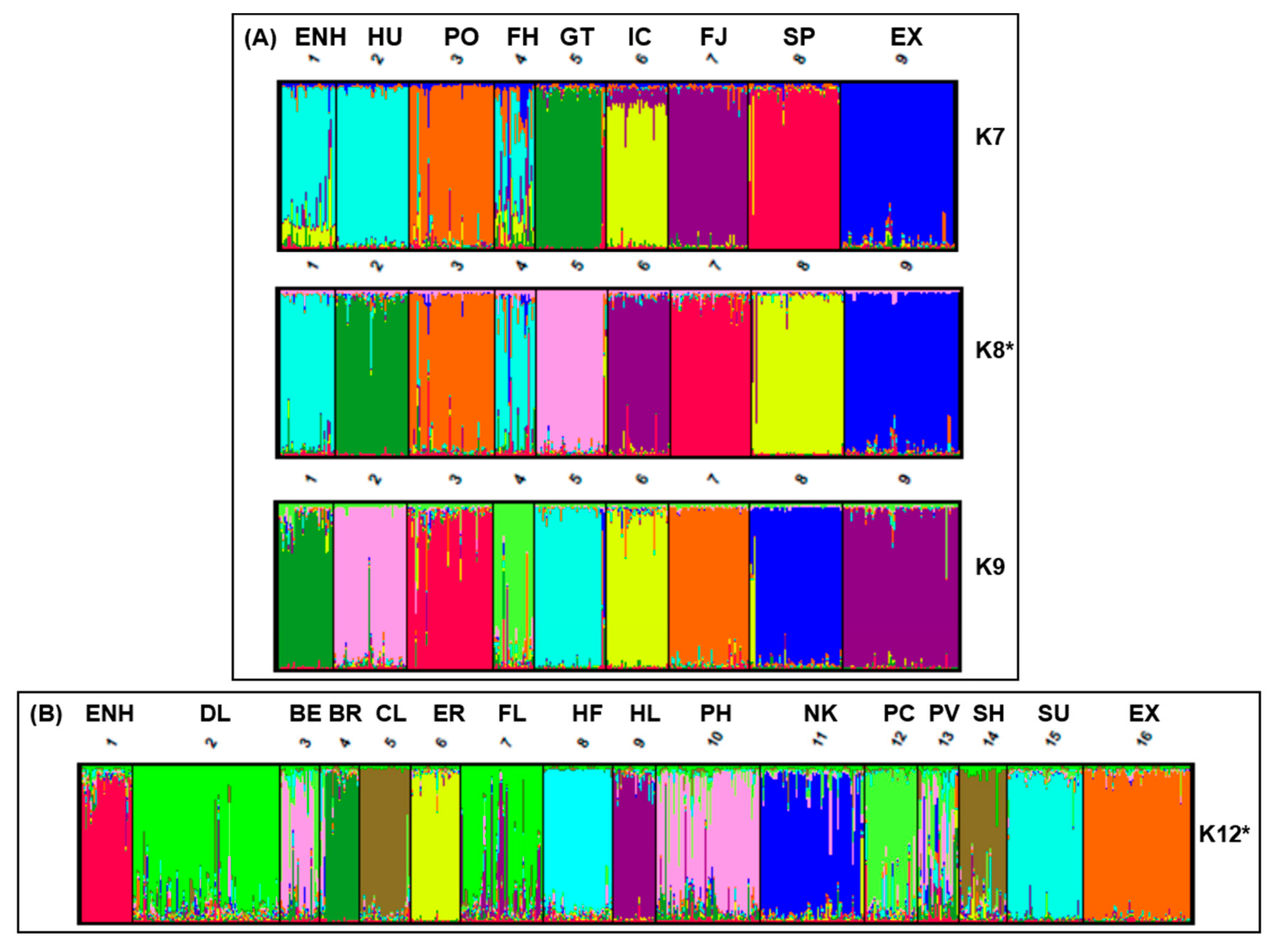
3.2. Population Structure
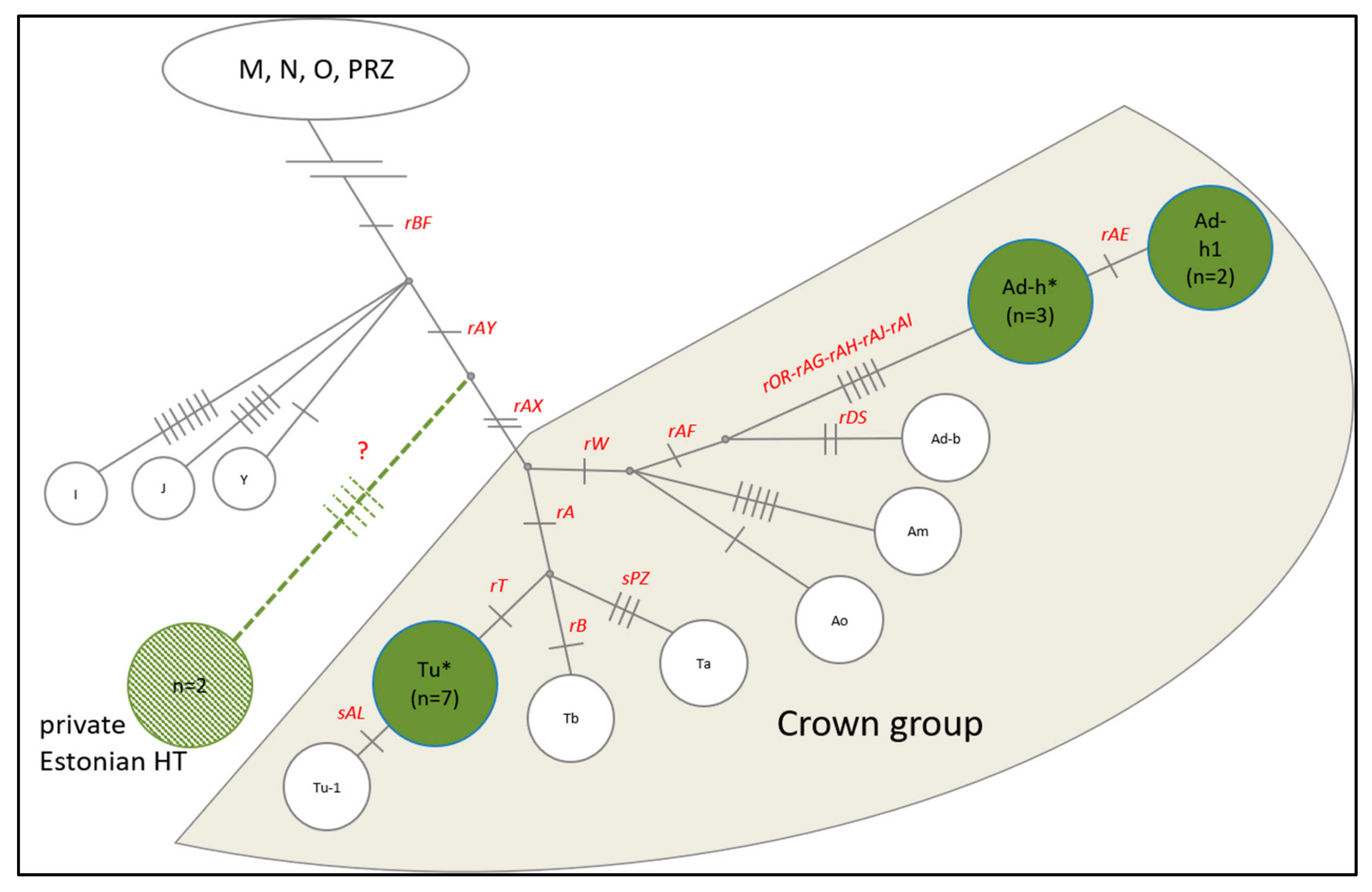
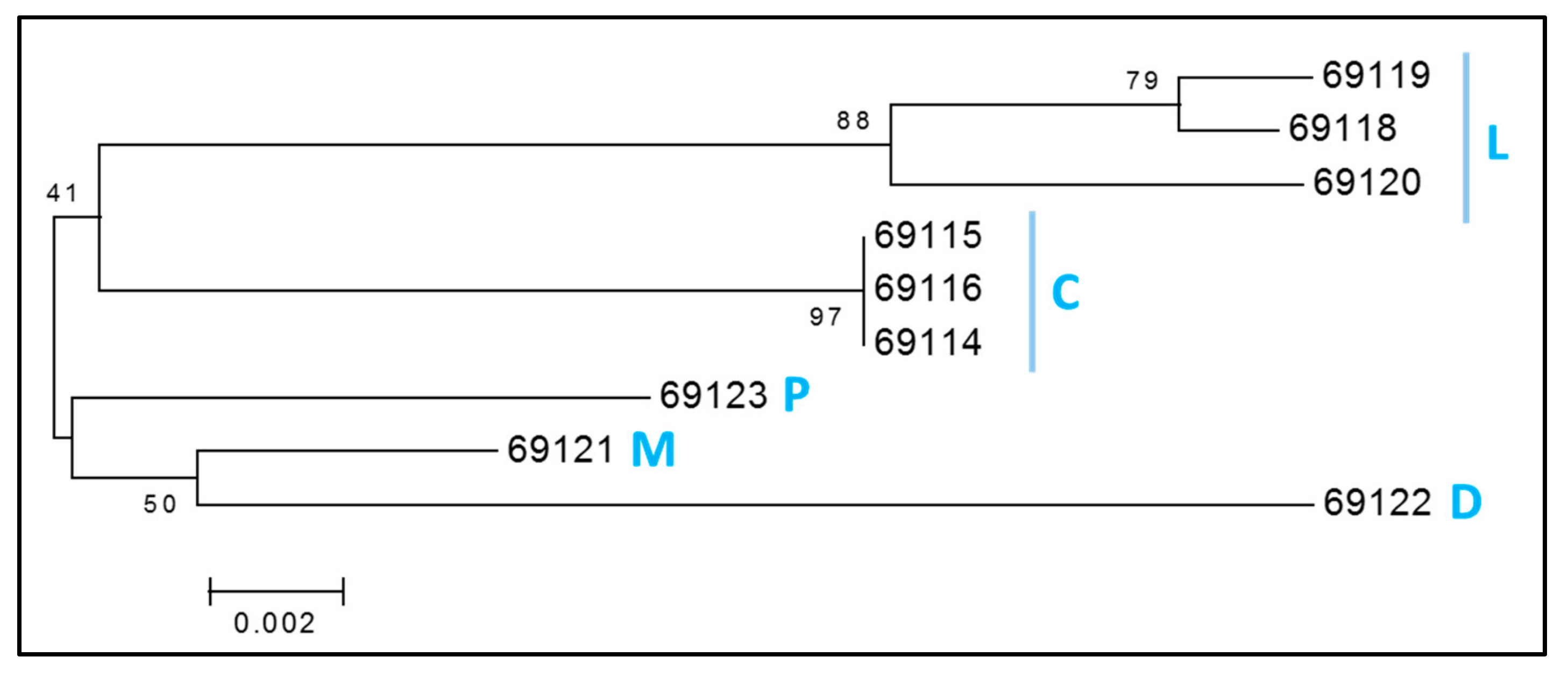
3.3. Y Chromosome and Mitochondrial Haplogroups
3.4. Genotyping for Gait Mutation in DMRT3
4. Discussion
5. Conclusions and Future Approaches
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maldre, L. Archeozoology. Estonian Native Horse. Horisont 1997, 6, 1–4. (In Estonian) [Google Scholar]
- Rannamäe, E. Zooarcheology of Equus caballus; Tartu University: Tartu, Estonia, 2008. (In Estonian) [Google Scholar]
- Lõugas, L. Investigating the history of the Estonian Native Horse. Oma Hobu 2006, Spring, 28–36. (In Estonian) [Google Scholar]
- Rooda, J.-A. Origins and breeding of the Estonian Native Horse. Oma Hobu 2004, 1, 4–5. (In Estonian) [Google Scholar]
- EstHorse. Available online: http://esthorse.ee/index.php?id=mainpage (accessed on 23 May 2019).
