Origin, Behaviour, and Transmission of B Chromosome with Special Reference to Plantago lagopus

{kind=link}
Abstract
:1. Introduction
2. Distribution of B Chromosomes
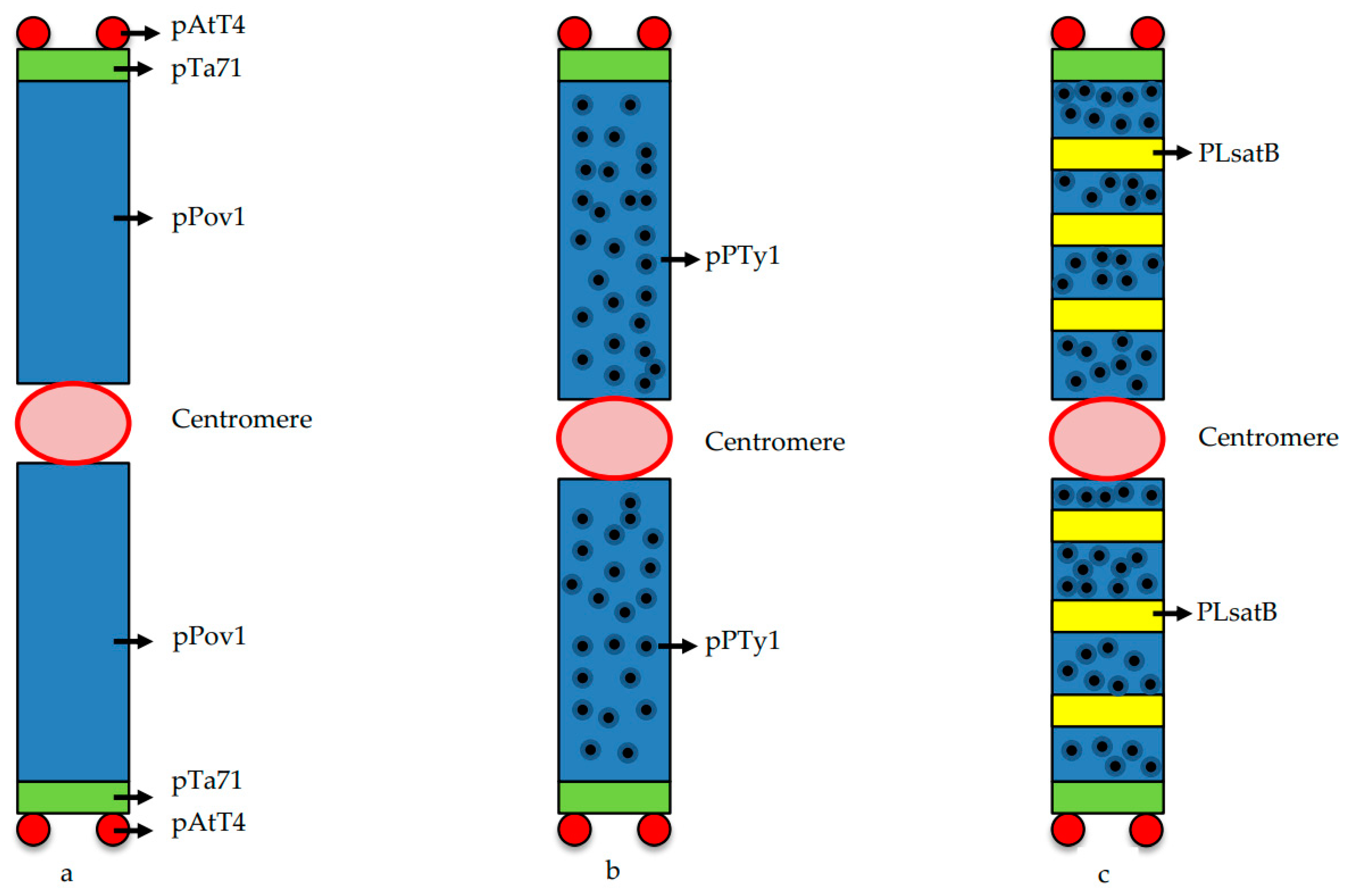
3. Structure of B Chromosomes
4. Molecular Composition
5. Transcriptionally Active Sequences on B Chromosomes
6. Drive Mechanisms and Transmission of B Chromosomes
7. B Chromosome in Relation to Breeding System
8. Evolution of B Chromosomes
9. Outlook
Acknowledgments
Conflicts of Interest
References
- Houben, A.; Banaei-Moghaddam, A.M.; Klemme, S.; Timmis, J.N. Evolution and biology of supernumerary B chromosomes. Cell. Mol. Life Sci. 2014, 71, 467–478. [Google Scholar] [CrossRef]
- Douglas, R.N.; Birchler, J.A. B Chromosomes. In Chromosome Structure and Aberrations; Springer: New Delhi, India, 2017; pp. 13–39. [Google Scholar]
- Carlson, W. The B chromosome of maize. In Handbook of Maize; Springer: New York, NY, USA, 2009; pp. 459–480. [Google Scholar]
- Huang, W.; Du, Y.; Zhao, X.; Jin, W. B chromosome contains active genes and impacts the transcription of A chromosomes in maize (Zea mays L.). BMC Plant Biol. 2016, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Houben, A. B chromosomes—A matter of chromosome drive. Front. Plant Sci. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.; Conte, M.A.; Fantinatti, B.E.A.; Cabral-De-Mello, D.C.; Carvalho, R.F.; Vicari, M.; Kocher, T.; Martins, C. Origin and evolution of B chromosomes in the cichlid fish Astatotilapia latifasciata based on integrated genomic analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef]
- Palestis, B.G.; Trivers, R.; Burt, A.; Jones, R.N. The distribution of B chromosomes across species. Cytogenet. Genome Res. 2004, 106, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.; Martis, M.; Hedley, P.E.; Šimková, H.; Liu, H.; Morris, J.A.; Steuernagel, B.; Taudien, S.; Roessner, S.; Gundlach, H.; et al. Unlocking the barley genome by chromosomal and comparative genomics. Plant Cell 2011, 23, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Kour, G.; Kaul, S.; Dhar, M.K. Molecular characterization of repetitive DNA sequences from B chromosome in Plantago lagopus L. Cytogenet. Genome Res. 2013, 142, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.K.; Koul, A.K. Genetic diversity among Plantagos III. Primary trisomy in Plantago lagopus L. Genetica 1984, 64, 135–138. [Google Scholar] [CrossRef]
- Dhar, M.K.; Friebe, B.; Koul, A.K.; Gill, B.S. Origin of an apparent B chromosome by mutation, chromosome fragmentation and specific DNA sequence amplification. Chromosoma 2002, 111, 332–340. [Google Scholar] [CrossRef]
- Datta, A.K.; Mandal, A.; Das, D.; Gupta, S.; Saha, A.; Paul, R.; Sengupta, S. B chromosomes in angiosperm—A review. Cytol. Genet. 2016, 50, 60–71. [Google Scholar] [CrossRef]
