Global Investigation of Cytochrome P450 Genes in the Chicken Genome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animals
2.3. Identification of P450 Members
2.4. Gene Characteristics and Sequence Analyses
2.5. RNA-seq Library Construction and Sequencing
2.6. Sequencing Data Processing
2.7. Quantitative Real-Time PCR (qRT-PCR)
3. Results
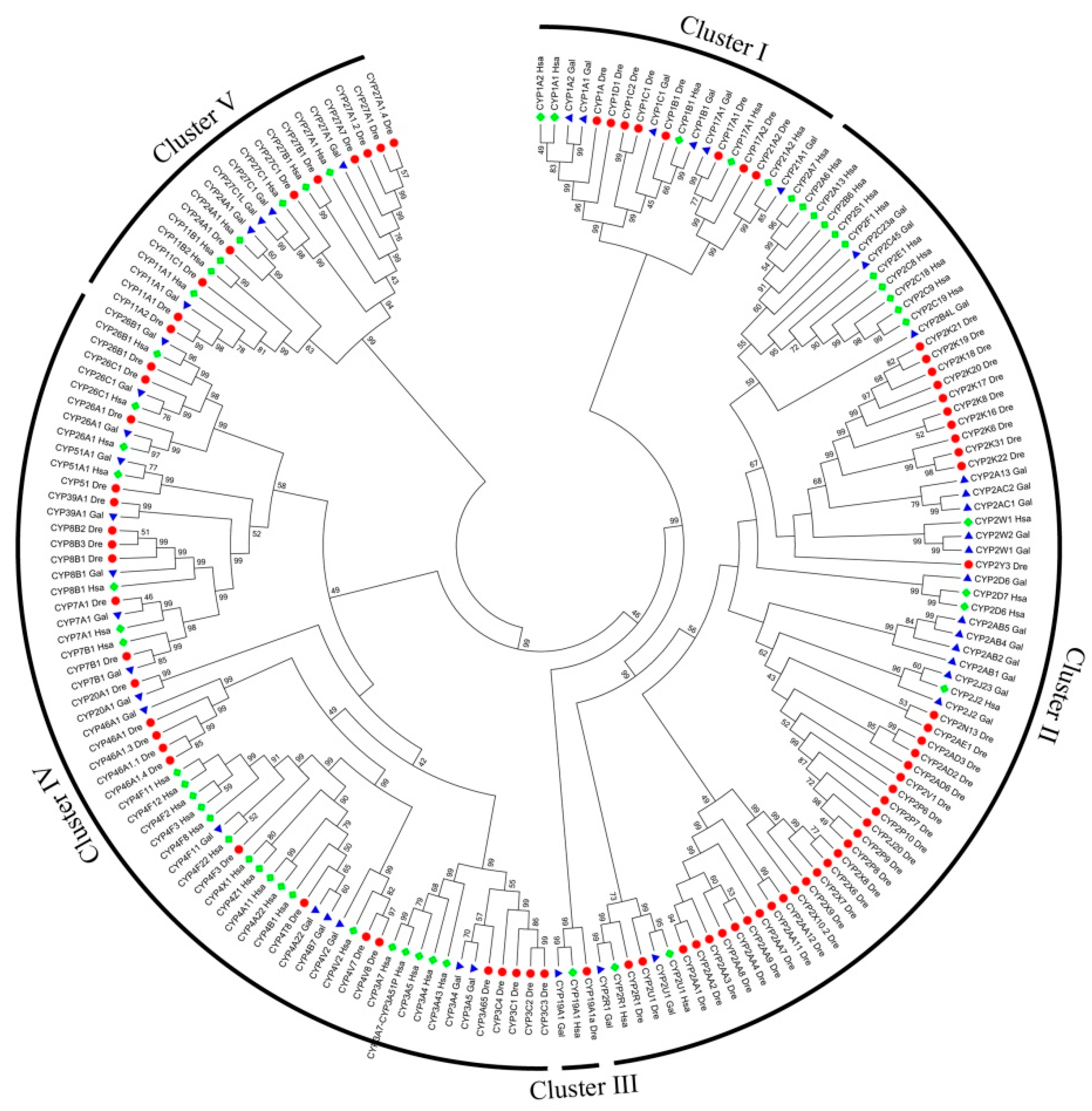
3.1. CYP Identification and Phylogenetic Analysis
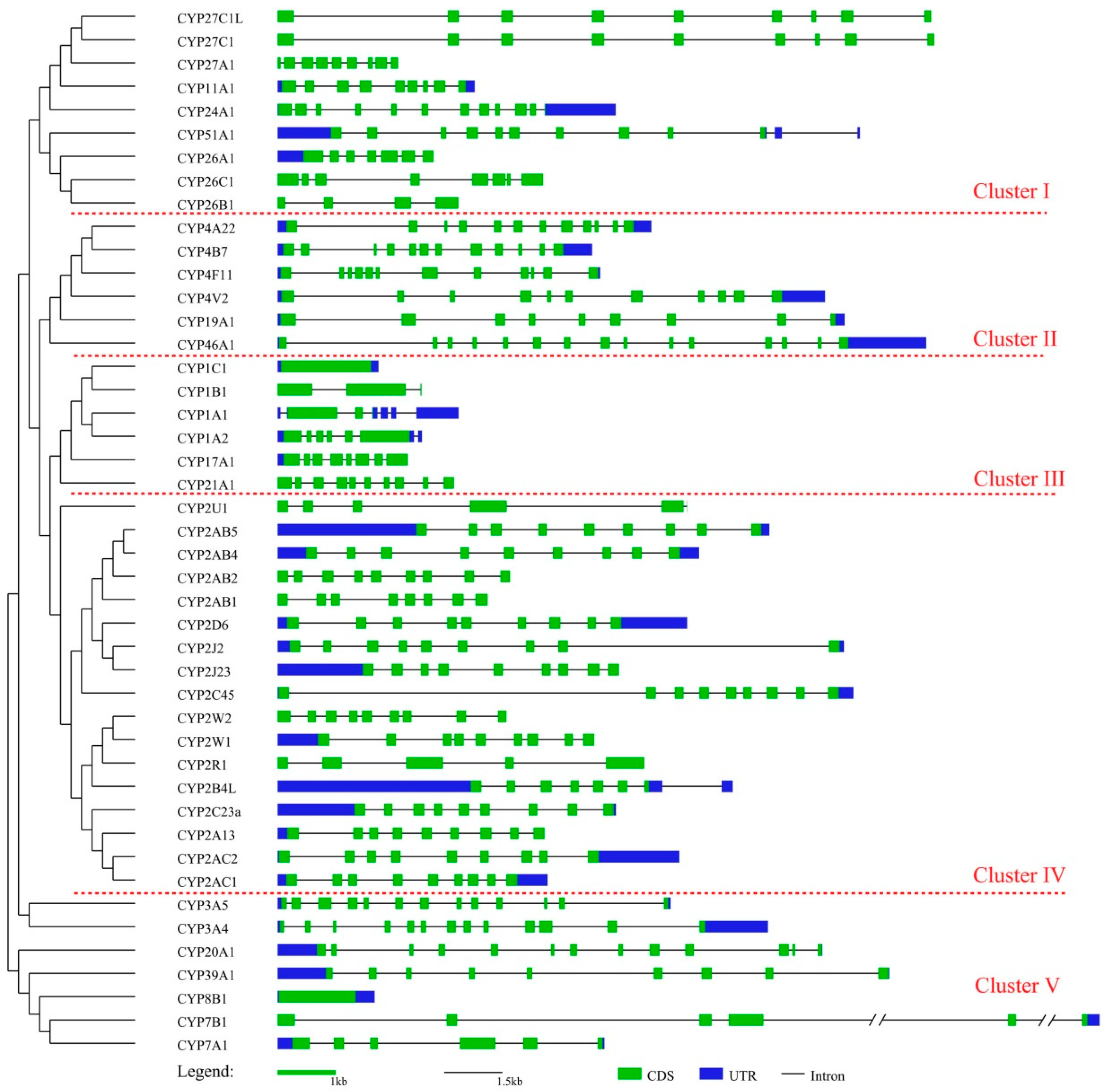
3.2. Phylogenetic and Gene Characteristic Analyses of cCYPs
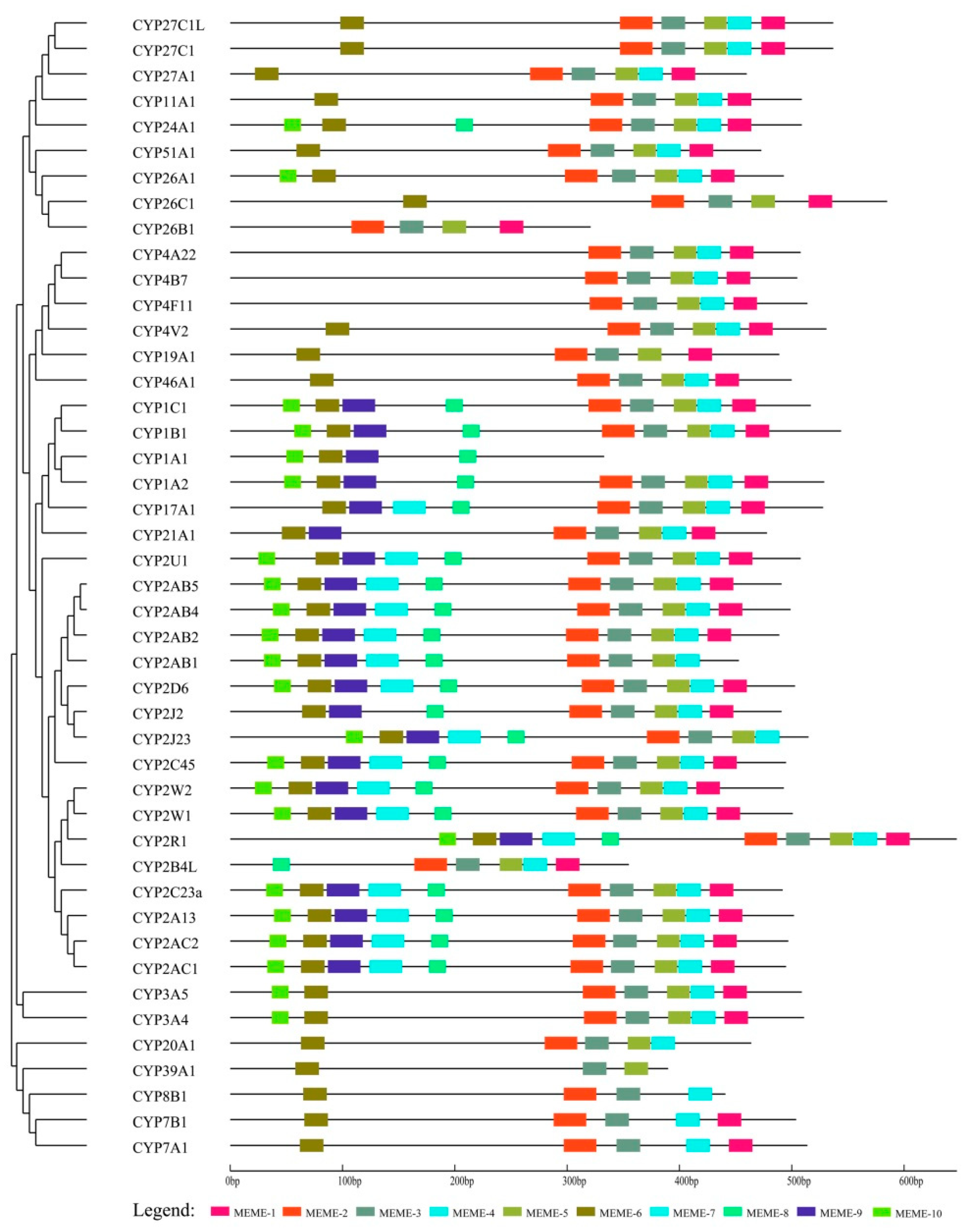
3.3. Conserved Motif Analysis of cCYPs
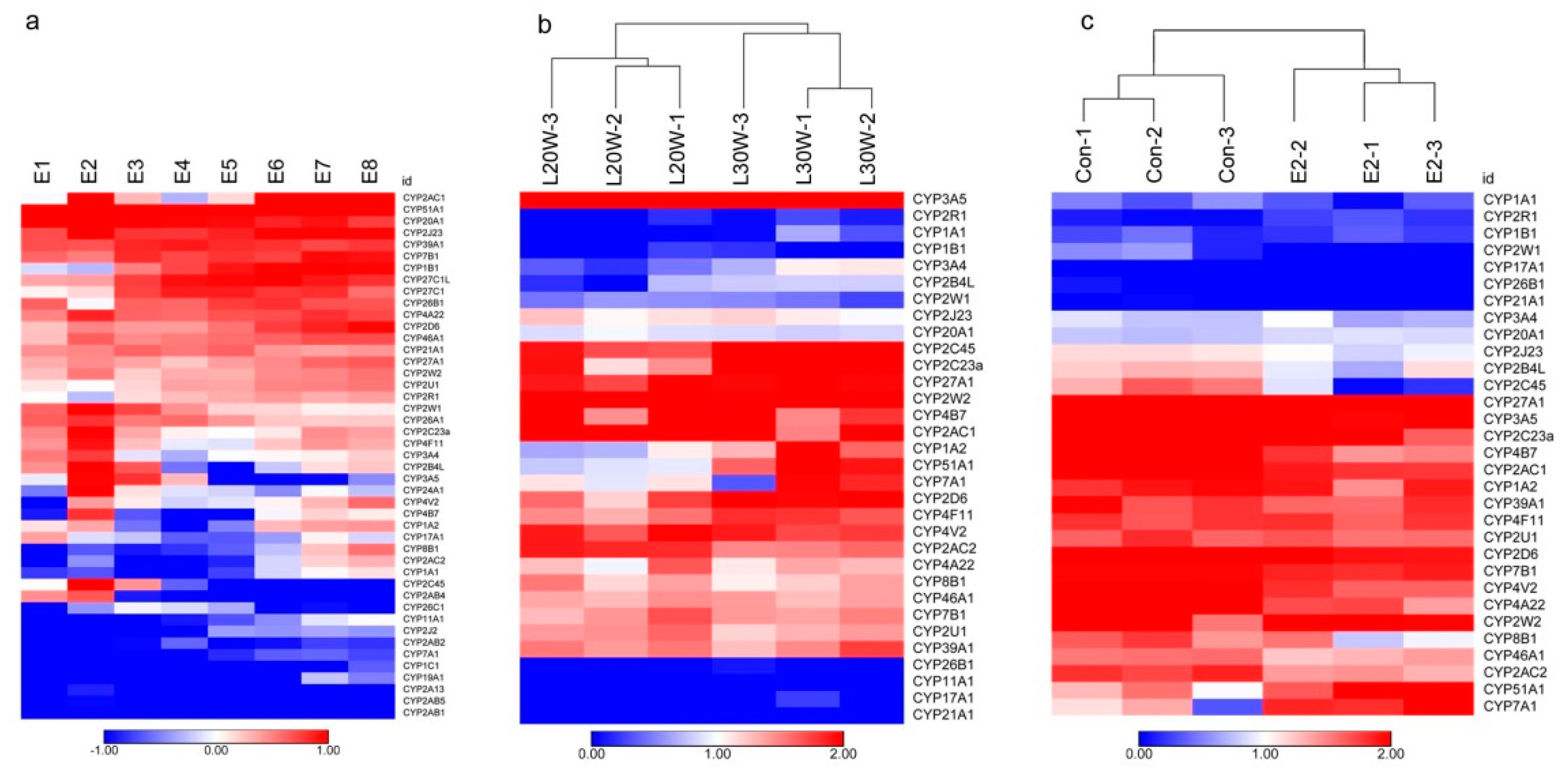
3.4. Expression Profile of cCYPs in Developing Chicken Embryo
3.5. Expression Profile of P450 Members in Chicken Liver
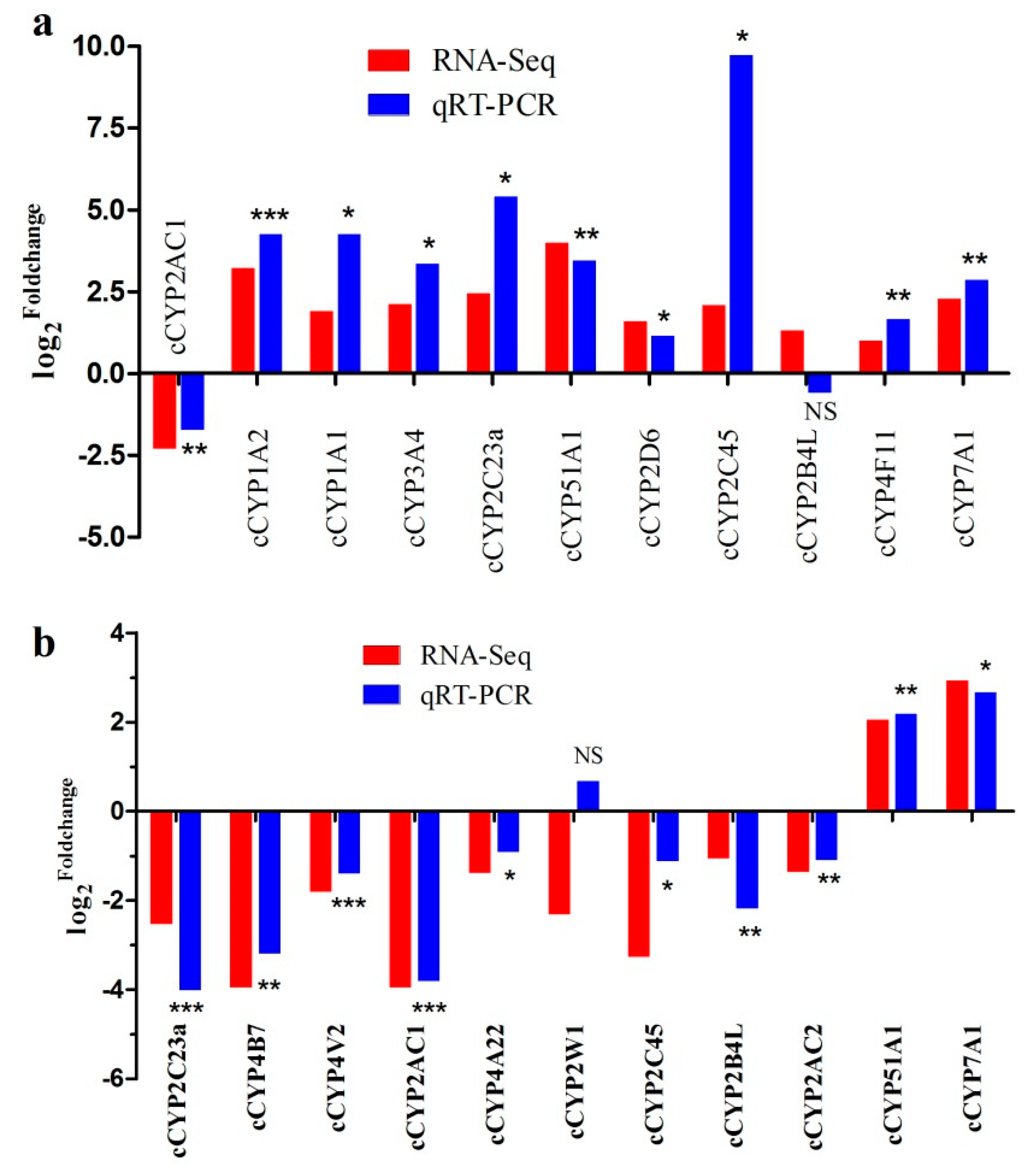
3.6. Identification of Estrogen-Responsive cCYPs
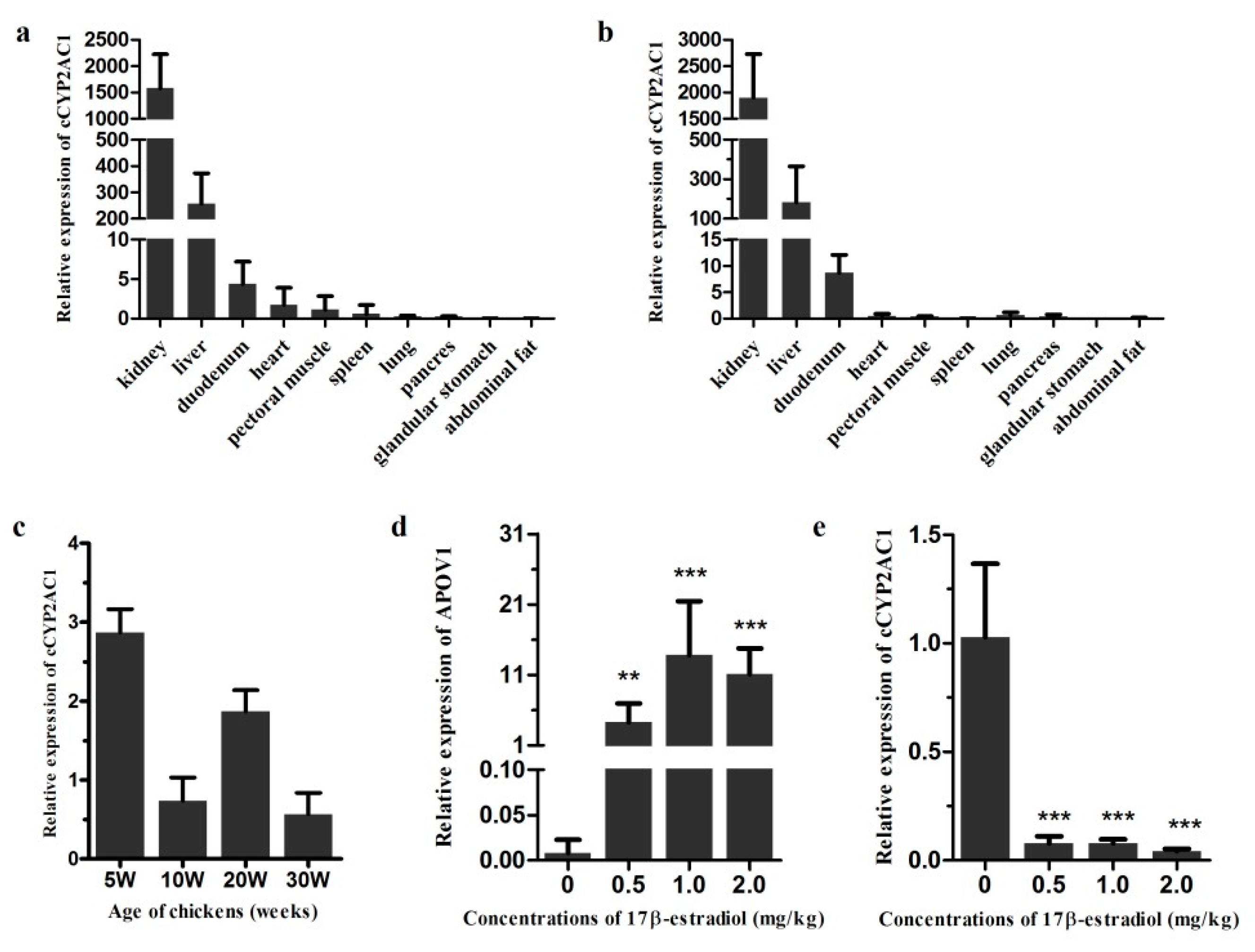
3.7. Expression Characteristics of cCYP2AC1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nelson, D.R. Plant cytochrome P450s from moss to poplar. Phytochem. Rev. 2006, 5, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Progress in tracing the evolutionary paths of cytochrome P450. Biochim. Biophys. Acta 2011, 1814, 14–18. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. The cytochrome p450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar]
- Nelson, D.R. Comparison of P450s from human and fugu: 420 million years of vertebrate P450 evolution. Arch. Biochem. Biophys. 2003, 409, 18–24. [Google Scholar] [CrossRef]
- Nelson, D.R.; Ming, R.; Alam, M.; Schuler, M.A. Comparison of Cytochrome P450 Genes from Six Plant Genomes. Trop. Plant Biol. 2008, 1, 216–235. [Google Scholar] [CrossRef]
