Satellite DNA Mapping in Pseudis fusca (Hylidae, Pseudinae) Provides New Insights into Sex Chromosome Evolution in Paradoxical Frogs

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Cytogenetic Analysis
2.1.1. Individuals, Chromosome Preparations, and Karyotype Description
2.1.2. Amplification of PcP190 Satellite DNA for Further Use as a Probe in Southern Blotting and In Situ Hybridization
2.1.3. Southern Blot Detection of PcP190 Satellite DNA
2.1.4. Fluorescent In Situ Hybridization and Comparative Genomic Hybridization
2.2. Phylogenetic and Genetic Distance Analyses
2.2.1. Taxon and Gene Sampling
2.2.2. DNA Extraction and Sequencing of H1 and Cytb Fragments
2.2.3. Sequence Analyses
3. Results
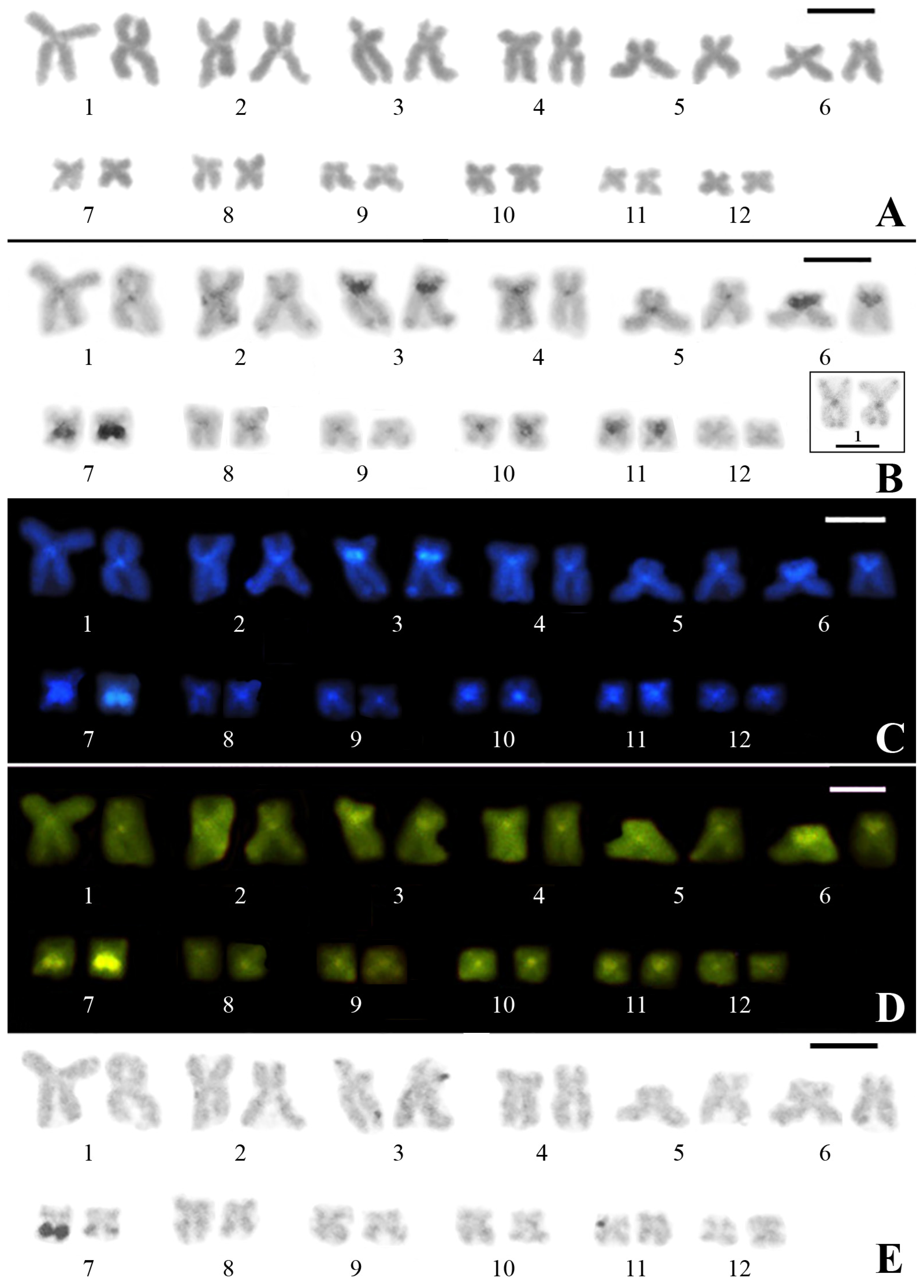
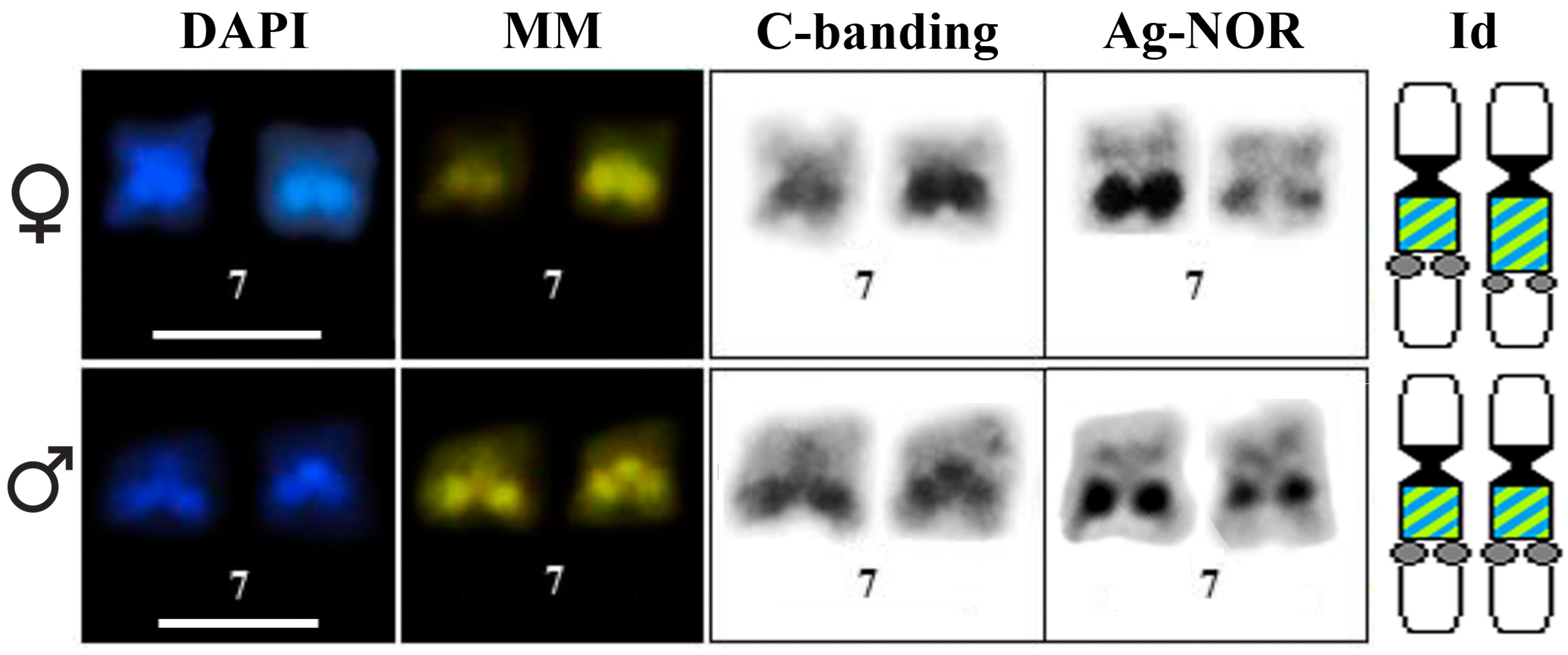
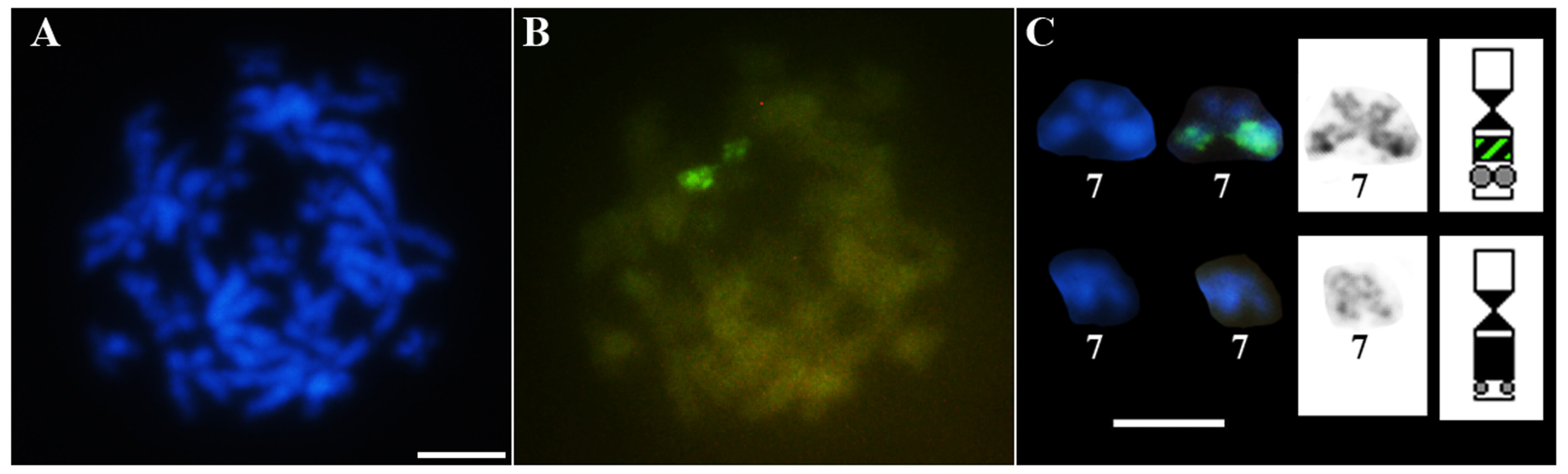
3.1. Classical Cytogenetic Analysis of Pseudis fusca from Carlos Chagas–MG
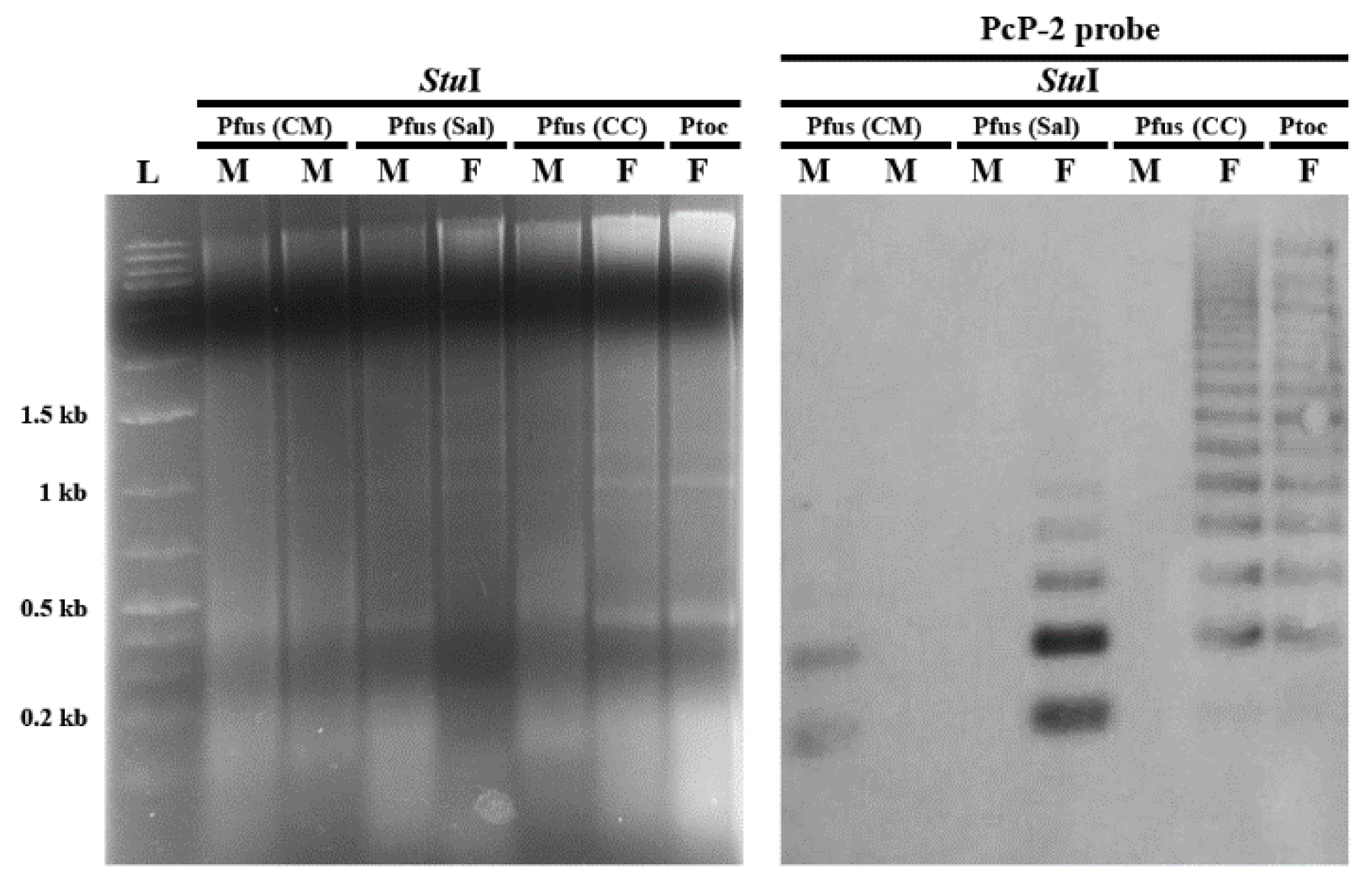
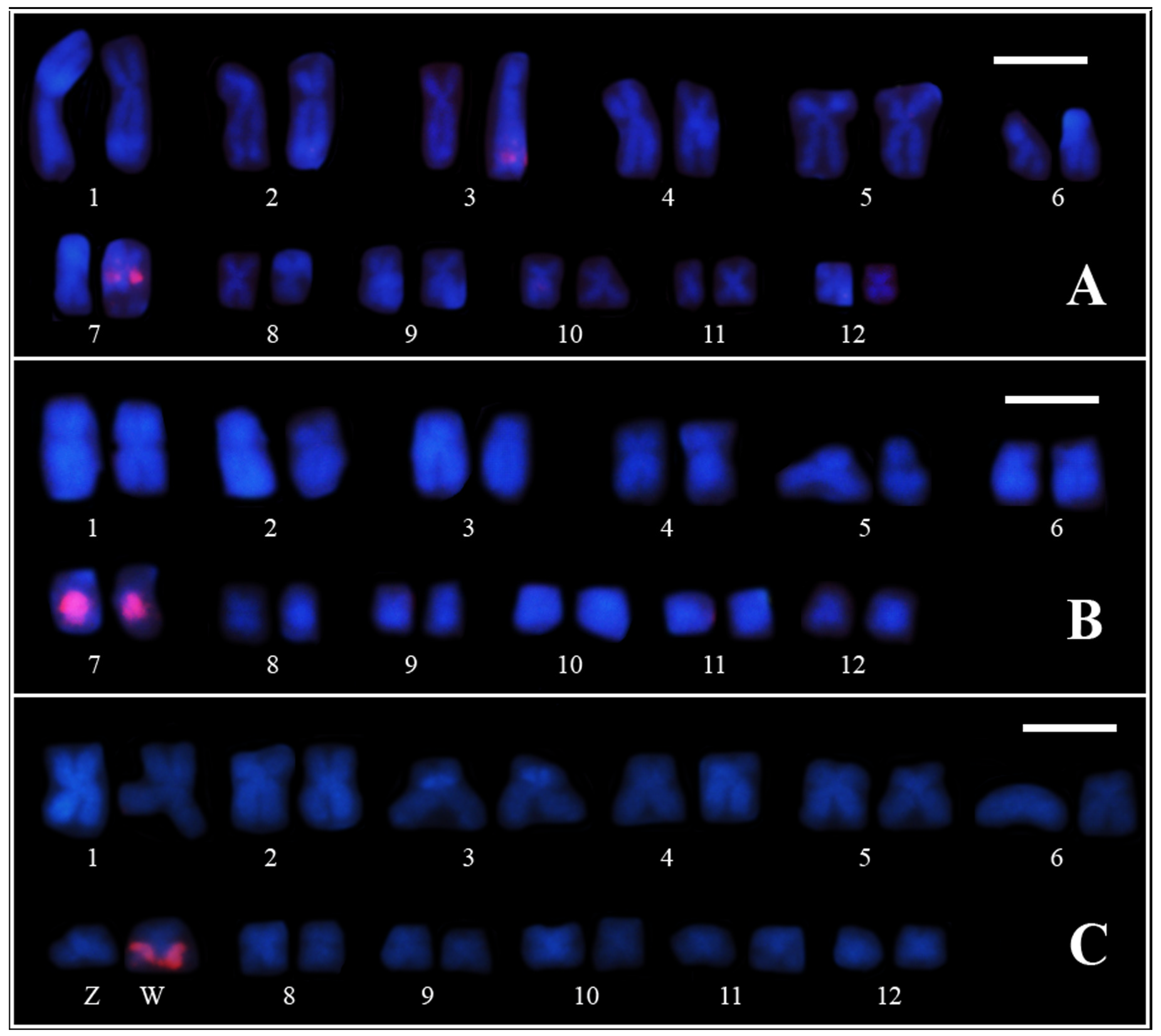
3.2. PcP190 satellite DNA and Comparative Genomic Hybridization
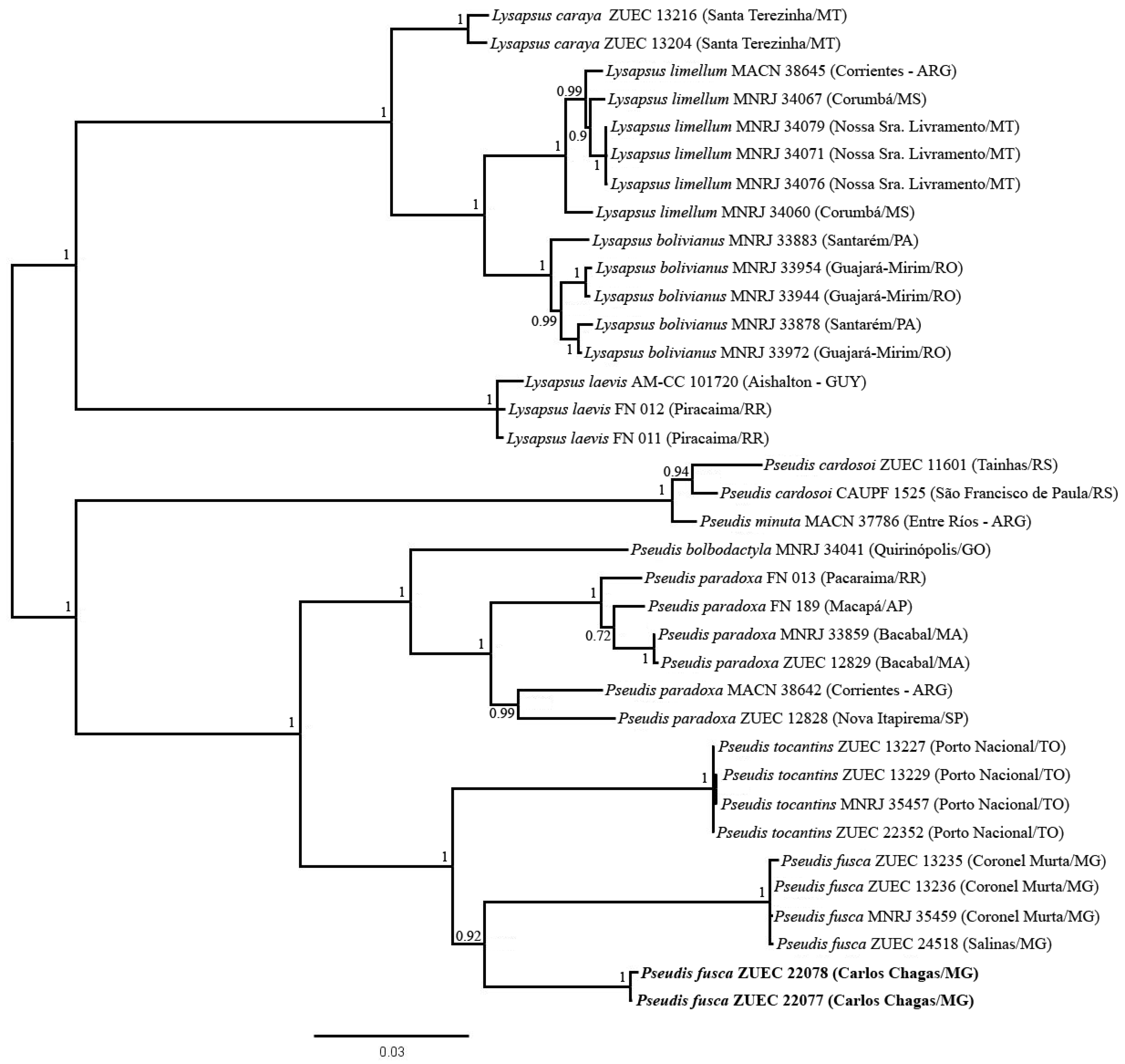
3.3. Phylogenetic Inferences and Genetic Distances
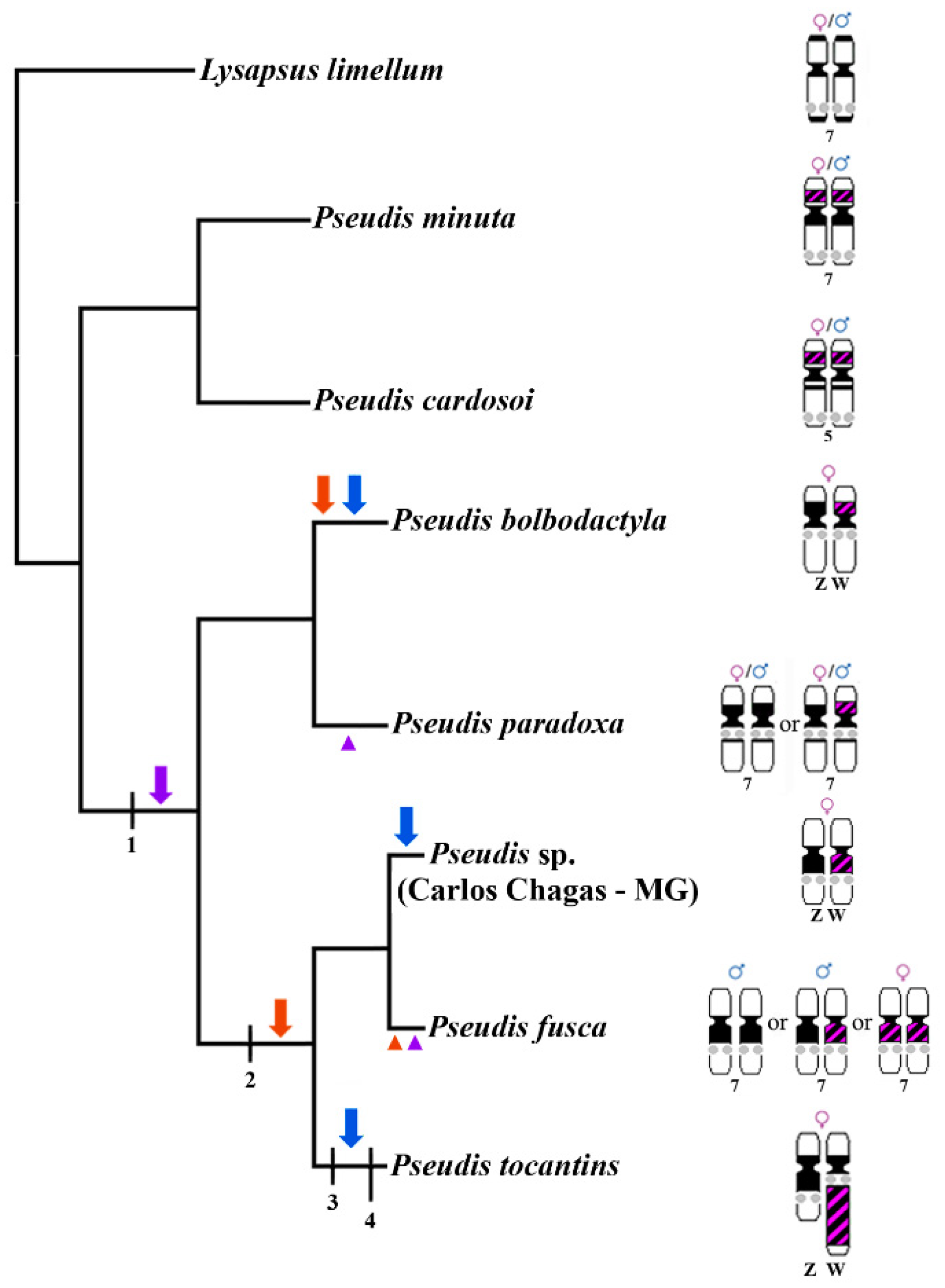
4. Discussion
4.1. Genetic Divergence: Could Pseudis fusca from Carlos Chagas be a New Species?
4.2. PcP190 Sequences of Pseudis fusca
4.3. Chromosome Rearrangements as a Major Force of Sex Chromosome Differentiation in Pseudis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Graves, J.A.M. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: historical insights and future perspectives. Proc. Biol. Sci. 2017, 284, 20162806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity. 