- Jones, R.N.; Viegas, W.; Houben, A. A century of B chromosomes in plants: So what? Ann. Bot. 2007, 101, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Vujosevic, M.; Blagojević, J. B chromosomes in populations of mammals. Cytogenet. Genome Res. 2004, 106, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Dementyeva, P.V.; Graphodatsky, A.S.; Volobouev, V.T.; Kukekova, A.V.; Trifonov, V.A. Genes on B chromosomes of vertebrates. Mol. Cytogenet. 2014, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.A.; Palestis, B.G.; Jones, R.N.; Trivers, R. Phyletic hot spots for B chromosomes in angiosperms. Evolution 2005, 59, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C.; Singh, V.; Bala, S.; Malik, R.A.; Sharma, V.; Kaur, K. Cytomorphological variations and new reports of B-chromosomes in the genus Plantago (Plantaginaceae) from the Northwest Himalaya. Flora 2017, 234, 69–76. [Google Scholar] [CrossRef]
- Frost, S. The cytological behaviour and mode of transmission of accessory chromosomes in Plantago serraria. Hereditas 1959, 45, 191–210. [Google Scholar] [CrossRef]
- GA, M.; Brown, G. Determination of chromosome number of Kuwaiti flora I. Cytologia 1999, 64, 181–196. [Google Scholar]
- Brullo, S.; Pavone, P.; Terassi, M.C. Karyological considerations on the genus Plantago in Sicily Italy. Candollea 1985, 40, 217–230. [Google Scholar]
- Trivers, R.; Burt, A.; Palestis, B.G. B chromosomes and genome size in flowering plants. Genome 2004, 47, 1–8. [Google Scholar] [CrossRef]
- Jones, R.N.; Rees, H. B Chromosomes, 1st ed.; Academic Press: London, UK; New York, NY, USA, 1982. [Google Scholar]
- Bernardino, A.C.; Cabral-de-Mello, D.C.; Machado, C.B.; Palacios-Gimenez, O.M.; Santos, N.; Loreto, V.B. B chromosome variants of the grasshopper Xyleus discoideus angulatus are potentially derived from pericentromeric DNA. Cytogenet. Genome Res. 2017, 152, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Ziegler, C.G.; Steinlein, C.; Nanda, I.; Haaf, T. Chromosome banding in Amphibia: XXIV. The B chromosomes of Gastrotheca espeletia (Anura, Hylidae). Cytogenet. Genome Res. 2002, 97, 205. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.P.; Bougourd, S.M. Unstable B-chromosomes in a European population of Allium schoenoprasum L. (Liliaceae). Biol. J. Linn. Soc. 1994, 52, 357–363. [Google Scholar] [CrossRef]
- Houben, A.; Thompson, N.; Ahne, R.; Leach, C.R.; Verlin, D.; Timmis, J.N. A monophyletic origin of the B chromosomes of Brachycome dichromosomatica (Asteraceae). Plant Syst. Evol. 1999, 219, 127–135. [Google Scholar] [CrossRef]
- Belyayev, A.; Raskina, O. Chromosome evolution in marginal populations of Aegilops speltoides: Causes and consequences. Ann. Bot. 2013, 111, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.; Lamatsch, D.K.; Steinlein, C.; Engel, W.; Schartl, M.; Schmid, M. The giant B chromosome of the cyprinid fish Alburnus alburnus harbours a retrotransposon-derived repetitive DNA sequence. Chromosome Res. 2003, 11, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Duílio, M.D.A.; Pansonato-Alves, J.C.; Utsunomia, R.; Araya-Jaime, C.; Ruiz-Ruano, F.J.; Daniel, S.N.; Foresti, F. Delimiting the origin of a B chromosome by FISH mapping, chromosome painting and DNA sequence analysis in Astyanax paranae (Teleostei, Characiformes). PLoS ONE 2014, 9, e94896. [Google Scholar]
- Carter, C.R.; Smith-White, S. The cytology of Brachycome lineariloba. Chromosoma 1972, 39, 361–379. [Google Scholar] [CrossRef]