- Pasicka, E. Polish Konik Horse Characteristics and historical background of native descendants of Tarpan. Acta Sci. Pol. Med. Vet. 2013, 12, 25–38. [Google Scholar]
- Librado, P.A.; Fages, C.; Gaunitz, M.; Leonardi, S.; Wagner, N.; Khan, K.; Hanghoj, S.A.; Alquraishi, A.H.; Alfarhan, K.A.; Al-Rasheid, C.D.; et al. The evolutionary origin and genetic makeup of domestic horses. Genetics 2016, 204, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Austen, C.S.; Gorrie, P.R.; Swinney, N.J. The Complete Illustrated Encyclopedia of Horses; Metro Books: New York, NY, USA, 2008; p. 351. [Google Scholar]
- Sild, E.; Rooni, S.; Värv, K.; Røed, R.; Popov, J.K.; Viinlass, H. Genetic diversity of Estonian horse breeds and their genetic affinity to northern European and some Asian breeds. Livest. Sci. 2019, 220, 57–66. [Google Scholar] [CrossRef]
- ARIB. Agricultural Registers and Information Board. Equine Database. Available online: https://ariel.pria.ee/hobu/%20 (accessed on 29 May 2019).
- FAO Food and Agriculture Organization of the United States. World Watch List for Animal Diversity; Scherf, B.D., Ed.; FAO: Rome, Italy, 2000. [Google Scholar]
- Randlaht, I. Estonian Native horse coat colors. In The Estonian Native Horse; Sarjas, A.R.A.A., Ed.; Estonian Native Horse Conservation Society: Tallinn, Estonia, 2011; pp. 33–41. (In Estonian) [Google Scholar]
- Raudsepp, T. A good horse has no color. In The Estonian Native Horse; Sarjas, A.R.A.A., Ed.; The Estonian Native Horse Conservation Society: Tallinn, Estonia, 2011; pp. 6–32. (In Estonian) [Google Scholar]
- Imsland, F.K.; McGowan, C.J.; Rubin, C.; Henegar, E.; Sundstrom, J.; Berglund, D.; Schwochow, U.; Gustafson, P.; Imsland, K.; Lindblad-Toh, G.; et al. Regulatory mutations in TBX3 disrupt asymmetric hair pigmentation that underlies Dun camouflage color in horses. Nat. Genet. 2016, 48, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Khanshour, M.A.; Juras, R.; Cothran, E.G. Microsatellite analysis of genetic variability in Waler horses from Australia. Aust. J. Zool. 2013, 61, 357–365. [Google Scholar] [CrossRef]
- Juras, R.; Cothran, E.G.; Klimas, R. Genetic Analysis of Three Lithuanian Native Horse Breeds. Acta Agric. Scand. Sect. A Anim. Sci. 2003, 53, 180–185. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Reynolds, J.W.B.S.; Cockerham, C.C. Estimation of the coancestry coefficient basis for a short-term genetic Distance. Genetics 1983, 105, 767–779. [Google Scholar] [PubMed]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package) version 3.6; Distributed by the author; Department of Genome Sciences, University of Washington: Seattle, WA, USA, 2004. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis, Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Belkhir, K.P.B.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX, version 4.05, Logiciel sous Windows TM pour la génétique des Populations; Institut des Sciences de l’Evolution, University of Montpellier II: Montpellier, France, 1996–2004.
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Ingerence of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar]
- Hovens, H.J.P.M.; Rijkers, T.A.J.M. The origins of the Exmoor pony: Did the wild horse survive in Britain? Lutra 2014, 56, 129–136. [Google Scholar]
