Real-Time Quantitative PCR Analysis of the Expression Pattern of the Hypoglycemic Polypeptide-P Gene in Momordica charantia

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Protein Isolation
2.4. qPCR Primer Design
2.5. qPCR Analysis
2.6. WB Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. qPCR Primer of MC β-actin
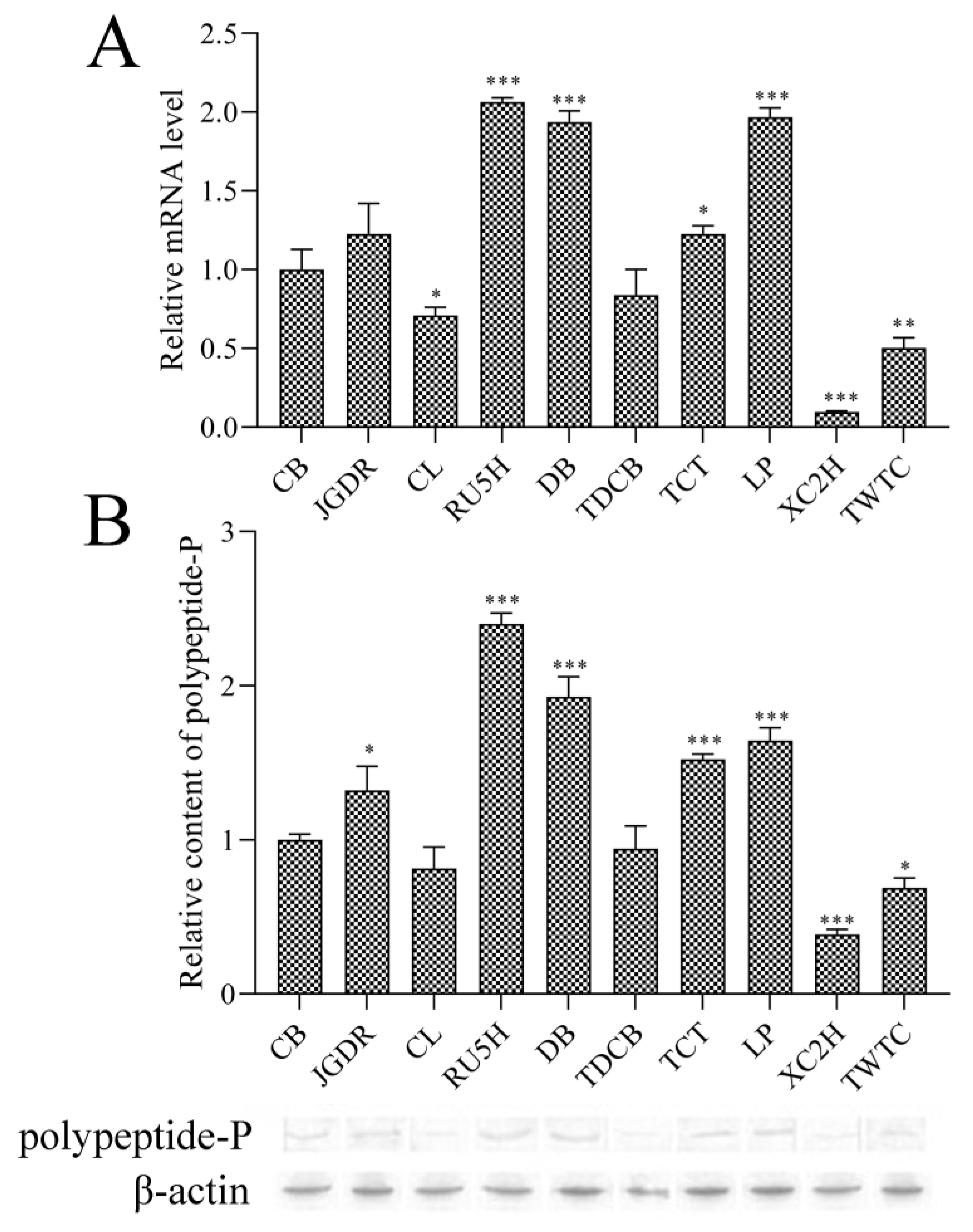
3.2. Expression Analysis of the Polypeptide-P Gene in Different Accessions
3.3. Expression Analysis of the Polypeptide-P Gene during Different Harvesting Seasons
3.4. Expression Analysis of the Polypeptide-P Gene in Different Tissues
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Patel, D.K.; Kumar, R.; Prasad, S.K.; Sairam, K.; Hemalatha, S. Antidiabetic and in vitro antioxidant potential of Hybanthus enneaspermus (Linn) F. Muell in streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2011, 1, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Laitonjam, W. Traditional medicinal plants of Manipur as anti-diabetics. J. Med. Plants Res. 2011, 5, 677–687. [Google Scholar]
- Chauhan, A.; Sharma, P.K.; Srivastava, P.; Kumar, N.; Dudhe, R. Plants Having Potential Antidiabetic Activity: A Review. Der Pharm. Lett. 2010, 2, 369–387. [Google Scholar]
- Pareek, H.; Sharma, S.; Khajja, B.S.; Jain, K.; Jain, G.C. Evaluation of hypoglycemic and anti-hyperglycemic potential of Tridax procumbens (Linn). BMC Complement. Altern. Med. 2009, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meenakshi, P.; Bhuvaneshwari, R.; Rathi, M.A.; Thirumoorthi, L.; Guravaiah, D.C.; Jiji, M.J.; Gopalakrishnan, V.K. Antidiabetic activity of ethanolic extract of Zaleya decandra in alloxan-induced diabetic rats. Appl. Biochem. Biotechnol. 2010, 162, 1153–1159. [Google Scholar] [CrossRef]
- Subratty, A.H.; Gurib Fakim, A.; Mahomoodally, F. Bitter melon: An exotic vegetable with medicinal values. Nutr. Food Sci. 2005, 35, 143–147. [Google Scholar] [CrossRef]
- Rajasekhar, M.D.; Babu, K.R.; Vinay, K.; Sampath, M.R.; Sameena, S.K.; Apparao, C.H. Antihyperglycemic and antioxidant activities of active fraction from the aqueous extract of Momordica cymbalaria fruits in Streptozotocin induced diabetic rats. Pharmacogn. Res. 2009, 1, 352. [Google Scholar]
- Tongia, A.; Tongia, S.K.; Dave, M. Phytochemical determination and extraction of Momordica charantia fruit and its hypoglycemic potentiation of oral hypoglycemic drugs in diabetes mellitus (NIDDM). Indian J. Physiol. Pharmacol. 2004, 48, 241–244. [Google Scholar]
- Grover, J.K.; Yadav, S.P. Pharmacological actions and potential uses of Momordica charantia: A review. J. Ethnopharmacol. 2004, 93, 123–132. [Google Scholar] [CrossRef]