- Visser, M.; Weber, K.; Rincon, G.; Merritt, D.A. Use of RNA-seq to determine variation in canine cytochrome P450 mRNA expression between blood, liver, lung, kidney and duodenum in healthy beagles. J. Vet. Pharmacol. Ther. 2017, 40, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.P.; Kawai, Y.K.; Ikenaka, Y.; Kawata, M.; Ikushiro, S.; Sakaki, T.; Ishizuka, M. Avian Cytochrome P450 (CYP) 1–3 Family Genes: Isoforms, Evolutionary Relationships, and mRNA Expression in Chicken Liver. PLoS ONE 2013, 8, e75689. [Google Scholar] [CrossRef]
- Rasmussen, M.K.; Theil, P.K.; Oksbjerg, N. Constitutive expression of cytochrome P450 in foetal and adult porcine livers—Effects of body weight. Toxicol. Lett. 2016, 258, 87–92. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Rasmussen, M.K.; Bertholdt, L.; Gudiksen, A.; Pilegaard, H.; Knudsen, J.G. Impact of fasting followed by short-term exposure to interleukin-6 on cytochrome P450 mRNA in mice. Toxicol. Lett. 2018, 282, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Konkel, A.; Schunck, W. Role of cytochrome P450 enzymes in the bioactivation of polyunsaturated fatty acids. Biochim. Biophys. Acta 2011, 1814, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Bistolas, N.; Wollenberger, U.; Jung, C.; Scheller, F.W. Cytochrome P450 biosensors: A review. Biosensors Bioelectron. 2005, 20, 2408–2423. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Cai, M.X.; Thomas, P.E.; Conney, A.H.; Zhu, B.T. Characterization of the Oxidative Metabolites of 17β-Estradiol and Estrone Formed by 15 Selectively Expressed Human Cytochrome P450 Isoforms. Endocrinology 2003, 144, 3382–3398. