Genome-Wide Identification and Characterization of Olfactory Receptor Genes in Chinese Perch, Siniperca chuatsi


Abstract
:1. Introduction
2. Material and Methods
2.1. Genome-Wide Identification of OR Genes in the Genomes of Chinese Perch and Other Vertebrates
2.2. Classification of OR Genes
2.3. Phylogenetic Analysis of OR Genes
2.4. dN/dS Analysis
2.5. Analysis of Protein-Conserved Motifs
2.6. Detection of Chinese Perch-Specific Gained and/or Lost OR Genes
2.7. Expression Profile Analysis of OR Genes
3. Results
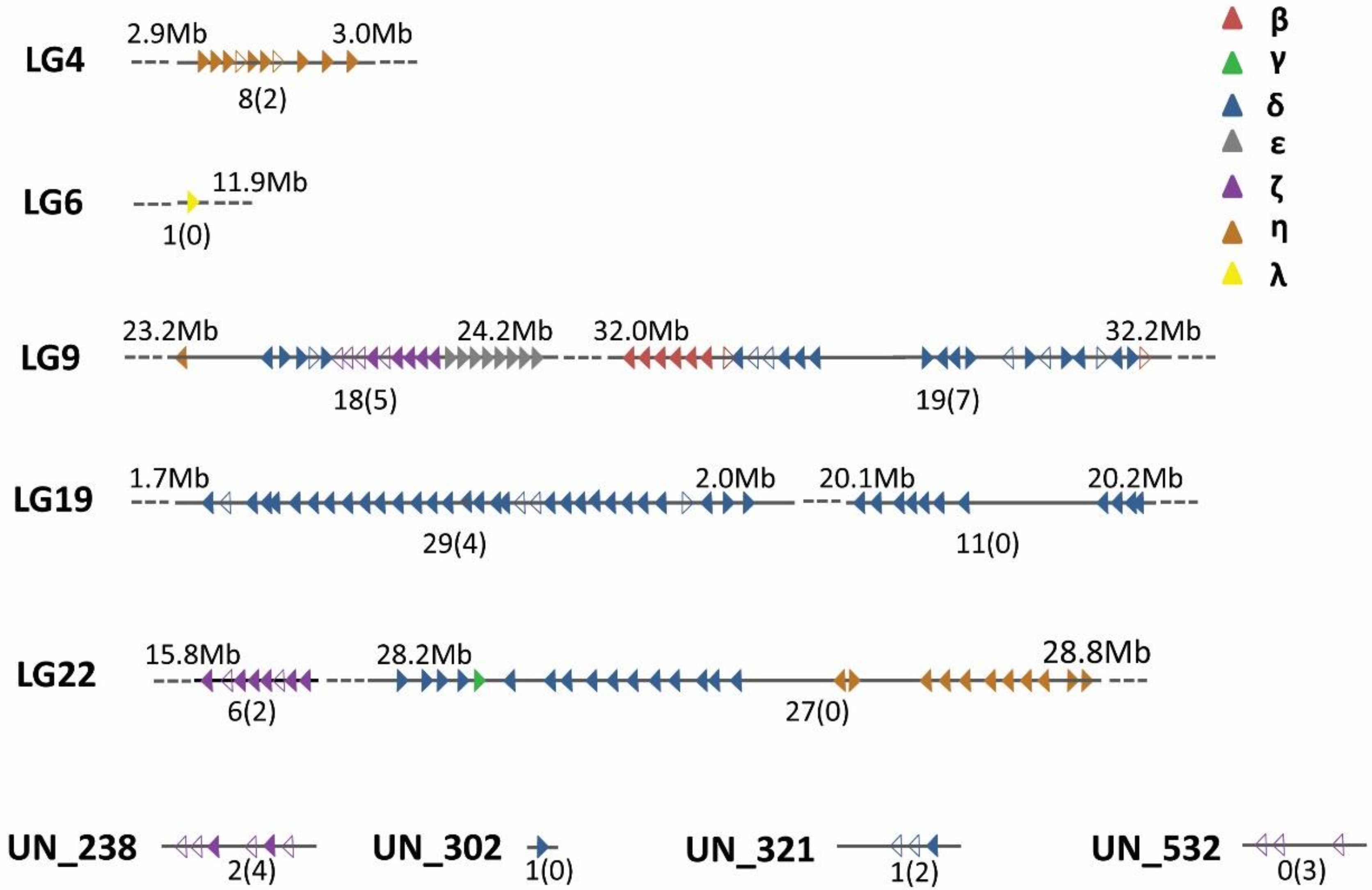
3.1. Genomic Organization of the Chinese Perch OR Repertoire
3.2. Classification of the OR Gene Repertoire
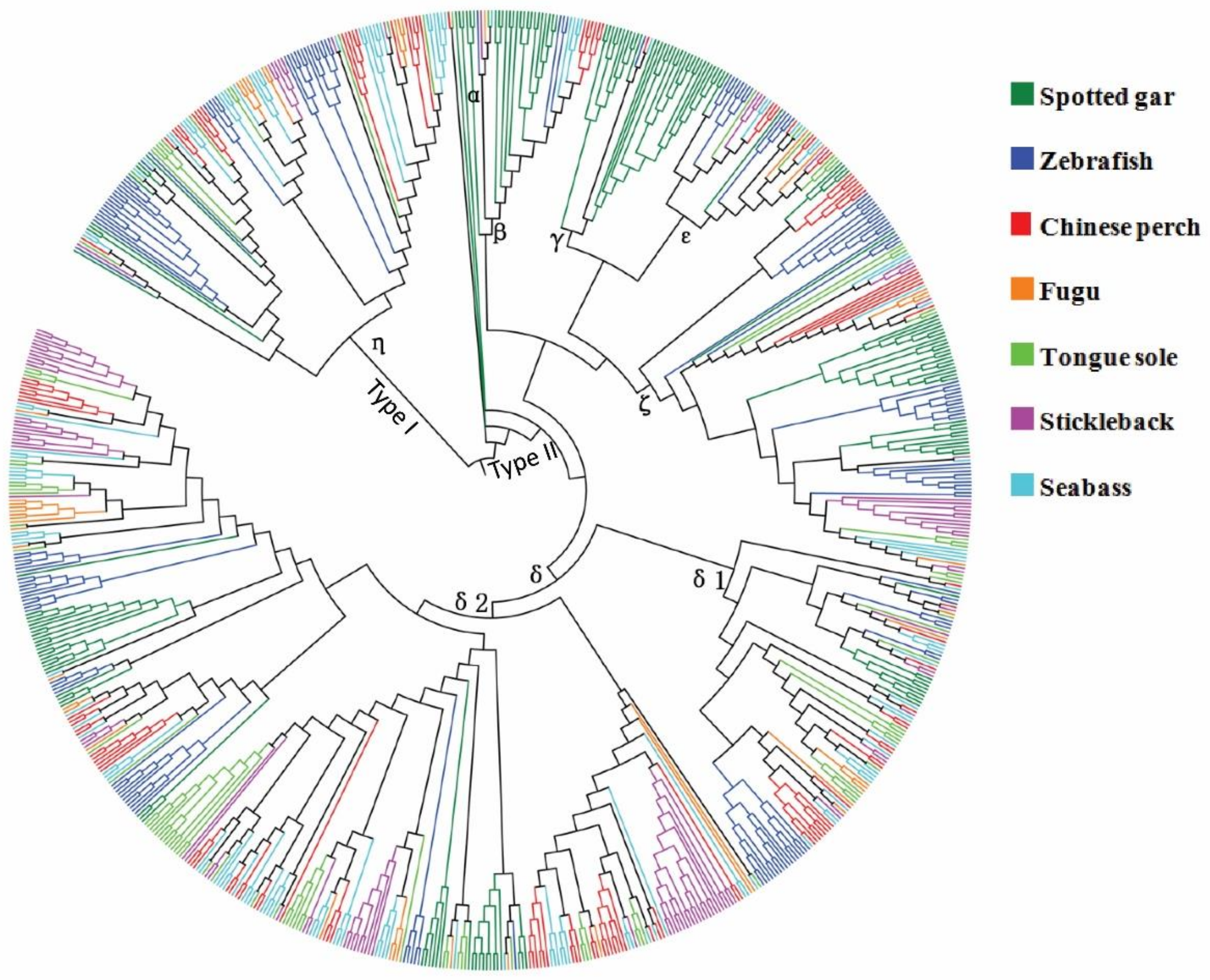
3.3. Phylogenetic Analysis of the OR Gene Repertoire
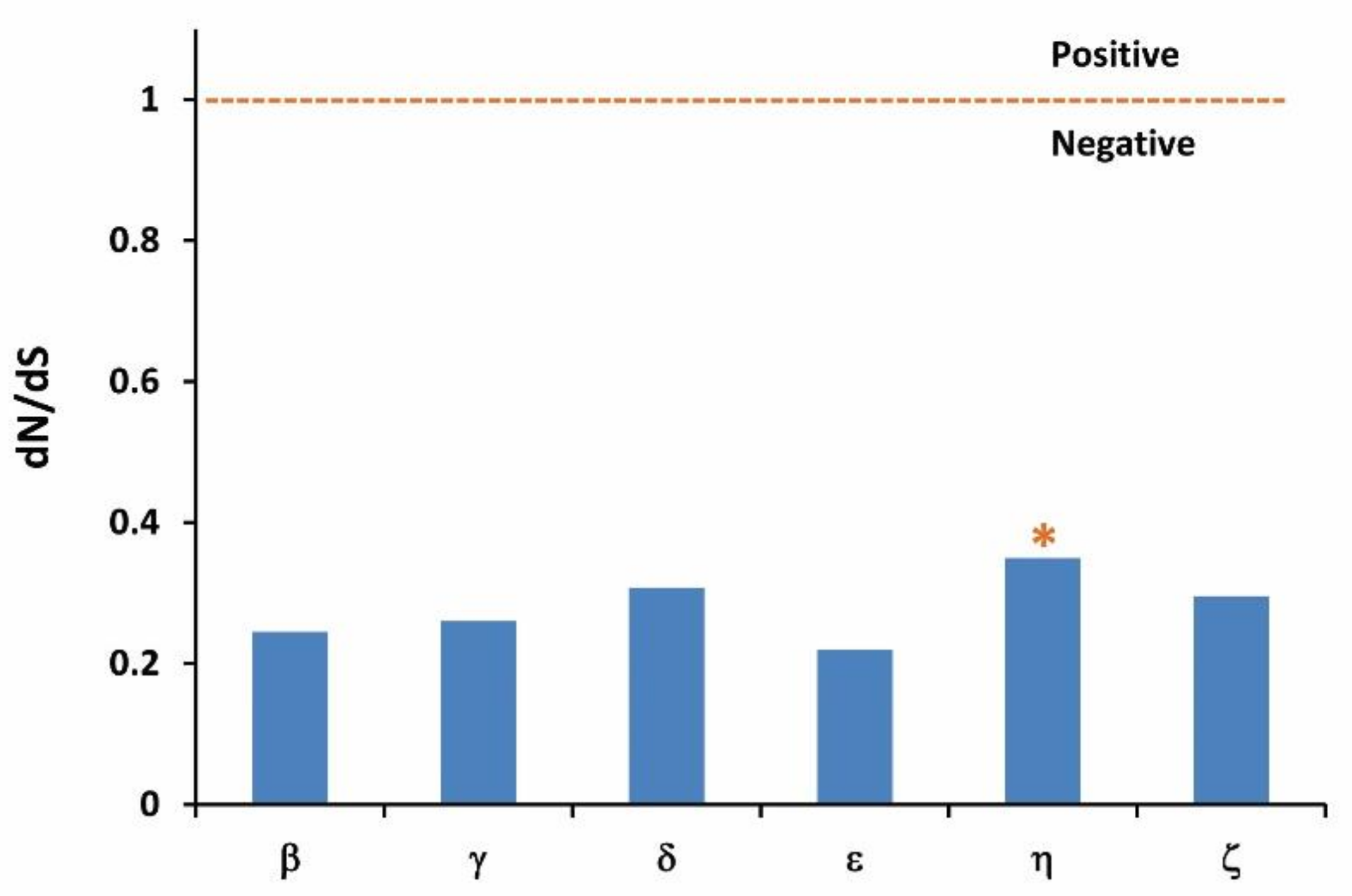
3.4. Evolution of dN/dS Ratio
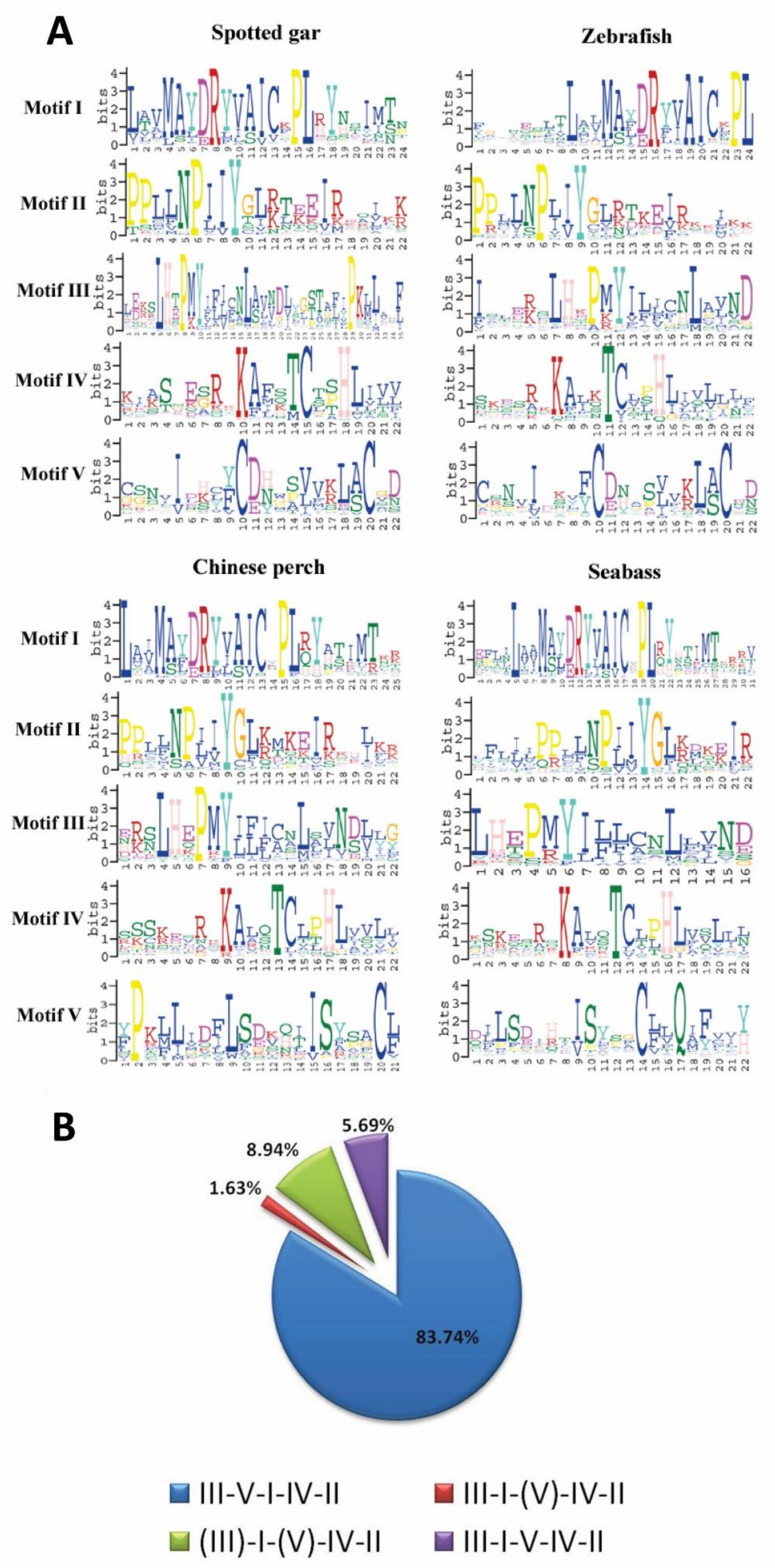
3.5. Patterns of Conserved Motifs for OR Genes
3.6. Potential Species-Specific OR Genes in Chinese Perch
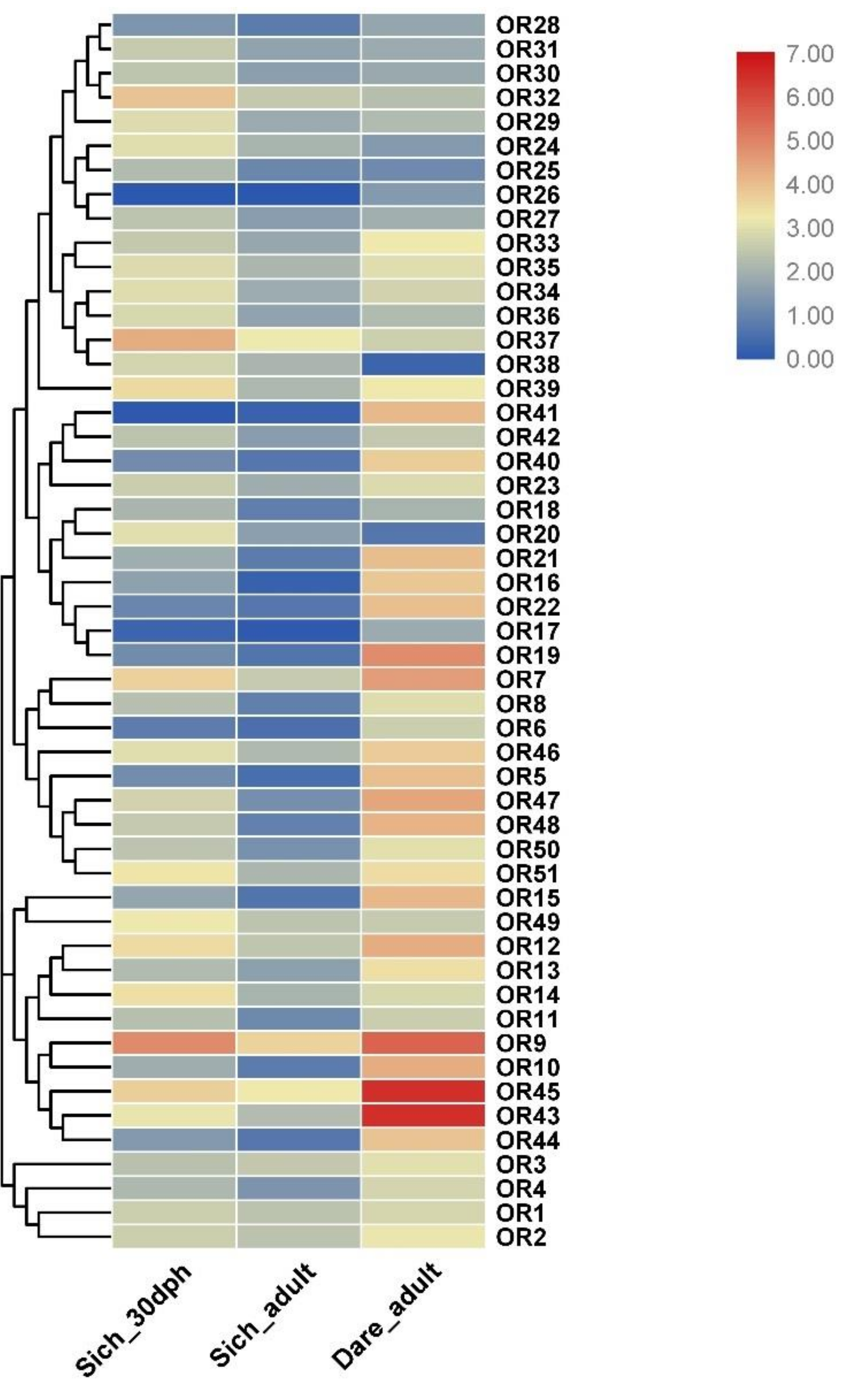
3.7. Expression Profile Analysis of OR Genes at Different Developmental Stages
4. Discussion
