Labeling of Monilinia fructicola with GFP and Its Validation for Studies on Host-Pathogen Interactions in Stone and Pome Fruit

 , , , ,
, , , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fungi and Bacteria Growth Conditions
2.2. Agrobacterium tumefaciens-Mediated M. fructicola Transformation
2.3. Molecular Analyses of M. fructicola Transformants
2.4. Characterization of GFP-M. fructicola Transformants
2.5. Microscopy Analysis of M. fructicola GFP-Transformants
3. Results
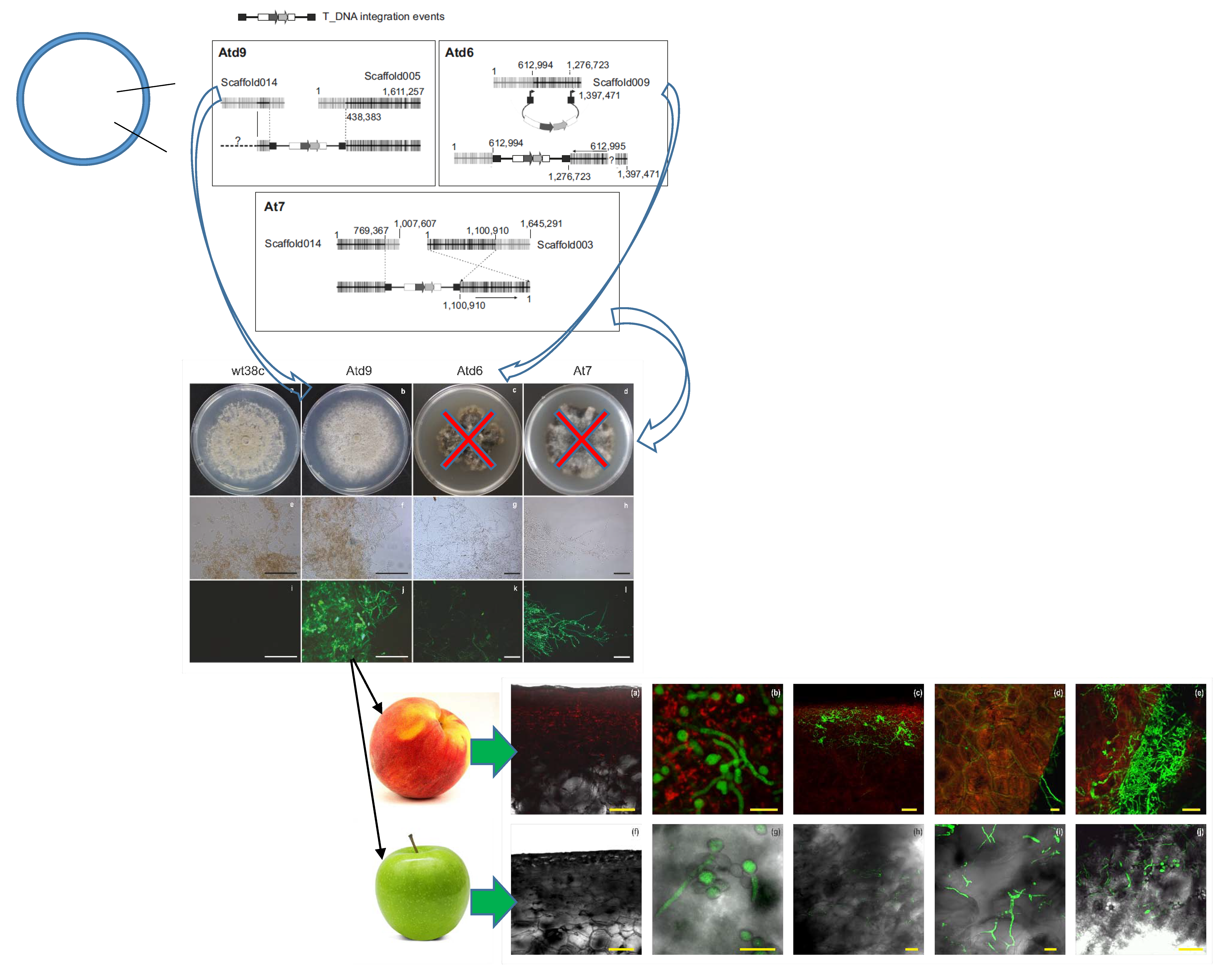
3.1. Generating Green-Fluorescent M. fructicola Strains by Transformation
3.2. Characterization of GFP-M. fructicola Transformants
3.3. Microscopy Analysis of M. fructicola GFP-Transformants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Byrde, R.J.W.; Willetts, H.J. The Brown Rot Fungi of Fruit: Their Biology and Control; Pergamon Press Ltd.: Oxford, UK, 1977; p. 188. [Google Scholar]
- Vasić, M.; Vico, I.; Jurick, W.M.; Duduk, N. Distribution and characterization of Monilinia spp. causing apple fruit decay in Serbia. Plant Dis. 2018, 102, 359–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cal, A.; Gell, I.; Usall, J.; Viñas, I.; Melgarejo, P. First report of brown rot caused by Monilinia fructicola in peach orchards in Ebro Valley, Spain. Plant Dis. 2009, 93, 763. [Google Scholar] [CrossRef] [PubMed]
- Villarino, M.; Egüen, B.; Lamarca, N.; Segarra, J.; Usall, J.; Melgarejo, P.; De Cal, A. Occurrence of Monilinia laxa and M. fructigena after introduction of M. fructicola in peach orchards in Spain. Eur. J. Plant Pathol. 2013, 137, 835–845. [Google Scholar] [CrossRef]
- Villarino, M.; Melgarejo, P.; De Cal, A. Growth and aggressiveness factors affecting Monilinia spp. survival peaches. Inter. J. Food Microbiol. 2016, 227, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Benitez, C.; Melgarejo, P.; De Cal, A.; Fontaniella, B. Microscopic analyses of latent and visible Monilinia fructicola infections in nectarines. PLoS ONE 2016, 11, e0160675. [Google Scholar] [CrossRef] [Green Version]
- Washington, W.S.; Pascoe, I. First record of Monilinia fructicola on strawberry fruit in Victoria, Australia. Australas. Plant Pathol. 2000, 29, 70. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Nichols, B.; Takeda, F.; Jurick, W.M. First report of Monilinia fructicola causing postharvest decay of strawberries (Fragaria × ananassa) in the United States. Plant Dis. 2017, 101, 1823. [Google Scholar] [CrossRef]
- Bevacqua, D.; Quilot-Turion, B.; Bolzoni, L. A model for temporal dynamics of brown rot spreading in fruit orchards. Phytopathology 2018, 108, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Duchoslavová, J.; Širučková, I.; Zapletalová, E.; Navrátil, M.; Šafářová, D. First report of brown rot caused by Monilinia fructicola on various stone and pome fruits in the Czech Republic. Plant Dis. 2007, 91, 907. [Google Scholar] [CrossRef]