- Houben, A.; Verlin, D.; Leach, C.R.; Timmis, J.N. The genomic complexity of micro B chromosomes of Brachycome dichromosomatica. Chromosoma 2001, 110, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Kubaláková, M.; Valárik, M.; Barto, J.; Vrána, J.; Cihalikova, J.; Molnár-Láng, M.; Dolezel, J. Analysis and sorting of rye (Secale cereale L.) chromosomes using flow cytometry. Genome 2003, 46, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Martis, M.M.; Klemme, S.; Banaei-Moghaddam, A.M.; Blattner, F.R.; Macas, J.; Schmutzer, T.; Novák, P. Selfish supernumerary chromosome reveals its origin as a mosaic of host genome and organellar sequences. Proc. Natl. Acad. Sci. USA 2012, 109, 13343–13346. [Google Scholar] [CrossRef] [Green Version]
- Kumke, K.; Macas, J.; Fuchs, J.; Altschmied, L.; Kour, J.; Dhar, M.K.; Houben, A. Plantago lagopus B chromosome is enriched in 5S rDNA-derived satellite DNA. Cytogenet. Genome Res. 2016, 148, 68–73. [Google Scholar] [CrossRef]
- Theuri, J.; Phelps-Durr, T.; Mathews, S.; Birchler, J. A comparative study of retrotransposons in the centromeric regions of A and B chromosomes of maize. Cytogenet. Genome Res. 2005, 110, 203–208. [Google Scholar] [CrossRef]
- Lamb, J.C.; Riddle, N.C.; Cheng, Y.M.; Theuri, J.; Birchler, J.A. Localization and transcription of a retrotransposon-derived element on the maize B chromosome. Chromosome Res. 2007, 15, 383–398. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhu, H.M.; Huang, Q.F.; Zhao, L.; Zhang, G.J.; Roy, S.W.; Zhao, R.P. Deciphering neo-sex and B chromosome evolution by the draft genome of Drosophila albomicans. BMC Genom. 2012, 13, 109. [Google Scholar] [CrossRef]
- Houben, A.; Demidov, D.; Gernand, D.; Meister, A.; Leach, C.R.; Schubert, I. Methylation of histone H3 in euchromatin of plant chromosomes depends on basic nuclear DNA content. Plant J. 2003, 33, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Camacho, J.P.M. B chromosomes. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: San Diego, CA, USA, 2005; pp. 223–286. [Google Scholar]
- Burt, A.; Trivers, R. B chromosomes. In Genes in Conflict: The Biology of Selfish Genetic Elements; Harvard University Press: Cambridge, MA, USA, 2006; pp. 325–380. [Google Scholar]
- Bidau, C.J. A nucleolar-organizing B chromosome showing segregation-distortion in the grasshopper Dichroplus pratensis (Melanoplinae, Acrididae). Can. J. Genet. Cytol. 1986, 28, 138–148. [Google Scholar] [CrossRef]
- Leach, C.R.; Houben, A.; Field, B.; Pistrick, K.; Demidov, D.; Timmis, J.N. Molecular evidence for transcription of genes on a B chromosome in Crepis capillaris. Genetics 2005, 171, 269–278. [Google Scholar] [CrossRef]
- Ruiz-Estévez, M.; López-León, M.D.; Cabrero, J.; Camacho, J.P. Ribosomal DNA is active in different B chromosome variants of the grasshopper Eyprepocnemis plorans. Genetica 2013, 141, 337–345. [Google Scholar] [CrossRef]
- Ruiz-Estévez, M.; Badisco, L.; Broeck, J.V.; Perfectti, F.; Lopez-Leon, M.D.; Cabrero, J.; Camacho, J.P. B chromosomes showing active ribosomal RNA genes contribute insignificant amounts of rRNA in the grasshopper Eyprepocnemis plorans. Mol. Gen. Genom. 2014, 289, 1209–1216. [Google Scholar] [CrossRef]