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Kisselev, P.; Schunck, W.; Roots, I. Inhibition of 17β-estradiol activation by CYP1A1: Genotype- and regioselective inhibition by St. John’s Wort and several natural polyphenols. Biochim. Biophys. Acta 2011, 1814, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Go, R.; Hwang, K.; Kim, C.; Byun, Y.; Nam, K.; Choi, K. Effect of dioxin and 17β-estradiol on the expression of cytochrome P450 1A1 gene via an estrogen receptor dependent pathway in cellular and xenografted models. Environ. Toxicol. 2017, 32, 2225–2233. [Google Scholar] [CrossRef]
- Almeida, D.; Maldonado, E.; Khan, I.; Silva, L.; Gilbert, M.T.P.; Zhang, G.; Jarvis, E.D.; Brien, S.J.O.; Johnson, W.E.; Antunes, A. Whole-Genome Identification, Phylogeny, and Evolution of the Cytochrome P450 Family 2 (CYP2) Subfamilies in Birds. Genome Biol. Evol. 2016, 8, 1115–1131. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Kawai, Y.K.; Watanabe, K.P.; Ishii, A.; Ohnuma, A.; Sawa, H.; Ikenaka, Y.; Ishizuka, M. De novo sequence analysis of cytochrome P450 1–3 genes expressed in ostrich liver with highest expression of CYP2G19. Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 201–208. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Li, H.; Wang, T.; Xu, C.; Wang, D.; Ren, J.; Li, Y.; Tian, Y.; Wang, Y.; Jiao, Y.; Kang, X. Transcriptome profile of liver at different physiological stages reveals potential mode for lipid metabolism in laying hens. BMC Genom. 2015, 16, 763. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, 1–13. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Paolini, M.; Pozzetti, L.; Sapone, A.; Biagi, G.L.; Cantelliforti, G. Development of basal and induced testosterone hydroxylase activity in the chicken embryo in ovo. Br. J. Pharmacol. 1997, 122, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Dawling, S.; Roodi, N.; Parl, F.F. Methoxyestrogens Exert Feedback Inhibition on Cytochrome P450 1A1 and 1B1. Cancer Res. 2003, 63, 3127–3132. [Google Scholar]