- Grabke, A.; Hu, M.J.; Luo, C.X.; Bryson, P.K.; Schnabel, G. First report of brown rot of apple caused by Monilinia fructicola in Germany. Plant Dis. 2011, 95, 772. [Google Scholar] [CrossRef]
- Vasić, M.; Duduk, N.; Ivanović, M.M.; Obradović, A.; Ivanović, M.S. First report of brown rot caused by Monilinia fructicola on stored apple in Serbia. Plant Dis. 2012, 96, 456. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.; Di Francesco, A.; Lantos, A.; Mari, M. First report of asiatic brown rot (Monilinia polystroma) and brown rot (Monilinia fructicola) on pears in Italy. Plant Dis. 2015, 99, 556. [Google Scholar] [CrossRef]
- Peter, K.A.; Gaskins, V.L.; Lehman, B.; Jurick, W.M. First report of brown rot on apple fruit caused by Monilinia fructicola in Pennsylvania. Plant Dis. 2015, 99, 1179. [Google Scholar] [CrossRef]
- Zhu, X.Q.; Niu, C.W.; Chen, X.Y.; Guo, L.Y. Monilinia species associated with brown rot of cultivated apple and pear fruit in China. Plant Dis. 2016, 100, 2240–2250. [Google Scholar] [CrossRef] [Green Version]
- Pereira, W.V.; May-De-Mio, L.L. First report of brown rot caused by Monilinia fructicola on apple in Brazil. Plant Dis. 2018, 102, 2657. [Google Scholar] [CrossRef]
- Horowitz, S.; Freeman, S.; Sharon, A. Use of green fluorescent protein-transgenic strains to study pathogenic and nonpathogenic lifestyles in Colletotrichum acutatum. Phytopathology 2002, 92, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cruz, J.; Romero, D.; de Vicente, A.; Pérez-García, A. Transformation of the cucurbit powdery mildew pathogen Podosphaera xanthii by Agrobacterium tumefaciens. New Phytol. 2017, 213, 1961–1973. [Google Scholar]
- Isshiki, A.; Ohtani, K.; Kyo, M.; Yamamoto, H.; Akimitsu, K. Green fluorescent detection of fungal colonization and endopolygalacturonase gene expression in the interaction of Alternaria citri with citrus. Phytopathology 2003, 93, 768–773. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Zhou, T.; Yu, H. Transformation of Botrytis cinerea with a Green Fluorescent Protein (GFP) Gene for the Study of Host-pathogen Interactions. Plant Pathol. J. 2006, 6, 134–140. [Google Scholar]
- De Silva, A.P.; Bolton, M.D.; Nelson, B.D. Transformation of Sclerotinia sclerotiorum with the green fluorescent protein gene and fluorescence of hyphae in four inoculated hosts. Plant Pathol. 2009, 58, 487–496. [Google Scholar] [CrossRef]
- Bansal, K.; Rollins, J. Exploring the Monilinia vaccinii-corymbosi-blueberry pathosystem for the discovery of genes governing host-specificity. Phytopathology 2016, 106, 152. [Google Scholar]
- Egüen, B.; Melgarejo, P.; De Cal, A. Sensitivity of Monilinia fructicola from Spanish peach orchards to thiophanate-methyl, iprodione, and cyproconazole: Fitness analysis and competitiveness. Eur. J. Plant Pathol. 2015, 141, 789–801. [Google Scholar] [CrossRef]