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellegren, H. Sex-chromosome evolution: Recent progress and the influence of male and female heterogamety. Nat. Rev. Genet. 2011, 12, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Targueta, C.P.; Vittorazzi, S.E.; Gatto, K.P.; Bruschi, D.P.; Veiga-Menoncello, A.C.P.; Recco-Pimentel, S.M.; Lourenço, L.B. Anuran Cytogenetics: An overview. In An Essential Guide to Cytogenetics; Norris, N., Miller, C., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2018; pp. 1–64. [Google Scholar]
- Ferro, J.M.; Cardozo, D.E.; Suárez, P.; Boeris, J.M.; Blasco-Zúñiga, A.; Barbero, G.; Gomes, A.; Gazoni, T.; Costa, W.; Nagamachi, C.Y.; et al. Chromosome evolution in Cophomantini (Amphibia, Anura, Hylinae). PLoS ONE 2018, 13, e0192861. [Google Scholar] [CrossRef] [PubMed]
- Gazoni, T.; Haddad, C.F.B.; Narimatsu, H.; Cabral-de-Mello, D.C.; Lyra, M.L.; Parise-Maltempi, P.P. More sex chromosomes than autosomes in the Amazonian frog Leptodactylus pentadactylus. Chromosoma 2018, 127, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Busin, C.S.; Andrade, G.V.; Bertoldo, J.; Del Grande, M.L.; Uetanabaro, M.; Recco-Pimentel, S.M. Cytogenetic analysis of four species of Pseudis (Anura, Hylidae), with the description of ZZ/ZW sex chromosomes in P. tocantins. Genetica 2008, 133, 119–127. [Google Scholar] [CrossRef]
- Gatto, K.P.; Busin, C.S.; Lourenço, L.B. Unraveling the sex chromosome heteromorphism of the paradoxical frog Pseudis tocantins. PLoS ONE 2016, 11, e0156176. [Google Scholar] [CrossRef]
- Gatto, K.P.; Mattos, J.V.; Seger, K.R.; Lourenço, L.B. Sex chromosome differentiation in the frog genus Pseudis involves satellite DNA and chromosome rearrangements. Front. Genet. 2018, 9, 301. [Google Scholar] [CrossRef]