4.1. Characterization of OR Genes in Chinese Perch
4.2. Phylogeny of OR Genes in Chinese Perch
4.3. Expression Levels of OR Genes in Chinese Perch OE
4.4. Adaptive Evolution of OR Genes in Chinese Perch
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kishida, T.; Thewissen, J.; Hayakawa, T.; Imai, H.; Agata, K. Aquatic adaptation and the evolution of smell and taste in whales. Zool. Lett. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Yang, Z.K.; Maldonado, E.; Li, C.; Zhang, G.J.; Gilbert, M.T.P.; Jarvis, E.D.; O’Brien, S.J.; Johnson, W.E.; Antunes, A. Olfactory receptor subgenomes linked with broad ecological adaptations in Sauropsida. Mol. Biol. Evol. 2015, 32, 2832–2843. [Google Scholar] [CrossRef] [PubMed]
- Yabuki, Y.; Koide, T.; Miyasaka, N.; Wakisaka, N.; Masuda, M.; Ohkura, M.; Nakai, J.; Tsuge, K.; Tsuchiya, S.; Sugimoto, Y. Olfactory receptor for prostaglandin F 2α mediates male fish courtship behavior. Nat. Neurosci. 2016, 19, 897. [Google Scholar] [CrossRef] [PubMed]
- Piersanti, S.; Frati, F.; Conti, E.; Gaino, E.; Rebora, M.; Salerno, G. First evidence of the use of olfaction in Odonata behaviour. J. Insect Physiol. 2014, 62, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Adipietro, K.A.; Mainland, J.D.; Matsunami, H. Functional evolution of mammalian odorant receptors. PLoS Genet. 2012, 8, e1002821. [Google Scholar] [CrossRef] [PubMed]
- Feldmesser, E.; Olender, T.; Khen, M.; Yanai, I.; Ophir, R.; Lancet, D. Widespread ectopic expression of olfactory receptor genes. BMC Genom. 2006, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.L.; Wang, L.J.; Tang, W.Q.; Wang, X.M.; Wang, C. Identification of olfactory receptor genes in the Japanese grenadier anchovy Coilia nasus. Genes Genom. 2017, 39, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, K.A.; Lubieniecki, K.P.; Koop, B.F.; Davidson, W.S. Identification of olfactory receptor genes in Atlantic salmon Salmo salar. J. Fish. Biol. 2012, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Branscomb, A.; Seger, J.; White, R.L. Evolution of odorant receptors expressed in mammalian testes. Genetics 2000, 156, 785–797. [Google Scholar] [PubMed]
- Buck, L.B. A novel multigene family may encode odorant receptors. Soc. Gen. Physiol. Ser. 1992, 47, 39–51. [Google Scholar] [PubMed]