- Michielse, C.B.; van Wijk, R.; Reijnen, L.; Cornelissen, B.J.; Rep, M. Insight into the molecular requirements for pathogenicity of Fusarium oxysporum f. sp. lycopersici through large-scale insertional mutagenesis. Genome Biol. 2009, 10, R4. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; Hooykaas, P.J.; van den Hondel, C.A.; Ram, A.F. Agrobacterium-mediated transformation of the filamentous fungus Aspergillus awamori. Nat. Protoc. 2008, 3, 1671–1678. [Google Scholar] [CrossRef]
- Hernández-Ortiz, P.; Espeso, E.A. Phospho-regulation and nucleocytoplasmic trafficking of CrzA in response to calcium and alkaline-pH stress in Aspergillus nidulans. Mol. Microbiol. 2013, 89, 532–551. [Google Scholar] [CrossRef]
- Meirinho, S.; Carvalho, M.; Dominguez, Á.; Choupina, A. Isolation and characterization by asymmetric PCR of the ENDO1 gene for glucan endo-1,3-β-D-glucosidase in Phytophthora cinnamomi associated with the ink disease of Castanea sativa Mill. Braz. Arch. Biol. Technol. 2010, 53, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Hoff, K.J.; Stanke, M. WebAUGUSTUS—A web service for training AUGUSTUS and predicting genes in eukaryotes. Nucleic Acids Res. 2013, 41, W123–W128. [Google Scholar] [CrossRef]
- Sauer, D.B.; Burroughs, R. Disinfection of seed surfaces with sodium hypochlorite. Phytopathology 1986, 76, 745–749. [Google Scholar] [CrossRef]
- Marcel, S.; Sawers, R.; Oakeley, E.; Angliker, H.; Paszkowski, U. Tissue-adapted invasion strategies of the rice blast fungus Magnaporthe oryzae. Plant Cell 2010, 22, 3177–3187. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.; Kim, D.W.; Park, J.S.; Khang, C.H. Live-cell fluorescence imaging to investigate the dynamics of plant cell death during infection by the rice blast fungus Magnaporthe oryzae. BMC Plant. Biol. 2016, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Hardham, A.R. Confocal Microscopy in Plant–Pathogen Interactions. In Plant Fungal Pathogens. Methods in Molecular Biology (Methods and Protocols); Bolton, M.T.B., Ed.; Humana Press: Totowa, NJ, USA, 2012; pp. 295–309. [Google Scholar]
- Buron-Moles, G.; López-Pérez, M.; González-Candelas, L.; Viñas, I.; Teixidó, N.; Usall, J.; Torres, R. Use of GFP-tagged strains of Penicillium digitatum and Penicillium expansum to study host-pathogen interactions in oranges and apples. Inter. J. Food Microbiol. 2012, 160, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagopodi, A.L.; Ram, A.F.; Lamers, G.E.; Punt, P.J.; Van den Hondel, C.A.; Lugtenberg, B.J.; Bloemberg, G.V. Novel aspects of tomato root colonization and infection by Fusarium oxysporum f. sp. radicis-lycopersici revealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker. Mol. Plant Microbe Interact. 