- Vittorazzi, S.E.; Lourenço, L.B.; Del-Grande, M.L.; Recco-Pimentel, S.M. Satellite DNA derived from 5S rDNA in Physalaemus cuvieri (Anura, Leiuperidae). Cytogenet. Genome Res. 2011, 134, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Vittorazzi, S.E.; Lourenço, L.B.; Recco-Pimentel, S.M. Long-time evolution and highly dynamic satellite DNA in leptodactylid and hylodid frogs. BMC Genet. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Rofe, R. Karyotypic variation in the Australian gekko Phyllodactylus marmoratus (Gray) (Gekkonidae: Reptilia). Chromosoma 1976, 54, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled silver staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.R.; Lourenço, L.B.; Rossa-Feres, D.C.; Lima, A.P.; Andrade, G.V.; Giaretta, A.A.; Egito, G.T.B.T.; Recco-Pimentel, S.M. Comparative cytogenetic analysis of some species of the Dendropsophus microcephalus group (Anura, Hylidae) in the light of phylogenetic inferences. BMC Genet. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritschi, E.F.; Maniats, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Viegas-Péquignot, E. In situ hybridization to chromosomes with biotinylated probes. In In Situ Hybridization: A Practical Approach; Wilkinson, D.G., Ed.; Oxford University Press: Oxford, UK, 1992; pp. 137–158. [Google Scholar]
- Garda, A.A.; Santana, D.J.; de São-Pedro, V.A. Taxonomic characterization of Paradoxical frogs (Anura, Hylidae, Pseudae): Geographic distribution, external morphology, and morphometry. Zootaxa 2010, 2666, 1–28. [Google Scholar] [CrossRef]