- Niimura, Y. On the origin and evolution of vertebrate olfactory receptor genes: Comparative genome analysis among 23 Chordate species. Genome Biol. Evol. 2009, 1, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Niimura, Y.; Nei, M. Evolutionary dynamics of olfactory receptor genes in fishes and tetrapods. Proc. Natl. Acad. Sci. USA 2005, 102, 6039–6044. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wang, K.; Lei, F.M.; Yu, D.; Zhao, H.B. Penguins reduced olfactory receptor genes common to other waterbirds. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Tsagkogeorga, G.; Muller, S.; Dessimoz, C.; Rossiter, S.J. Comparative genomics reveals contraction in olfactory receptor genes in bats. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Hayden, S.; Bekaert, M.; Crider, T.A.; Mariani, S.; Murphy, W.J.; Teeling, E.C. Ecological adaptation determines functional mammalian olfactory subgenomes. Genome Res. 2010, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Niimura, Y.; Matsui, A.; Touhara, K. Extreme expansion of the olfactory receptor gene repertoire in African elephants and evolutionary dynamics of orthologous gene groups in 13 placental mammals. Genome Res. 2014, 24, 1485–1496. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Niimura, Y.; Nozawa, M. The evolution of animal chemosensory receptor gene repertoires: Roles of chance and necessity. Nat. Rev. Genet. 2008, 9, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Gilad, Y.; Wiebel, V.; Przeworski, M.; Lancet, D.; Paabo, S. Loss of olfactory receptor genes coincides with the acquisition of full trichromatic vision in primates. PLoS Biol. 2004, 2, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.; Khan, I.; Johnson, W.E.; Gilbert, M.T.P.; Zhang, G.J.; Jarvis, E.D.; O’Brien, S.J.; Antunes, A. Gene loss, adaptive evolution and the co-evolution of plumage coloration genes with opsins in birds. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Hart, N.S. The visual ecology of avian photoreceptors. Prog. Retin. Eye Res. 2001, 20, 675–703. [Google Scholar] [CrossRef]
- Zhou, Y.S.; Yan, X.J.; Xu, S.L.; Zhu, P.; He, X.X.; Liu, J.X. Family structure and phylogenetic analysis of odorant receptor genes in the large yellow croaker (Larimichthys crocea). BMC Evol. Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.F.; Liu, J.K.; Huang, B.Y. The role of sense organs in the feeding behaviour of Chinese perch. J. Fish. Biol. 1998, 52, 1058–1067. [Google Scholar] [CrossRef]
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 25 February 2019).
- UCSC Genome Bioinformatics. Dicentrarchus labrax (dicLab v1.0c). Available online: http://seabass.mpipz.mpg.de/ (accessed on 25 February 2019).
- EMBOSS Explorer. Available online: http://emboss.bioinformatics.nl/cgi-bin/emboss/getorf/ (accessed on 25 February 2019).
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Delport, W.; Poon, A.F.Y.; Frost, S.D.W.; Pond, S.L.K. Datamonkey 2010: A suite of phylogenetic analysis tools for evolutionary biology. Bioinformatics 2010, 26, 2455–2457. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Gojobori, T. A method for detecting positive selection at single amino acid sites. Mol. Biol. Evol. 1999, 16, 1315–1328. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-glycosylation sites in human proteins. 2004. in preparation. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/.