- Juras, R.; Cothran, E.G. Genetic diversity of the Zemaitukai horse. EAAP 2004, 116, 177–183. [Google Scholar]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2003, 4, 137–138. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Wallner, B.N.; Palmieri, C.; Vogl, D.; Rigler, E.; Bozlak, T.; Druml, V.; Jagannathan, T.; Leeb, R.; Fries, J.; Tetens, G.; et al. Y Chromosome Uncovers the Recent Oriental Origin of Modern Stallions. Curr. Biol. 2017, 27, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Felkel, S.C.; Vogl, D.; Rigler, V.; Jagannathan, T.; Leeb, R.; Fries, M.; Neuditschko, S.; Rieder, B.; Velie, G.; Lindgren, C.-J.; et al. Asian horses deepen the MSY phylogeny. Anim. Genet. 2018, 49, 90–93. [Google Scholar] [CrossRef]
- Cothran, E.; Juras, R.; Macijauskiene, V. Mitochondrial DNA D-loop sequence variation among 5 maternal lines of the Zemaitukai horse breed. Genet. Mol. Biol 2005, 28, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Hooshiar Kashani, B.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Promerova, M.L.S.; Andersson, R.; Juras, M.C.; Penedo, M.; Reissmann, T.; Tozaki, R.; Bellone, S.; Dunner, P.; Horin, F.; Imsland, P.; et al. Worldwide frequency distribution of the ‘Gait keeper’ mutation in the DMRT3 gene. Anim. Genet. 2014, 45, 274–282. [Google Scholar] [CrossRef]
- Staiger, E.A.M.S.; Almen, M.; Promerova, S.; Brooks, E.G.; Cothran, F.; Imsland, K.; Jaderkvist Fegraeus, G.; Lindgren, H.; Mehrabani Yeganeh, S.; Mikko, J.L.; et al. The evolutionary history of the DMRT3 ‘Gait keeper’ haplotype. Anim. Genet. 2017, 48, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Andersson, L.S.M.; Larhammar, F.; Memic, H.; Wootz, D.; Schwochow, C.J.; Rubin, K.; Patra, T.; Arnason, L.; Wellbring, G.; Hjalm, F.; et al. Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice. Nature 2012, 488, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Zabek, T.A.; Zyga, A.R.; Slota, E. Analysis of genetic variation in Malopolski Horses using molecular and pedigree data. Ann. Anim. Sci. 2006, 6, 13–27. [Google Scholar]
- Mackowski, M.S.; Mucha, G.C.; Cieslak, J. Genetic diversity in Hucul and Polish primitive horse breeds. Arch. Anim. Breed. 2015, 58, 23–31. [Google Scholar] [CrossRef]
- Van de Goor, L.H.; van Haeringen, W.A.; Lenstra, J.A. Population studies of 17 equine STR for forensic and phylogenetic analysis. Anim. Genet. 2011, 42, 627–633. [Google Scholar] [CrossRef]
- Andersson, L. Analysis of Inbreeding in the Swedish Gotland Pony using pedigree information and microsatellite markers. In Animal Breeding and Genetics; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2010; p. 37. [Google Scholar]
- Aberle, K.S.H.; Hamann, C.D.; Distl, O. Genetic diversity in German draught horse breeds compared with a group of primitive, riding and wild horses by means of microsatellite DNA markers. Anim. Genet. 2004, 35, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.L.; Mickelson, J.R.; Cothran, E.G.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; Brama, P.; et al. Genetic diversity in the modern horse illustrated from genome-wide SNP data. PLoS ONE 2013, 8, e54997. [Google Scholar] [CrossRef] [PubMed]
- McCue, M.E.; Bannasch, D.L.; Petersen, J.L.; Gurr, J.; Bailey, E.; Binns, M.M.; Distl, O.; Guérin, G.; Hasegawa, T.; Hill, E.W.; et al. A high density SNP array for the domestic horse and extant Perissodactyla: Utility for association mapping, genetic diversity, and phylogeny studies. PLoS Genet. 2012, 8, e1002451. [Google Scholar] [CrossRef]