- Carchilan, M.; Delgado, M.; Ribeiro, T.; Costa-Nunes, P.; Caperta, A.; Morais-Cecílio, L.; Jones, R.N.; Viegas, W.; Houben, A. Transcriptionally active heterochromatin in rye B chromosomes. Plant Cell 2007, 19, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Carchilan, M.; Kumke, K.; Mikolajewski, S.; Houben, A. Rye B chromosomes are weakly transcribed and might alter the transcriptional activity of a chromosome sequences. Chromosoma 2009, 118, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Klemme, S.; Banaei Moghaddam, A.M.; Macas, J.; Wicker, T.; Novák, P.; Houben, A. High copy sequences reveal distinct evolution of the rye B chromosome. New Phytol. 2013, 199, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Martis, M.M.; Macas, J.; Gundlach, H.; Himmelbach, A.; Altschmied, L.; Mayer, K.F.X.; Houben, A. Genes on B chromosomes: Old questions revisited with new tools. Biochim. Biophys. Acta 2015, 1849, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Ramos, É.; Cardoso, A.L.; Brown, J.; Marques, D.F.; Fantinatti, B.E.; Cabral-de-Mello, D.C.; Martins, C. The repetitive DNA element BncDNA, enriched in the B chromosome of the cichlid fish Astatotilapia latifasciata, transcribes a potentially noncoding RNA. Chromosoma 2016, 126, 313–323. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Kukekova, A.V.; Yudkin, D.V.; Trifonov, V.A.; Vorobieva, N.V.; Beklemisheva, V.R.; Perelman, P.L.; Graphodatskaya, D.A.; Trut, L.N.; Ferguson-Smith, M.A.; et al. The proto-oncogene c-KIT maps to canid B-chromosomes. Chromosome Res. 2005, 13, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Yudkin, D.V.; Trifonov, V.A.; Kukekova, A.V.; Vorobieva, N.V.; Rubtsova, N.V.; Yang, F.; Acland, G.M.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Mapping of KIT adjacent sequences on canid autosomes and B chromosomes. Cytogenet. Genome Res. 2007, 116, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Dementyeva, P.V.; Larkin, D.M.; O’Brien, P.C.; Perelman, P.L.; Yang, F.; Graphodatsky, A.S. Transcription of a protein-coding gene on B chromosomes of the Siberian roe deer (Capreolus pygargus). BMC Biol. 2013, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Meier, K.; Karimi-Ashtiyani, R.; Houben, A. Formation and expression of pseudogenes on the B chromosome of rye. Plant Cell 2013, 25, 2536–2544. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Gabriel, T.S.; Martis, M.M.; Gursinsky, T.; Schubert, V.; Vrána, J.; Doležel, J.; Grundlach, H.; Altschmied, L.; Scholz, U.; et al. Rye B chromosomes encode a functional Argonaute-like protein with in vitro slicer activities similar to it’s a chromosome paralog. New Phytol. 2016, 213, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Bougourd, S.M.; Jones, R.N. B chromosomes: A physiological enigma. New Phytol. 1997, 137, 43–54. [Google Scholar] [CrossRef]
- González-Sánchez, M.; González-González, E.; Molina, F.; Chiavarino, A.M.; Rosato, M.; Puertas, M.J. One gene determines maize B chromosome accumulation by preferential fertilisation; another gene(s) determines their meiotic loss. Heredity 2003, 90, 122. [Google Scholar] [CrossRef]
- Jackson, R.C.; Newmark, P. Effects of supernumerary chromosomes on production of pigment in Haplopappus gracilis. Science 1960, 132, 1316–1317. [Google Scholar] [CrossRef]