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Gojobori, T. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol. Biol. Evol. 1986, 3, 418–426. [Google Scholar] [PubMed]
- Azzouzi, N.; Barloy-Hubler, F.; Galibert, F. Inventory of the cichlid olfactory receptor gene repertoires: identification of olfactory genes with more than one coding exon. BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, L.R.; Ahuja, G.; Ivandic, I.; Syed, A.S.; Marioni, J.C.; Korsching, S.I.; Logan, D.W. Molecular and neuronal homology between the olfactory systems of zebrafish and mouse. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Gilad, Y.; Bustamante, C.D.; Lancet, D.; Paabo, S. Natural selection on the olfactory receptor gene family in humans and chimpanzees. Am. J. Hum. Genet. 2003, 73, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Lee, K.; Choi, H.; Choi, M.K.; Le, M.T.; Song, N.; Kim, J.H.; Seo, H.G.; Oh, J.W.; Lee, K.; Kim, T.H.; Park, C. The complete swine olfactory subgenome: Expansion of the olfactory gene repertoire in the pig genome. BMC Genom. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Conides, A.; Klaoudatos, D.; Glamuzina, B.; Dulcic, J. Observations on the foraging behaviour of adult broodstock European sea bass, Dicentrarchus labrax (L. 1758) in captivity and conceptualisation for the farming of the species. Acta Adriat. 2016, 57, 107–113. [Google Scholar]
- Lee, K.; Nguyen, D.T.; Choi, M.; Cha, S.Y.; Kim, J.H.; Dadi, H.; Seo, H.G.; Seo, K.; Chun, T.; Park, C. Analysis of cattle olfactory subgenome: The first detail study on the characteristics of the complete olfactory receptor repertoire of a ruminant. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Alioto, T.S.; Ngai, J. The odorant receptor repertoire of teleost fish. BMC Genom. 2005, 6, 173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Firestein, S. The olfactory receptor gene superfamily of the mouse. Nat. Neurosci. 2002, 5, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Peatman, E.; Tang, H.; Lewis, J.; Liu, Z. Profiling of gene duplication patterns of sequenced teleost genomes: Evidence for rapid lineage-specific genome expansion mediated by recent tandem duplications. BMC Genom. 2012, 13, 246. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Zardoya, R.; Meyer, A. Evolutionary relationships of the coelacanth, lungfishes, and tetrapods based on the 28S ribosomal RNA gene. Proc. Natl. Acad. Sci. USA 1996, 93, 5449–5454. [Google Scholar] [CrossRef] [PubMed]
- Picone, B.; Hesse, U.; Panji, S.; Van Heusden, P.; Jonas, M.; Christoffels, A. Taste and odorant receptors of the coelacanth-A gene repertoire in transition. J. Exp. Zool. Part B 2014, 322, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Braasch, I.; Gehrke, A.R.; Smith, J.J.; Kawasaki, K.; Manousaki, T.; Pasquier, J.; Amores, A.; Desvignes, T.; Batzel, P.; Catchen, J. The spotted gar genome illuminates vertebrate evolution and facilitates human-teleost comparisons. Nat. Genet. 2016, 48, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liu, S.; Yao, J.; Li, N.; Yuan, Z.; Zhou, T.; Li, Q.; Liu, Z. Genomic organization and evolution of olfactory receptors and trace amine-associated receptors in channel catfish, Ictalurus punctatus. Biochim. Biophys. Acta Gen. Subj. 2016, 1861, 644–651. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liang, X.F.; Sun, J.; Li, L.; Yu, Y.; Huang, W.; Qu, C.M.; Cao, L.; Bai, X.L.; Tao, Y.X. Insights into food preference in hybrid F1 of Siniperca chuatsi (female) x Siniperca scherzeri (male) mandarin fish through transcriptome analysis. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Makalowski, W.; Boguski, M.S. Evolutionary parameters of the transcribed mammalian genome: An analysis of 2820 orthologous rodent and human sequences. Proc. Natl. Acad. Sci. USA 1998, 95, 9407–9412. [Google Scholar] [CrossRef]