- Miura, T.; Itoh, C.; Iwamoto, N.; Kato, M.; Kawai, M.; Park, S.R.; Suzuki, I. Hypoglycemic activity of the fruit of the Momordica charantia in type 2 diabetic mice. J. Nutr. Sci. Vitaminol. 2001, 47, 340–344. [Google Scholar] [CrossRef] [Green Version]
- Virdi, J.; Sivakami, S.; Shahani, S.; Suthar, A.C.; Banavalikar, M.M.; Biyani, M.K. Antihyperglycemic effects of three extracts from Momordica charantia. J. Ethnopharmacol. 2003, 88, 107–111. [Google Scholar] [CrossRef]
- Ahmad, Z.; Zamhuri, K.F.; Yaacob, A.; Siong, C.H.; Selvarajah, M.; Ismail, A.; Nazrul Hakim, M. In Vitro anti-diabetic activities and chemical analysis of polypeptide-k and oil isolated from seeds of Momordica charantia (bitter gourd). Molecules 2012, 17, 9631–9640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwa, V.S.; Bhandari, C.M.; Pangaria, A.; Goyal, R.K. Clinical trial in patients with diabetes mellitus of an insulin-like compound obtained from plant source. Upsala J. Med. Sci. 1977, 82, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Khanna, P.; Jain, S.C.; Panagariya, A.; Dixit, V.P. Hypoglycemic activity of polypeptide-p from a plant source. J. Nat. Prod. 1981, 44, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Jini, D. Antidiabetic effects of Momordica charantia (bitter melon) and its medicinal potency. Asian Pac. J. Trop. Dis. 2013, 3, 93–102. [Google Scholar] [CrossRef]
- Raman, A.; Lau, C. Anti-diabetic properties and phytochemistry of Momordica charantia L. (Cucurbitaceae). Phytomedicine 1996, 2, 349–362. [Google Scholar] [CrossRef]
- Ali, L.; Khan, A.K.; Mamun, M.I.; Mosihuzzaman, M.; Nahar, N.; Nur-e-Alam, M.; Rokeya, B. Studies on hypoglycemic effects of fruit pulp, seed, and whole plant of Momordica charantia on normal and diabetic model rats. Planta Med. 1993, 59, 408–412. [Google Scholar] [CrossRef]
- Tian, M.; Zeng, X.Q.; Song, H.L.; Hu, S.X.; Wang, F.J.; Zhao, J.; Hu, Z.B. Molecular diversity and hypoglycemic polypeptide-P content of Momordica charantia in different accessions and different seasons. J. Sci. Food Agric. 2015, 95, 1328–1335. [Google Scholar] [CrossRef]
- Yuan, X.; Gu, X.; Tang, J. Purification and characterisation of a hypoglycemic peptide from Momordica Charantia, L. Var. abbreviata Ser. Food Chem. 2008, 111, 415–420. [Google Scholar] [CrossRef]
- Zhang, F.; Niu, B.; Wang, Y.; Chen, F.; Wang, S.; Xu, Y.; Jiang, L.; Gao, S.; Wu, J.; Tang, L. A novel betaine aldehyde dehydrogenase gene from Jatropha curcas, encoding an enzyme implicated in adaptation to environmental stress. Plant Sci. 2008, 174, 510–518. [Google Scholar] [CrossRef]