- Staub, R.W. Leaf striping correlated with the presence of B chromosomes in maize. J. Hered. 1987, 78, 71–74. [Google Scholar] [CrossRef]
- Holmes, D.S.; Bougourd, S.M. B-chromosome selection in Allium schoenoprasum II. Experimental populations. Heredity 1991, 67, 117. [Google Scholar] [CrossRef]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Okada, N. B chromosomes have a functional effect on female sex determination in Lake Victoria cichlid fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef]
- Paliwal, R.L.; Hyde, B.B. The association of a single B-chromosome with male sterility in Plantago coronopus. Am. J. Bot. 1959, 46, 460–466. [Google Scholar] [CrossRef]
- Harvey, A.W.; Hewitt, G.M. B chromosomes slow development in a grasshopper. Heredity 1979, 42, 397. [Google Scholar] [CrossRef]
- Hewitt, G.M.; Butlin, R.K.; East, T.M. Testicular dysfunction in hybrids between parapatric subspecies of the grasshopper Chorthippus parallelus. Biol. J. Linn. Soc. 1987, 31, 25–34. [Google Scholar] [CrossRef]
- Dover, G.A.; Riley, R. Variation at two loci affecting homoeologous meiotic chromosome pairing in Triticum aestivum × Aegilops mutica hybrids. Nat. New Biol. 1972, 235, 61. [Google Scholar] [CrossRef]
- Kousaka, R.; Endo, T.R. Effect of a rye B chromosome and its segments on homoeologous pairing in hybrids between common wheat and Aegilops variabilis. Genes Genet. Syst. 2012, 87, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dherawattana, A.; Sadanaga, K. Cytogenetics of a crown rust-resistant hexaploid oat with 42 + 2 fragment chromosomes 1. Crop Sci. 1973, 13, 591–594. [Google Scholar] [CrossRef]
- Werren, J.H. The paternal-sex-ratio chromosome of Nasonia. Am. Nat. 1991, 137, 392–402. [Google Scholar] [CrossRef]
- Cheng, Z.K.; Yu, H.X.; Yan, H.H.; Gu, M.H.; Zhu, L.H. B chromosome in a rice aneuploid variation. Theor Appl. Genet. 2000, 101, 564–568. [Google Scholar] [CrossRef]
- Zima, J.; Piálek, J.; Macholán, M. Possible heterotic effects of B chromosomes on body mass in a population of Apodemus flavicollis. Can. J. Zool. 2003, 81, 1312–1317. [Google Scholar] [CrossRef]
- Jenkins, G.; Jones, R.N. B chromosomes in hybrids of temperate cereals and grasses. Cytogenet. Genome Res. 2004, 106, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Larracuente, A.M.; Presgraves, D.C. The selfish Segregation Distorter gene complex of Drosophila melanogaster. Genetics 2012, 192, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Dhar, M.K.; Kour, G.; Kaul, S. B chromosome in Plantago lagopus Linnaeus, 1753 shows preferential transmission and accumulation through unusual processes. Comp. Cytogenet. 2017, 11, 375. [Google Scholar] [CrossRef]
- Camacho, J.P.M.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. B 2000, 355, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.H.; Albini, S.M.; Whitehorn, J.A.F. Ultrastructure of meiotic pairing in B chromosomes of Crepis capillaris. Chromosoma 1991, 100, 193–202. [Google Scholar] [CrossRef]