- Berkowitz, E.A.; Chu, W.W.; Evans, M.I. Insulin inhibits the estrogen-dependent expression of the chicken very low density apolipoprotein II gene in Leghorn male hepatoma cells. Mol. Endocrinol. 1993, 7, 507–514. [Google Scholar] [PubMed]
- Wijnholds, J.; Muller, E.; Ab, G. Oestrogen facilitates the binding of ubiquitous and liver-enriched nuclear proteins to the apoVLDL II promoter in vivo. Nucleic Acids Res. 1991, 19, 33–41. [Google Scholar] [CrossRef]
- Wijnholds, J.; Philipsen, J.N.J.; Ab, G. Tissue-specific and steroid-dependent interaction of transcription factors with the oestrogen-inducible apoVLDL II promoter in vivo. EMBO J. 1988, 7, 2757–2763. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Roussel, R.R.; Roberts, M.; Marek, D.; Wood, S.G.; Walton, H.S.; Dwyer, B.E.; Sinclair, P.R.; Sinclair, J.F. Effect of Arsenite on Induction of CYP1A and CYP2H in Primary Cultures of Chick Hepatocytes. Toxicol. Appl. Pharmacol. 1998, 150, 376–382. [Google Scholar] [CrossRef]
- Hamilton, J.W.; Bement, W.J.; Sinclair, P.R.; Sinclair, J.F.; Alcedo, J.A.; Wetterhahn, K.E. Inhibition of protein synthesis increases the transcription of the phenobarbital-inducible CYP2H1 and CYP2H2 genes in chick embryos hepatocytes. Arch. Biochem. Biophys. 1992, 298, 96–104. [Google Scholar] [CrossRef]
- Louis, C.A.; Wood, S.G.; Walton, H.S.; Sinclair, P.R.; Sinclair, J.F. Mechanism of the synergistic induction of CYP2H by isopentanol plus ethanol: Comparison to glutethimide and relation to induction of 5-aminolevulinate synthase. Arch. Biochem. Biophys. 1998, 360, 239–247. [Google Scholar] [CrossRef]