2002, 15, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, T.X.; Ngo, T.T.; Mai, L.T.D.; Bui, T.T.; Le, D.H.; Bui, H.T.V.; Nguyen, H.Q.; Ngo, B.X.; Tran, V.T. A highly efficient Agrobacterium tumefaciens-mediated transformation system for the postharvest pathogen Penicillium digitatum using DsRed and GFP to visualize citrus host colonization. J. Microbiol. Methods. 2018, 144, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Gril, T.; Celar, F.; Munda, A.; Javornik, B.; Jakse, J. AFLP analysis of intraspecific variation between Monilinia laxa isolates from different hosts. Plant Dis. 2008, 92, 1616–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Yuan, N.; Schnabel, G.; Luo, C. Function of the genetic element “Mona” associated with fungicide resistance in Monilinia fructicola. Mol. Plant Pathol. 2017, 18, 90–97. [Google Scholar] [CrossRef]
- Chou, C.M.; Yu, F.Y.; Yu, P.L.; Ho, J.F.; Bostock, R.M.; Chung, K.R.; Huang, J.-W.; Lee, M.H. Expression of five endopolygalacturonase genes and demonstration that MfPG1 overexpression diminishes virulence in the brown rot pathogen Monilinia fructicola. PLoS ONE 2015, 10, e0132012. [Google Scholar] [CrossRef]
- Lee, M.H.; Bostock, R.M. Agrobacterium T-DNA-mediated integration and gene replacement in the brown rot pathogen Monilinia fructicola. Curr. Genet. 2006, 49, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Chiu, C.M.; Roubtsova, T.; Chou, C.M.; Bostock, R.M. Overexpression of a redox-regulated cutinase gene, MfCUT1, increases virulence of the brown rot pathogen Monilinia fructicola on Prunus spp. Mol. Plant Microbe Interact. 2010, 23, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Sun, Z.; Schnabel, G. Development of spontaneous hygromycin B resistance in Monilinia fructicola and Its impact on growth rate, morphology, susceptibility to demethylation Inhibitor fungicides, and sporulation. Phytopathology 2003, 93, 1354–1359. [Google Scholar] [CrossRef]
- Yu, P.L.; Wang, C.L.; Chen, P.Y.; Lee, M.H. YAP1 homologue-mediated redox sensing is crucial for a successful infection by Monilinia fructicola. Mol. Plant Pathol. 2016, 1, 1–15. [Google Scholar]
- Marek, S.M.; Pan, Z.; Ciuffetti, L.M.; Bostock, R.M. Agrobacterium tumefaciens-mediated transformation of Monilinia fructicola with green fluorescent protein. In Proceedings of the APS Annual Meeting, Milkwaukee, WI, USA, 27–31 July 2002. P2002-0673-AMA. [Google Scholar]
- Li, G.; Zhou, Z.; Liu, G.; Zheng, F.; He, C. Characterization of T-DNA insertion patterns in the genome of rice blast fungus Magnaporthe oryzae. Curr. Genet. 2007, 51, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, J.; Jeon, J.; Chi, M.H.; Goh, J.; Yoo, S.Y.; Park, J.; Jung, K.; Kim, H.; Rho, H.S.; et al. Genome-wide analysis of T-DNA integration into the chromosomes of Magnaporthe oryzae. Mol. Microbiol. 2007, 66, 371–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Patel, G.; Heist, M.; Betts, M.F.; Tucker, S.L.; Galadima, N.; Donofrio, N.M.; Brown, D.; Mitchell, T.K.; Xu, J.R.; et al. A systematic analysis of T-DNA insertion events in Magnaporthe oryzae. Fungal Genet. Biol. 2007, 44, 1050–1064. [Google Scholar] [CrossRef] [PubMed]