- Xu, W.F.; Shi, W.M. Expression profiling of the 14-3-3 gene family in response to salt stress and potassium and iron deficiencies in young tomato (Solanum lycopersicum) roots: Analysis by real-time RT-PCR. Ann. Bot. 2006, 98, 965–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, J.H.; Clough, S.J.; Chan, W.C.; Vodkin, L.O. Tissue-specific gene silencing mediated by a naturally occurring chalcone synthase gene cluster in Glycine max. Plant Cell 2004, 16, 819–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, A.S.; Scaboo, A.M.; Wu, X.; Nguyen, H.T.; Pantalone, V.R.; Bilyeu, K.D. Characterization of the MIPS gene family in Glycine max. Plant Breed. 2006, 125, 493–500. [Google Scholar] [CrossRef]
- Wang, B.L.; Zhang, W.J.; Zhao, J.; Wang, F.J.; Fan, L.Q.; Wu, Y.X.; Hu, Z.B. Gene cloning and expression of a novel hypoglycaemic peptide from Momordica charantia. J. Sci. Food Agric. 2011, 91, 2443–2448. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Pirttilä, A.M.; Halonen, M.; Hohtola, A. Isolation of high quality RNA from bilberry (Vaccinium myrtillus L.) fruit. Mol. Biotechnol. 2001, 19, 201–203. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No. | Accession | Origin a | Fruit Character b |
|---|---|---|---|
| 1 | DB | Hunan | White, medium long, continuous ridges |
| 2 | CB | Hunan | Light green, medium-long c, continuous ridges |
| 3 | LP | Hunan | Green, long d, discontinuous ridges |
| 4 | RU5H | Fujian | Glossy green, medium-long, discontinuous ridges |
| 5 | TDCB | Hunan | Greenish white, extra-long e, discontinuous ridges |
| 6 | TWTC | Taiwan | Glossy green, extra-long, discontinuous ridges |
| 7 | CL | Guangdong | Light green, long, continuous ridges |
| 8 | XC2H | Fujian | Green, medium-long, continuous ridges |
| 9 | JGDR | Changsha | Glossy green, medium-long, discontinuous ridges |
| 10 | TCT | Guangdong | Glossy green, long, discontinuous ridges |
| Gene | Primer Name | Amplified Length | Primer Sequence (5′–3′) |
|---|---|---|---|
| MC β-actin | MC-actin-F | 124bp | CCCTCCCTCATGCAATTCTC |
| MC-actin-R | TCGGCAGTAGTGGTAAACATGTAAC | ||
| polypeptide-P | P-F | 148bp | CGTGATGAAGGCAAAGTGGA |
| P-R | TATCGCCAAACGGGGTAATG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-S.; Zeng, X.-Q.; Yang, X.-Z.; Liu, W.; Li, P.-F.; Wang, F.-J.; Zhao, J. Real-Time Quantitative PCR Analysis of the Expression Pattern of the Hypoglycemic Polypeptide-P Gene in Momordica charantia. Genes 2019, 10, 1044. https://doi.org/10.3390/genes10121044
Wang Y-S, Zeng X-Q, Yang X-Z, Liu W, Li P-F, Wang F-J, Zhao J. Real-Time Quantitative PCR Analysis of the Expression Pattern of the Hypoglycemic Polypeptide-P Gene in Momordica charantia. Genes. 2019; 10(12):1044. https://doi.org/10.3390/genes10121044
Chicago/Turabian StyleWang, Yi-Shuai, Xiang-Qing Zeng, Xu-Zhong Yang, Wei Liu, Peng-Fei Li, Fu-Jun Wang, and Jian Zhao. 2019. "Real-Time Quantitative PCR Analysis of the Expression Pattern of the Hypoglycemic Polypeptide-P Gene in Momordica charantia" Genes 10, no. 12: 1044. https://doi.org/10.3390/genes10121044