- Santana, D.J.; de Magalhães, F.M.; de São-Pedro, V.A.; Mângia, S.; Amado, T.F.; Garda, A.A. Calls and tadpoles of the species of Pseudis (Anura, Hylidae, Pseudae). Herpetol. J. 2016, 26, 139–148. [Google Scholar]
- Wiens, J.J.; Kuczynski, C.A.; Hua, X.; Moen, D.S. An expanded phylogeny of treefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Mol. Phylogenet. Evol. 2010, 55, 871–882. [Google Scholar] [CrossRef]
- Duellman, W.E.; Marion, A.B.; Hedges, S.B. Phylogenetics, classification, and biogeography of the treefrogs (Amphibia: Anura: Arboranae). Zootaxa 2016, 4104, 1–109. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The simple fool’s guide to PCR version 2. Univ. Hawaii 2002, 1–45. [Google Scholar]
- Garda, A.A.; Cannatella, D.C. Phylogeny and biogeography of paradoxical frogs (Anura, Hylidae, Pseudae) inferred from 12S and 16S mitochondrial DNA. Mol. Phylogenet. Evol. 2007, 44, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Graybeal, A. Phylogenetic relationships of bufonid frogs and tests of alternate macroevolutionary hypotheses characterizing their radiation. Zool. J. Linn. Soc. 1997, 119, 297–338. [Google Scholar] [CrossRef] [Green Version]
- Feller, A.E.; Hedges, S.B. Molecular evidence for the early history of living amphibians. Mol. Phylogenet. Evol. 1998, 9, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.; Schneider, C.J.; Wake, D.B. Evolutionary relationships within the Ensatina eschscholtzii complex confirm the ring species interpretation. Syst. Biol. 1992, 41, 273–291. [Google Scholar] [CrossRef]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef]