- Orlando, L.; Ginolhac, A.; Zhang, G.; Froese, D.; Albrechtsen, A.; Stiller, M.; Schubert, M.; Cappellini, E.; Petersen, B.; Moltke, I.; et al. Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse. Nature 2013, 499, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Bjørnstad, G.; Nilsen, N.Ø.; Røed, K.H. Genetic relationship between Mongolian and Norwegian horses? Anim. Genet. 2003, 34, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Der Sarkissian, C.; Ermini, L.; Schubert, M.; Jónsson, H.; Albrechtsen, A.; Fumagalli, M.; Yang, M.A.; Gamba, C.; Seguin-Orlando, A.; et al. Tracking the origins of Yakutian horses and the genetic basis for their fast adaptation to subarctic environments. Proc. Natl. Acad. Sci. USA 2015, 112, E6889–E6897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaunitz, C.; Fages, A.; Hanghøj, K.; Albrechtsen, A.; Khan, N.; Schubert, M.; Seguin-Orlando, A.; Owens, I.J.; Felkel, S.; Bignon-Lau, O.; et al. Ancient genomes revisit the ancestry of domestic and Przewalski’s horses. Science 2018, 360, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Der Sarkissian, C.; Balanovsky, O.; Brandt, G.; Khartanovich, V.; Buzhilova, A.; Koshel, S.; Zaporozhchenko, V.; Gronenborn, D.; Moiseyev, V.; Kolpakov, E.; et al. Ancient DNA reveals prehistoric gene-flow from siberia in the complex human population history of North East Europe. PLoS Genet. 2013, 9, e1003296. [Google Scholar] [CrossRef] [PubMed]
- Rootsi, S.; Zhivotovsky, L.A.; Baldovič, M.; Kayser, M.; Kutuev, I.A.; Khusainova, R.; Bermisheva, M.A.; Gubina, M.; Fedorova, S.A.; Ilumäe, A.M.; et al. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe. Eur. J. Hum. Genet. 2007, 15, 204–211. [Google Scholar] [CrossRef]
- Gurgul, A.; Jasielczuk, I.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Stefaniuk-Szmukier, M.; Szmatoła, T.; Polak, G.; Tomczyk-Wrona, I.; Bugno-Poniewierska, M. A genome-wide scan for diversifying selection signatures in selected horse breeds. PLoS ONE 2019, 14, e0210751. [Google Scholar] [CrossRef] [PubMed]
- Szwaczkowski, T.; Greguła-Kania, M.; Stachurska, A.; Borowska, A.; Jaworski, Z.; Gruszecki, T.M. Inter- and intra-genetic diversity in the Polish Konik horse: Implications for the conservation program. Can. J. Anim. Sci. 2016, 96, 570–580. [Google Scholar] [CrossRef]
- Warmuth, V.; Eriksson, A.; Bower, M.A.; Cañon, J.; Cothran, G.; Distl, O.; Glowatzki-Mullis, M.L.; Hunt, H.; Luís, C.; do Mar Oom, M.; et al. European domestic horses originated in two holocene refugia. PLoS ONE 2011, 6, e18194. [Google Scholar] [CrossRef]
- Fages, A.; Hanghøj, K.; Khan, N.; Gaunitz, C.; Seguin-Orlando, A.; Leonardi, M.; Constantz, C.M.; Gamba, C.; Al-Rasheid, K.A.; Albizuri, S.; et al. Tracking Five Millennia of Horse Management with Extensive Ancient Genome Time Series. Cell 2019, 177, 1419–1435. [Google Scholar] [CrossRef] [PubMed]
- Felkel, S.; Vogl, C.; Rigler, D.; Dobretsberger, V.; Chowdhary, B.P.; Distl, O.; Fries, R.; Jagannathan, V.; Janečka, J.E.; Leeb, T.; et al. The horse Y chromosome as an informative marker for tracing sire lines. Sci. Rep. 2019, 9, 6095. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Vogl, C.; Shukla, P.; Burgstaller, J.P.; Druml, T.; Brem, G. Identification of genetic variation on the horse y chromosome and the tracing of male founder lineages in modern breeds. PLoS ONE 2013, 8, e60015. [Google Scholar] [CrossRef] [PubMed]
- Wutke, S.; Sandoval-Castellanos, E.; Benecke, N.; Döhle, H.J.; Friederich, S.; Gonzalez, J.; Hofreiter, M.; Lõugas, L.; Magnell, O.; Malaspinas, A.S.; et al. Decline of genetic diversity in ancient domestic stallions in Europe. Sci. Adv. 2018, 4, eaap9691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippold, S.; Knapp, M.; Kuznetsova, T.; Leonard, J.A.; Benecke, N.; Ludwig, A.; Rasmussen, M.; Cooper, A.; Weinstock, J.; Willerslev, E.; et al. Discovery of lost diversity of paternal horse lineages using ancient DNA. Nat. Commun. 2011, 2, 450. [Google Scholar] [CrossRef]