- Hu, S.X. Effect of age on hepatic cytochrome P450 of Ross 708 broiler chickens. Poult. Sci. 2013, 92, 1283–1291. [Google Scholar] [CrossRef]
- Kubota, A.; Stegeman, J.J.; Goldstone, J.V.; Nelson, D.R.; Kim, E.; Tanabe, S.; Iwata, H. Cytochrome P450 CYP2 genes in the common cormorant: Evolutionary relationships with 130 diapsid CYP2 clan sequences and chemical effects on their expression. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 280–289. [Google Scholar]
- Jacobs, E.T.; Van Pelt, C.; Forster, R.; Zaidi, W.; Hibler, E.A.; Galligan, M.A.; Haussler, M.R.; Jurutka, P.W. CYP24A1 and CYP27B1 Polymorphisms Modulate Vitamin D Metabolism in Colon Cancer Cells. Cancer Res. 2013, 73, 2563–2573. [Google Scholar] [CrossRef] [Green Version]
- Dilworth, F.J.; Scott, I.; Green, A.; Strugnell, S.; Guo, Y.; Roberts, E.A.; Kremer, R.; Calverley, M.J.; Makin, H.L.J.; Jones, G. Different Mechanisms of Hydroxylation Site Selection by Liver and Kidney Cytochrome P450 Species (CYP27 and CYP24) Involved in Vitamin D Metabolism. J. Biol. Chem. 1995, 270, 16766–16774. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.H.; O’Loughlin, P.D.; May, B.K.; Morris, H.A. Quantification of mRNA for the vitamin D metabolizing enzymes CYP27B1 and CYP24 and vitamin D receptor in kidney using real-time reverse transcriptase- polymerase chain reaction. J. Mol. Endocrinol. 2003, 31, 123. [Google Scholar] [CrossRef]
- Dong, L.M.; Ulrich, C.M.; Hsu, L.; Duggan, D.; Benitez, D.S.; White, E.; Slattery, M.L.; Farin, F.M.; Makar, K.W.; Carlson, C.S. Vitamin D Related Genes, CYP24A1 and CYP27B1, and Colon Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2540–2548. [Google Scholar] [CrossRef] [Green Version]