- Manrique-Poyato, M.I.; López-León, M.D.; Gómez, R.; Perfectti, F.; Camacho, J.P.M. Population genetic structure of the grasshopper Eyprepocnemis plorans in the south and east of the Iberian Peninsula. PLoS ONE 2013, 8, e59041. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Schubert, V.; Kumke, K.; Weiβ, O.; Klemme, S.; Nagaki, K.; Macas, J.; Gonzalez-Sanchez, M.; Heredia, V.; Gomez-Revilla, D.; et al. Nondisjunction in favor of a chromosome: The mechanism of rye B chromosome drive during pollen mitosis. Plant Cell 2012, 24, 4124–4134. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.R.; Nasuda, S.; Jones, N.; Dou, Q.; Akahori, A.; Wakimoto, M.; Tsujimoto, H. Dissection of rye B chromosomes, and nondisjunction properties of the dissected segments in a common wheat background. Genes Genet. Syst. 2008, 83, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, T.; Schramke, V.; Hamilton, G.L.; White, S.A.; Teng, G.; Martienssen, R.A.; Allshire, R.C. RNA interference is required for normal centromere function in fission yeast. Chromosome Res. 2003, 11, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Vos, L.J.; Famulski, J.K.; Chan, G.K. How to build a centromere: From centromeric and pericentromeric chromatin to kinetochore assembly. Biochem. Cell Biol. 2006, 84, 619–639. [Google Scholar] [CrossRef] [PubMed]
- Bauerly, E.; Hughes, S.E.; Vietti, D.R.; Miller, D.E.; McDowell, W.; Hawley, R.S. Discovery of supernumerary B chromosomes in Drosophila melanogaster. Genetics 2014, 113, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Kichigin, I.G.; Larkin, D.M.; O’Brien, P.C.; Ferguson-Smith, M.A.; Yang, F.; Graphodatsky, A.S. Contrasting origin of B chromosomes in two cervids (Siberian roe deer and grey brocket deer) unravelled by chromosome-specific DNA sequencing. BMC Genom. 2016, 17, 618. [Google Scholar] [CrossRef]
- Houben, A.; Carchilan, M. Plant B chromosomes: What makes them different? In Plant Cytogenetics; Springer: New York, NY, USA, 2012; pp. 59–77. [Google Scholar]
- Jones, R.N.; Houben, A. B chromosomes in plants: Escapees from the A chromosome genome? Trends Plant Sci. 2003, 8, 417–442. [Google Scholar] [CrossRef]
- Bidau, C.J. Influence of a rare unstable B-chromosome on chiasma frequency and nonhaploid sperm production in Dichroplus pratensis (Melanoplinae, Acrididae). Genetica 1987, 73, 201–210. [Google Scholar] [CrossRef]
- Burt, A.; Trivers, R. Selfish DNA and breeding system in flowering plants. Proc. R. Soc. Lond. 1998, 265, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Muntzing, A. Cyto-genetics of accessory chromosomes (B-chromosomes). Caryologia 1954, 6, 282–301. [Google Scholar]
- Puertas, M.J.; Ramirez, A.; Baeza, F. The transmission of B chromosomes in Secale cereale and Secale vavilovii populations. II. Dynamics of populations. Heredity 1987, 58, 81. [Google Scholar] [CrossRef]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Krohne, D.T.; Baker, I.; Baker, H.G. The maintenance of the gynodioecious breeding system in Plantago lanceolata L. Am. Midl. Nat. 1980, 103, 269–279. [Google Scholar] [CrossRef]
- Charlesworth, B.; Charlesworth, D. A model for the evolution of dioecy and gynodioecy. Am. Nat. 1978, 112, 975–997. [Google Scholar] [CrossRef]
- Marques, A.; Klemme, S.; Houben, A. Evolution of plant B chromosome enriched sequences. Genes 2018, 9, 515. [Google Scholar] [CrossRef]