- Seger, K.R.; Teixeira, B.F.V.; Rossa-Feres, D.C.; Lima, A.P.; Andrade, G.V.; Giaretta, A.A.; Lourenço, L.B. Five independent lineages revealed by integrative taxonomy in the Dendropsophus nanus species complex (Anura, Hylidae). Unpublished work. 2019. [Google Scholar]
- Hedges, S.B. Molecular evidence for the origin of birds. Proc. Natl. Acad. Sci. USA 1994, 91, 2621–2624. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2 2004. Distributed by the author. Evolutionary Biology Centre, Uppsala University.
- Vences, M.; Thomas, M.; Bonett, R.M.; Vieites, D.R. Deciphering amphibian diversity through DNA barcoding: chances and challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1859–1868. [Google Scholar] [CrossRef] [Green Version]
- Fouquet, A.; Gilles, A.; Vences, M.; Marty, C.; Blanc, M.; Gemmell, N.J. Underestimation of species richness in Neotropical frogs revealed by mtDNA analyses. PLoS ONE 2007, 2, e1109. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Aguiar-Júnior, O.; Bacci Jr, M.; Lima, A.P.; Rossa-Feres, D.D.C.; Haddad, C.F.B.; Recco-Pimentel, S.M. Phylogenetic relationships of Pseudis and Lysapsus (Anura, Hylidae, Hylinae) inferred from mitochondrial and nuclear gene sequences. Cladistics 2007, 23, 455–463. [Google Scholar] [CrossRef]
- Díaz, L.M.; Hedges, S.B.; Schmid, M. A new cryptic species of the genus Eleutherodactylus (Amphibia: Anura: Eleutherodactylidae) from Cuba. Zootaxa 2012, 3220, 44–60. [Google Scholar]
- Nuñez, J.J.; Rabanal, F.E.; Formas, J.R. Description of a new species of Eupsophus (Amphibia: Neobatrachia) from the Valdivian Coastal range, Southern Chile: an integrative taxonomic approach. Zootaxa 2012, 3305, 53–68. [Google Scholar] [CrossRef]