- Zanger, U.M.; Kagawa, N.; Lund, J.; Waterman, M.R. Distinct biochemical mechanisms for cAMP-dependent transcription of CYP17 and CYP21. FASEB J. 1992, 6, 719–723. [Google Scholar] [CrossRef]
- Nakajin, S.; Hall, P.F. Microsomal cytochrome P-450 from neonatal pig testis. Purification and properties of A C21 steroid side-chain cleavage system (17 alpha-hydroxylase-C17,20 lyase). J. Biol. Chem. 1981, 256, 3871–3876. [Google Scholar]
- Hanukoglu, I. Steroidogenic enzymes: Structure, function, and role in regulation of steroid hormone biosynthesis. J. Steroid Biochem. Mol. Biol. 1992, 43, 779–804. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.S.; Chern, H.D.; Chang, K.J.; Cheng, C.W.; Hsu, S.M.; Shen, C.Y. Breast cancer risk associated with genotype polymorphism of the estrogen-metabolizing genes CYP17, CYP1A1, and COMT: A multigenic study on cancer susceptibility. Cancer Res. 1999, 59, 4870–4875. [Google Scholar]
- Picado-Leonard, J.; Miller, W.L. Cloning and sequence of the human gene for P450c17 (steroid 17 alpha-hydroxylase/17,20 lyase): Similarity with the gene for P450c21. DNA 1987, 6, 439–448. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Takahashi, Y.; Egawa, C.; Noguchi, S. Breast Cancer Risk Associated with CYP1A1 Genetic Polymorphisms in Japanese Women. Breast J. 2002, 8, 209–215. [Google Scholar] [CrossRef]
- Taioli, E.; Trachman, J.; Chen, X.; Toniolo, P.; Garte, S. A CYP1A1 Restriction Fragment Length Polymorphism Is Associated with Breast Cancer in African-American Women. Cancer Res. 1995, 55, 3757–3758. [Google Scholar]
- Zhu, B.T.; Conney, A.H. Functional role of estrogen metabolism in target cells: Review and perspectives. Carcinogenesis 1998, 19, 1–27. [Google Scholar] [CrossRef]