- Hosid, E.; Brodsky, L.; Kalendar, R.; Raskina, O.; Belyayev, A. Diversity of long terminal repeat retrotransposon genome distribution in natural populations of the wild diploid wheat Aegilops speltoides. Genetics 2012, 190, 263–274. [Google Scholar] [CrossRef]
- Berdnikov, V.A.; Gorel, F.L.; Kosterin, O.E.; Bogdanova, V.S. Tertiary trisomics in the garden pea as a model of B chromosome evolution in plants. Heredity 2003, 91, 577. [Google Scholar] [CrossRef]
- Rimpau, J.; Flavell, R.B. Characterisation of rye B chromosome DNA by DNA/DNA hybridisation. Chromosoma 1975, 52, 207–217. [Google Scholar] [CrossRef]
- Timmis, K.; Cabello, F.; Cohen, S.N. Cloning, isolation, and characterization of replication regions of complex plasmid genomes. Proc. Natl. Acad. Sci. USA 1975, 72, 2242–2246. [Google Scholar] [CrossRef]
- Sandery, M.J.; Forster, J.W.; Blunden, R.; Jones, R.N. Identification of a family of repeated sequences on the rye B chromosome. Genome 1990, 33, 908–913. [Google Scholar] [CrossRef]
- Wilkes, T.M.; Francki, M.G.; Langridge, P.; Karp, A.; Jones, R.N.; Forster, J.W. Analysis of rye B-chromosome structure using fluorescence in situ hybridization (FISH). Chromosome Res. 1995, 3, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, T.V.; Andreenkova, O.V.; Bochkaerev, M.N.; Borissov, Y.M.; Bogdanchikova, N.; Borodin, P.M.; Rubtsov, N.B. B chromosomes of Korean field mouse Apodemus peninsulae (Rodentia, Murinae) analysed by microdissection and FISH. Cytogenet. Genome Res. 2002, 96, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.M.; Lin, B.Y. Molecular organization of large fragments in the maize B chromosome: Indication of a novel repeat. Genetics 2004, 166, 1947–1961. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Banaei-Moghaddam, A.M.; Klemme, S.; Blattner, F.R.; Niwa, K.; Guerra, M.; Houben, A. B chromosomes of rye are highly conserved and accompanied the development of early agriculture. Ann. Bot. 2013, 112, 527–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.E.D.; Thomas, R.; Trifonov, V.A.; Wayne, R.K.; Graphodatsky, A.S.; Breen, M. Anchoring the dog to its relatives reveals new evolutionary breakpoints across 11 species of the Canidae and provides new clues for the role of B chromosomes. Chromosome Res. 2011, 19, 685. [Google Scholar] [CrossRef]
- Shirasu, K.; Schulman, A.H.; Lahaye, T.; Schulze-Lefert, P. A contiguous 66-kb barley DNA sequence provides evidence for reversible genome expansion. Genome Res. 2000, 10, 908–915. [Google Scholar] [CrossRef]
- Coleman, J.J.; Rounsley, S.D.; Rodriguez-Carres, M.; Kuo, A.; Wasmann, C.C.; Grimwood, J.; Schwartz, D.C. The genome of Nectria haematococca: Contribution of supernumerary chromosomes to gene expansion. PLoS Genet. 2009, 5, e1000618. [Google Scholar] [CrossRef]
- Cohen, S.; Segal, D. Extrachromosomal circular DNA in eukaryotes: Possible involvement in the plasticity of tandem repeats. Cytogenet. Genome Res. 2009, 124, 327–338. [Google Scholar] [CrossRef]
- Cohen, S.; Houben, A.; Segal, D. Extrachromosomal circular DNA derived from tandemly repeated genomic sequences in plants. Plant J. 2008, 53, 1027–1034. [Google Scholar] [CrossRef]