- Vilà, C.; Leonard, J.A.; Götherström, A.; Marklund, S.; Sandberg, K.; Lidén, K.; Wayne, R.K.; Ellegren, H. Widespread Origins of Domestic Horse Lineages. Science 2001, 291, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Gamba, C.; Gaunitz, C.; Der Sarkissian, C.; Pruvost, M.; Albrechtsen, A.; Fages, A.; Khan, N.; Schubert, M.; Jagannathan, V. Ancient genomic changes associated with domestication of the horse. Science 2017, 356, 442–445. [Google Scholar] [CrossRef]
- Kusza, S.; Priskin, K.; Ivankovic, A.; Jedrzejewska, B.; Podgorski, T.; Jávor, A.; Mihók, S. Genetic characterization and population bottleneck in the Hucul horse based on microsatellite and mitochondrial data. Biol. J. Linn. Soc. 2013, 109, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.S.; et al. Coat color variation at the beginning of horse domestication. Science 2009, 324, 485. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk-Szmukier, M.; Ropka-Molik, K.; Piórkowska, K.; Szmatoła, T.; Długosz, B.; Pisarczyk, W.; Bugno-Poniewierska, M. Variation in TBX3 Gene Region in Dun Coat Color Polish Konik Horses. J. Equine Vet. Sci. 2017, 49, 60–62. [Google Scholar] [CrossRef]
- Mackowski, M.; Wodas, L.; Brooks, S.A.; Cieslak, J. TBX3 and ASIP genotypes reveal discrepancies in officially recorded coat colors of Hucul horses. Animal 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population (Abbreviation) | N | MNA (SE) | Ae (SE) | HO (SE) | HE (SE) | FIS (SE) |
|---|---|---|---|---|---|---|
| Estonian Native Horse (ENH) | 33 | 6.600 (0.423) | 3.983 (0.309) | 0.766 (0.033) | 0.723 (0.025) | −0.059 (0.023) |
| Hucul (HU) | 45 | 6.933 (0.441) | 4.033 (0.418) | 0.732 (0.047) | 0.709 (0.037) | −0.018 (0.031) |
| Konik/Polish Primitive (PO) | 53 | 7.133 (0.446) | 3.820 (0.280) | 0.696 (0.039) | 0.703 (0.038) | 0.006 (0.019) |
| Finn Horse (FH) | 25 | 6.400 (0.567) | 4.160 (0.367) | 0.717 (0.056) | 0.705 (0.053) | −0.019 (0.023) |
| Gotland Pony (GT) | 43 | 5.533 (0.322) | 3.024 (0.216) | 0.650 (0.03) | 0.644 (0.026) | −0.009 (0.024) |
| Icelandic Horse (IC) | 38 | 6.067 (0.384) | 3.526 (0.260) | 0.711 (0.036) | 0.686 (0.032) | −0.037 (0.024) |
| Norwegian Fjord (FJ) | 50 | 5.867 (0.496) | 3.355 (0.232) | 0.694 (0.049) | 0.665 (0.043) | −0.042 (0.019) |
| Shetland Pony (SP) | 66 | 6.933 (0.358) | 3.637 (0.249) | 0.649 (0.037) | 0.703 (0.024) | 0.083 (0.032) |
| Exmoor Pony (EX) | 70 | 5.133 (0.413) | 2.984 (0.221) | 0.619 (0.038) | 0.635 (0.030) | 0.035 (0.026) |
| Breed | N | Wild-Type (C/C) | Carrier (C/A) | Mutant (A/A) |
|---|---|---|---|---|
| Estonian Native Horse | 33 | 33 | - | - |
| Hucul Pony | 52 | 50 | 2 | - |
| Exmoor Pony | 29 | 29 | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castaneda, C.; Juras, R.; Khanshour, A.; Randlaht, I.; Wallner, B.; Rigler, D.; Lindgren, G.; Raudsepp, T.; Cothran, E.G. Population Genetic Analysis of the Estonian Native Horse Suggests Diverse and Distinct Genetics, Ancient Origin and Contribution from Unique Patrilines. Genes 2019, 10, 629. https://doi.org/10.3390/genes10080629
Castaneda C, Juras R, Khanshour A, Randlaht I, Wallner B, Rigler D, Lindgren G, Raudsepp T, Cothran EG. Population Genetic Analysis of the Estonian Native Horse Suggests Diverse and Distinct Genetics, Ancient Origin and Contribution from Unique Patrilines. Genes. 2019; 10(8):629. https://doi.org/10.3390/genes10080629
Chicago/Turabian StyleCastaneda, Caitlin, Rytis Juras, Anas Khanshour, Ingrid Randlaht, Barbara Wallner, Doris Rigler, Gabriella Lindgren, Terje Raudsepp, and E. Gus Cothran. 2019. "Population Genetic Analysis of the Estonian Native Horse Suggests Diverse and Distinct Genetics, Ancient Origin and Contribution from Unique Patrilines" Genes 10, no. 8: 629. https://doi.org/10.3390/genes10080629