- Chen, Z.; Hurh, Y.; Na, H.; Kim, J.; Chun, Y.; Kim, D.; Kang, K.; Cho, M.; Surh, Y. Resveratrol inhibits TCDD-induced expression of CYP1A1 and CYP1B1 and catechol estrogen-mediated oxidative DNA damage in cultured human mammary epithelial cells. Carcinogenesis 2004, 25, 2005–2013. [Google Scholar] [CrossRef]
- Wang, Y.; You, F.M.; Lazo, G.R.; Luo, M.; Thilmony, R.; Gordon, S.; Kianian, S.F.; Gu, Y.Q. PIECE: A database for plant gene structure comparison and evolution. Nucleic Acids Res. 2013, 41, 1159–1166. [Google Scholar] [CrossRef]
- Pirola, Y.; Rizzi, R.; Picardi, E.; Pesole, G.; Vedova, G.D.; Bonizzoni, P. PIntron: A fast method for detecting the gene structure due to alternative splicing via maximal pairings of a pattern and a text. BMC Bioinform. 2012, 13, 1–12. [Google Scholar] [CrossRef]
- Lin, Y.S.; Dowling, A.L.S.; Quigley, S.D.; Farin, F.M.; Zhang, J.; Lamba, J.K.; Schuetz, E.G.; Thummel, K.E. Co-regulation of CYP3A4 and CYP3A5 and contribution to hepatic and intestinal midazolam metabolism. Mol. Pharmacol. 2002, 62, 162–172. [Google Scholar] [CrossRef]
- Blevins, S.; Siegel, P.B.; Blodgett, D.J.; Ehrich, M.; Lewis, R.M. Liver enzymes in White Leghorns selected for the sheep red blood cell immune response. Poult. Sci. 2012, 91, 322–326. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Mean FPKM_20W | Mean FPKM_30W | Log2fold Change | FDR | UP/DOWN |
|---|---|---|---|---|---|
| CYP2AC1 | 454.764 | 92.029 | −2.305 | 3.32 × 10−3 | DOWN |
| CYP1A2 | 7.137 | 66.436 | 3.219 | 8.01 × 10−7 | UP |
| CYP1A1 | 0.687 | 2.576 | 1.907 | 2.45 × 10−4 | UP |
| CYP3A4 | 2.250 | 9.706 | 2.109 | 4.41 × 10−8 | UP |
| CYP2C23a | 41.463 | 226.648 | 2.451 | 1.48 × 10−5 | UP |
| CYP51A1 | 7.207 | 114.250 | 3.987 | 5.26 × 10−13 | UP |