- Young, J.M.; Shykind, B.M.; Lane, R.P.; Tonnes-Priddy, L.; Ross, J.A.; Walker, M.; Williams, E.M.; Trask, B.J. Odorant receptor expressed sequence tags demonstrate olfactory expression of over 400 genes, extensive alternate splicing and unequal expression levels. Genome Biol. 2003, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, E.M.; Zhang, W.Y.; Gelis, L.; Deng, Y.; Noldus, J.; Hatt, H. Activation of an olfactory receptor inhibits proliferation of prostate cancer cells. J. Biol. Chem. 2009, 284, 16218–16225. [Google Scholar] [CrossRef] [PubMed]
- Sacchettini, J.C.; Poulter, C.D. Biochemistry—Creating isoprenoid diversity. Science 1997, 277, 1788–1789. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef]
- Kajiya, K.; Inaki, K.; Tanaka, M.; Haga, T.; Kataoka, H.; Touhara, K. Molecular bases of odor discrimination: Reconstitution of olfactory receptors that recognize overlapping sets of odorants. J. Neurosci. 2001, 21, 6018–6025. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome Number | No. of Functional Genes | No. of Pseudogenes (%) | Total | No. of Clusters | |
|---|---|---|---|---|---|
| LG4 | 8 | 2 | (20) | 10 | 1 |
| LG6 | 1 | 0 | (0) | 1 | 1 |
| LG9 | 37 | 12 | (24) | 49 | 2 |
| LG19 | 40 | 4 | (9) | 44 | 2 |
| LG22 | 33 | 2 | (6) | 35 | 2 |
| LGUN | 4 | 9 | - | 13 | - |
| Total | 123 | 29 | (19) | 152 | 8 |
| Species | α | β | γ | δ | ε | ζ | η | Non-OR | Total | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Spotted gar | 3 | 18 | 34(10) | 55(2) | 3 | 34(2) | 8 | 1 | 156(14) | This study |
| Zebrafish | 0 | 4(2) | 1 | 62(7) | 12(1) | 37(4) | 38(7) | 0 | 154(21) | [11] |
| Chinese perch | 0 | 6(1) | 1 | 74(13) | 8 | 13(13) | 22(2) | 1 | 123(29) | This study |
| Seabass | 0 | 5(1) | 1 | 79(2) | 6(1) | 9 | 31 | 0 | 131(4) | This study |
| Tongue sole | 0 | 1 | 0 | 62(4) | 6 | 10 | 16 | 1 | 96(4) | This study |
| Stickleback | 0 | 1 | 0(3) | 71(41) | 4 | 18(4) | 8(4) | 0 | 102(52) | [11] |
| Fugu | 0 | 1 | 0 | 30(25) | 2(1) | 4(2) | 10(11) | 0 | 47(39) | [11] |
| Number of OR Genes Belonging to Common Subfamilies in the Species | |||||
|---|---|---|---|---|---|
| Sich | Dila | Cyse | Taru | Gaac | |
| Sich, Dila, Cyse, Taru, Gaac | 15 | 11 | 11 | 5 | 16 |
| Sich, Dila, Cyse, Taru | 3 | 3 | 1 | 1 | - |
| Sich, Dila, Cyse, Gaac | 16 | 21 | 8 | - | 10 |
| Sich, Dila, Taru, Gaac | 35 | 22 | - | 4 | 5 |
| Sich, Cyse, Taru, Gaac | 3 | - | 4 | 2 | 2 |
| Dila, Cyse, Taru, Gaac | - | 5 | 6 | 2 | 14 |
| Sich, Dila, | 28 | 23 | - | - | - |
| Sich | 7 | - | - | - | - |
| Dila | - | 13 | - | - | - |
| Cyse | - | - | 50 | - | - |
| Taru | - | - | - | 9 | - |
| Gaac | - | - | - | - | 22 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, L.-Y.; Liang, X.-F.; He, S. Genome-Wide Identification and Characterization of Olfactory Receptor Genes in Chinese Perch, Siniperca chuatsi. Genes 2019, 10, 178. https://doi.org/10.3390/genes10020178
Lv L-Y, Liang X-F, He S. Genome-Wide Identification and Characterization of Olfactory Receptor Genes in Chinese Perch, Siniperca chuatsi. Genes. 2019; 10(2):178. https://doi.org/10.3390/genes10020178
Chicago/Turabian StyleLv, Li-Yuan, Xu-Fang Liang, and Shan He. 2019. "Genome-Wide Identification and Characterization of Olfactory Receptor Genes in Chinese Perch, Siniperca chuatsi" Genes 10, no. 2: 178. https://doi.org/10.3390/genes10020178