- Maluszynska, J.; Schweizer, D. Ribosomal RNA genes in B chromosomes of Crepis capillaris detected by non-radioactive in situ hybridization. Heredity 1989, 62, 59. [Google Scholar] [CrossRef] [PubMed]
- Stitou, S.; de La Guardia, R.D.; Jiménez, R.; Burgos, M. Inactive ribosomal cistrons are spread throughout the B chromosomes of Rattus rattus (Rodentia, Muridae). Implications for their origin and evolution. Chromosome Res. 2000, 8, 305–311. [Google Scholar] [CrossRef]
- Marschner, S.; Kumke, K.; Houben, A. B chromosomes of B. dichromosomatica show a reduced level of euchromatic histone H3 methylation marks. Chromosome Res. 2007, 15, 215. [Google Scholar] [CrossRef] [PubMed]
- Bass, H.; Birchler, J.A. Plant Cytogenetics: Genome Structure and Chromosome Function; Springer Science & Business Media: New York, NY, USA, 2011; Volume 4. [Google Scholar]
- Battaglia, E. Cytogenetics of B-chromosomes. Caryologia 1964, 17, 245–299. [Google Scholar] [CrossRef]
- Gernand, D.; Rutten, T.; Varshney, A.; Rubtsova, M.; Prodanovic, S.; Brüß, C.; Houben, A. Uniparental chromosome elimination at mitosis and interphase in wheat and pearl millet crosses involves micronucleus formation, progressive heterochromatinization, and DNA fragmentation. Plant Cell 2005, 17, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, W.; Friebe, B.; Gill, B.S. The origin of a “zebra” chromosome in wheat suggests nonhomologous recombination as a novel mechanism for new chromosome evolution and step changes in chromosome number. Genetics 2008, 179, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Teruel, M.; Cabrero, J.; Montiel, E.E.; Acosta, M.J.; Sánchez, A.; Camacho, J.P.M. Microdissection and chromosome painting of X and B chromosomes in Locusta migratoria. Chromosome Res. 2009, 17, 11. [Google Scholar] [CrossRef]
- Rajicic, M.; Romanenko, S.A.; Karamysheva, T.V.; Blagojević, J.; Adnađević, T.; Budinski, I.; Vujošević, M. The origin of B chromosomes in yellow-necked mice (Apodemus flavicollis)—Break rules but keep playing the game. PLoS ONE 2017, 12, e0172704. [Google Scholar] [CrossRef]
- Green, D.M. Structure and evolution of B chromosomes in amphibians. Cytogenet. Genome Res. 2004, 106, 235–242. [Google Scholar] [CrossRef]
- Cioffi, M.D.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA; Karger Publishers: Basel, Switzerland, 2012; Volume 7, pp. 197–221. [Google Scholar]
- Berend, S.A.; Hale, D.W.; Engstrom, M.D.; Greenbaum, I.F. Cytogenetics of collared lemmings (Dicrostonyx groenlandicus). II. Meiotic behavior of B chromosomes suggests a Y-chromosome origin of supernumerary chromosomes. Cytogenet. Genome Res. 2001, 95, 85–91. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhar, M.K.; Kour, J.; Kaul, S. Origin, Behaviour, and Transmission of B Chromosome with Special Reference to Plantago lagopus. Genes 2019, 10, 152. https://doi.org/10.3390/genes10020152
Dhar MK, Kour J, Kaul S. Origin, Behaviour, and Transmission of B Chromosome with Special Reference to Plantago lagopus. Genes. 2019; 10(2):152. https://doi.org/10.3390/genes10020152
Chicago/Turabian StyleDhar, Manoj K., Jasmeet Kour, and Sanjana Kaul. 2019. "Origin, Behaviour, and Transmission of B Chromosome with Special Reference to Plantago lagopus" Genes 10, no. 2: 152. https://doi.org/10.3390/genes10020152