- Bourras, S.; Meyer, M.; Grandaubert, J.; Lapalu, N.; Fudal, I.; Linglin, J.; Ollivier, B.; Blaise, F.; Balesdent, M.-H.; Rouxel, T.; et al. Incidence of genome structure, DNA asymmetry, and cell physiology on T-DNA integration in chromosomes of the phytopathogenic fungus Leptosphaeria maculans. G3 Genes Genomes Genet. 2012, 2, 891–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarino, M.; Etxebeste, O.; Mendizabal, G.; Garzia, A.; Ugalde, U.; Espeso, E.A. Boron tolerance in Aspergillus nidulans is sustained by the SltA pathway through the SLC-family transporters SbtA and SbtB. Genes 2017, 8, 188. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Description | Reference |
|---|---|---|---|
| gfpF | CCCATGAGTAAAGGAGAAGAACTT | gfp gene | [18] |
| gfpR | CTATTTGTATAGTTCATCCATGCCATGTGTA | ||
| hphF | TTCGATGTAGGAGGGCGTGGATATG | hph gene | [18] |
| hphR | GGTTTCCACTATCGGCGAGTACTTC | ||
| LB1 | TTAATTGCGTTGCGCTCACTGC | TAIL-PCR | [18] |
| LB2 | GCTTTCCAGTCGGGAAACCTGTC | ||
| LB3 | GAGCAATTCGGCGTTAATTCAGT | ||
| RB1 | GGCACTGGCCGTCGTTTTACAAC | ||
| RB2 | AACGTCGTGACTGGGAAAACCCT | ||
| RB3 | CCCTTCCCAACAGTTGCGCA | ||
| AD1 | WGTGNAGWANCANAGA | ||
| AD2 | NGTCGASWGANAWGAA | ||
| InsLB (Atd9) | AATACCCTGTTGAGATTTGG | Left/right border junction of Atd9 strain | This study |
| InsRB (Atd9) | TACTGGCTAGGCAAGAGTCTGG | ||
| scf005-F | TCATCATAACGGCAGGGAGG | Amplification of putative deletion on Atd9 strain | This study |
| scf005-R | GCCACCACAAATTCCAAGCG | ||
| scf014-F | TGTGGCGTCTGCTTGTATCC | Amplification of putative duplication on Atd9 strain | This study |
| sc014-R | TTTGGCGAGGTCATCATAGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Pires, S.; Espeso, E.A.; Baró-Montel, N.; Torres, R.; Melgarejo, P.; De Cal, A. Labeling of Monilinia fructicola with GFP and Its Validation for Studies on Host-Pathogen Interactions in Stone and Pome Fruit. Genes 2019, 10, 1033. https://doi.org/10.3390/genes10121033
Rodríguez-Pires S, Espeso EA, Baró-Montel N, Torres R, Melgarejo P, De Cal A. Labeling of Monilinia fructicola with GFP and Its Validation for Studies on Host-Pathogen Interactions in Stone and Pome Fruit. Genes. 2019; 10(12):1033. https://doi.org/10.3390/genes10121033
Chicago/Turabian StyleRodríguez-Pires, Silvia, Eduardo Antonio Espeso, Nuria Baró-Montel, Rosario Torres, Paloma Melgarejo, and Antonieta De Cal. 2019. "Labeling of Monilinia fructicola with GFP and Its Validation for Studies on Host-Pathogen Interactions in Stone and Pome Fruit" Genes 10, no. 12: 1033. https://doi.org/10.3390/genes10121033