- Bruschi, D.P.; Lucas, E.M.; Garcia, P.C.A.; Recco-Pimentel, S.M. Molecular and morphological evidence reveals a new species in the Phyllomedusa hypochondrialis group (Hylidae, Phyllomedusinae) from the Atlantic Forest of the highlands of Southern Brazil. PLoS ONE 2014, 9, e105608. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.J.; Carter, T.F.; Greenbaum, E.; Gvo, V.; Kelley, D.B.; Mclaughlin, P.J.; Pauwels, O.S.G.; Portik, D.M.; Stanley, E.L.; Tinsley, R.C.; Tobias, M.L.; Blackburn, D.C. Genetics, morphology, advertisement calls, and historical records distinguish six new polyploid species of African clawed frog (Xenopus, Pipidae) from West and Central Africa. PLoS ONE 2015, 10, e0142823. [Google Scholar] [CrossRef]
- Vences, M.; Thomas, M.; Van Der Meijden, A.; Chiari, Y.; Vieites, D.R. Comparative performance of 16S rRNA gene in DNA barcoding of amphibians. Front. Zool. 2005, 2. [Google Scholar] [CrossRef]
- Amador, L.; Parada, A.; D’Elía, G.; Guayasamin, J.M. Uncovering hidden specific diversity of Andean glassfrogs of the Centrolene buckleyi species complex (Anura: Centrolenidae). PeerJ 2018, 6, e5856. [Google Scholar] [CrossRef] [PubMed]
- Blotto, B.L.; Nuñez, J.J.; Basso, N.G.; Úbeda, C.A.; Wheeler, W.C.; Faivovich, J. Phylogenetic relationships of a Patagonian frog radiation, the Alsodes + Eupsophus clade (Anura: Alsodidae), with comments on the supposed paraphyly of Eupsophus. Cladistics 2013, 18, 113–131. [Google Scholar] [CrossRef]
- Busin, C.S.; Vinciprova, G.; Recco-Pimentel, S.M. Chromosomal rearrangements as the source of variation in the number of chromosomes in Pseudis (Amphibia, Anura). Genetica 2001, 110, 131–141. [Google Scholar] [CrossRef]