| CYP2D6 | 37.170 | 112.042 | 1.592 | 1.31 × 10−4 | UP |
| CYP2C45 | 59.638 | 253.827 | 2.090 | 3.68 × 10−6 | UP |
| CYP2B4L | 2.537 | 6.291 | 1.310 | 1.87 × 10−2 | UP |
| CYP4F11 | 28.597 | 57.350 | 1.004 | 5.91 × 10−4 | UP |
| CYP7A1 | 11.291 | 55.082 | 2.286 | 2.11 × 10−2 | UP |
| Gene Name | Mean FPKM_Con | Mean FPKM_E2.0 | Log2fold Change | FDR | UP/DOWN |
|---|---|---|---|---|---|
| CYP2C23a | 502.082 | 86.824 | −2.53176 | 7.96 × 10−13 | DOWN |
| CYP4B7 | 618.130 | 39.801 | −3.95704 | 1.45 × 10−27 | DOWN |
| CYP4V2 | 172.665 | 49.238 | −1.81013 | 3.77 × 10−11 | DOWN |
| CYP2AC1 | 1027.453 | 66.303 | −3.95385 | 1.47 × 10−50 | DOWN |
| CYP4A22 | 110.614 | 42.371 | −1.38437 | 4.54 × 10−6 | DOWN |
| CYP2W1 | 2.976 | 0.594 | −2.32607 | 1.80 × 10−6 | DOWN |
| CYP2C45 | 33.088 | 3.414 | −3.27673 | 1.67 × 10−6 | DOWN |
| CYP2B4L | 18.566 | 8.897 | −1.06128 | 2.87 × 10−2 | DOWN |
| CYP2AC2 | 62.489 | 24.216 | −1.36764 | 5.24 × 10−9 | DOWN |
| CYP51A1 | 21.543 | 89.010 | 2.046759 | 2.18 × 10−5 | UP |
| CYP7A1 | 12.513 | 95.430 | 2.931051 | 1.72 × 10−5 | UP |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, J.; Yang, L.; Li, Q.; Zhang, Q.; Sun, C.; Liu, X.; Yang, N. Global Investigation of Cytochrome P450 Genes in the Chicken Genome. Genes 2019, 10, 617. https://doi.org/10.3390/genes10080617
Ren J, Yang L, Li Q, Zhang Q, Sun C, Liu X, Yang N. Global Investigation of Cytochrome P450 Genes in the Chicken Genome. Genes. 2019; 10(8):617. https://doi.org/10.3390/genes10080617
Chicago/Turabian StyleRen, Junxiao, Liyu Yang, Quanlin Li, Qinghe Zhang, Congjiao Sun, Xiaojun Liu, and Ning Yang. 2019. "Global Investigation of Cytochrome P450 Genes in the Chicken Genome" Genes 10, no. 8: 617. https://doi.org/10.3390/genes10080617