- Barrio, A.; Rubel, D.P. Caracteristicas del cariótipos de los pseudidos. Physis 1970, 79, 505–510. [Google Scholar]
- Steinemann, S.; Steinemann, M. Retroelements: Tools for sex chromosome evolution. Cytogenet. Genome Res. 2005, 110, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Chalopin, D.; Volff, J.-N.; Galiana, D.; Anderson, J.L.; Schartl, M. Transposable elements and early evolution of sex chromosomes in fish. Chromosome Res. 2015, 23, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Steflova, P.; Tokan, V.; Vogel, I.; Lexa, M.; Macas, J.; Novak, P.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Contrasting patterns of transposable element and satellite distribution on sex chromosomes (XY1Y2) in the dioecious plant Rumex acetosa. Genome Biol. Evol. 2013, 5, 769–782. [Google Scholar] [CrossRef]
- Stöck, M.; Savary, R.; Betto-Colliard, C.; Biollay, S.; Jourdan-Pineau, H.; Perrin, N. Low rates of X-Y recombination, not turnovers, account for homomorphic sex chromosomes in several diploid species of Palearctic green toads (Bufo viridis subgroup). J. Evol. Biol. 2013, 26, 674–682. [Google Scholar] [CrossRef]
- Dufresnes, C.; Borzée, A.; Horn, A.; Stöck, M.; Ostini, M.; Sermier, R.; Wassef, J.; Litvinchuck, S.N.; Kosch, T.A.; Waldman, B.; Jang, Y.; Brelsford, A.; Perrin, N. Sex-chromosome homomorphy in Palearctic tree frogs results from both turnovers and X - Y recombination. Mol. Biol. Evol. 2015, 32, 2328–2337. [Google Scholar] [CrossRef]
- Rodrigues, N.; Studer, T.; Dufresnes, C.; Perrin, N. Sex-chromosome recombination in common frogs brings water to the Fountain-of-Youth. Mol. Biol. Evol. 2018, 35, 942–948. [Google Scholar] [CrossRef]
- Evans, B.J.; Pyron, R.A.; Wiens, J.J. Polyploidization and Sex Chromosome Evolution in Amphibians. In Polyploidy and Genome Evolution; Soltis, P.S., Soltis, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 385–410. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
|---|---|---|---|---|---|---|---|
| 1. P. fusca (CM/Sal/Ara/Jeq) | 0/0.13 | 5.98 * | 7.77 | 8.71 | 8.81 | 13.59 | 13.53 |
| 2. P. fusca (Carlos Chagas) | 7.16 | 0/0.17 | 6.30 | 7.79 | 7.82 | 12.64 | 12.85 |
| 3. P. tocantins | 6.99 | 6.08 | 0.42/0.45 | 8.45 | 8.70 | 13.64 | 13.77 |
| 4. P. bolbodactyla | 8.02 | 8.15 | 7.05 | 2.57/2.30 | 5.66 | 12.24 | 12.39 |
| 5. P. paradoxa | 8.88 | 9.12 | 8.15 | 6.07 | 2.40/2.73 | 12.40 | 12.48 |
| 6. P. minuta | 13.64 | 11.82 | 12.55 | 11.83 | 13.01 | 0.20/0.62 | 1.47 |
| 7. P. cardosoi | 13.06 | 11.97 | 12.65 | 11.62 | 12.72 | 1.25 | 0.41/0.39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatto, K.P.; Seger, K.R.; Garcia, P.C.d.A.; Lourenço, L.B. Satellite DNA Mapping in Pseudis fusca (Hylidae, Pseudinae) Provides New Insights into Sex Chromosome Evolution in Paradoxical Frogs. Genes 2019, 10, 160. https://doi.org/10.3390/genes10020160
Gatto KP, Seger KR, Garcia PCdA, Lourenço LB. Satellite DNA Mapping in Pseudis fusca (Hylidae, Pseudinae) Provides New Insights into Sex Chromosome Evolution in Paradoxical Frogs. Genes. 2019; 10(2):160. https://doi.org/10.3390/genes10020160
Chicago/Turabian StyleGatto, Kaleb Pretto, Karin Regina Seger, Paulo Christiano de Anchieta Garcia, and Luciana Bolsoni Lourenço. 2019. "Satellite DNA Mapping in Pseudis fusca (Hylidae, Pseudinae) Provides New Insights into Sex Chromosome Evolution in Paradoxical Frogs" Genes 10, no. 2: 160. https://doi.org/10.3390/genes10020160