Upregulated microRNA-106a Promotes Porcine Preadipocyte Proliferation and Differentiation by Targeting Different Genes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Porcine Preadipocytes
2.2. Transfection of miRNA Agomir
- Ssc-NC sense: 5-UUCUCCGAACGUGUCACGUTT-3;
- Ssc-antisense: 5-ACGUGACACGUUCGGAGAATT-3;
- Ssc-miR-106a sense: 5-AAAAGUGCUUACAGUGCAGGUAGC-3;
- Ssc-antisense: 5-UACCUGCACUGUAAGCACUUUUUU-3.
2.3. RNA Extractions and RT-qPCR
2.4. Western Blot Analysis
2.5. 5-Ethynyl-20-Deoxyuridine (EdU) Staining
2.6. Cell Counting Kit-8 (CCK-8)
2.7. Flow Cytometry
2.8. Luciferase Reporter Assay
2.9. BODIPY Staining of Lipid Droplets
2.10. Oil Red O Staining and Dye Extraction Analysis
2.11. Bioinformatics Method
2.12. Statistical Analysis
3. Results
3.1. miR-106a is Widely Expressed in Various Porcine Tissues and is Closely Related to The Cell Cycle and Adipose Metabolism
3.2. Upregulated miR-106a Promotes the Proliferation of Porcine Preadipocytes
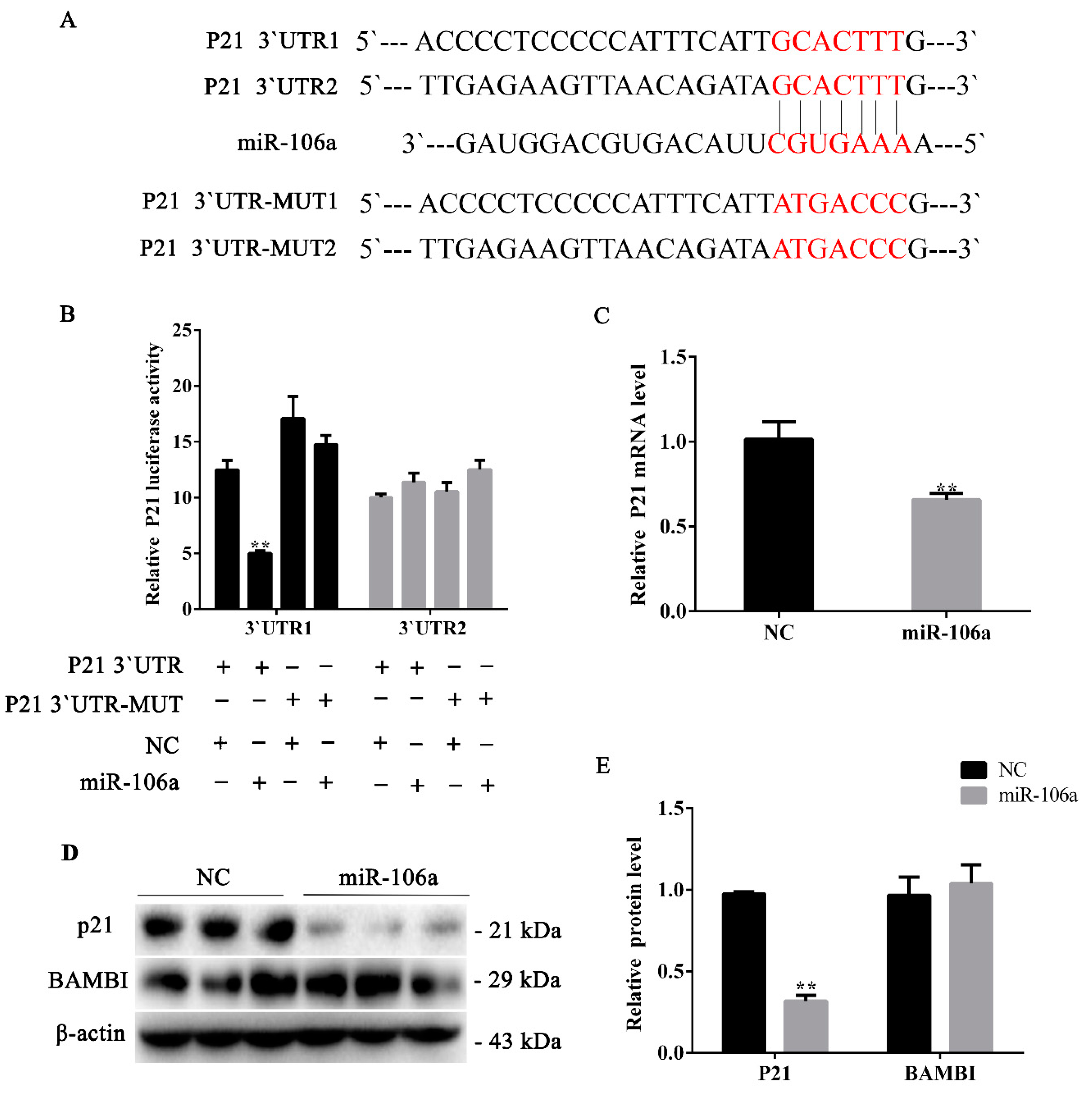
3.3. miR-106a Can Target p21 Instead of BAMBI in Proliferative Phase
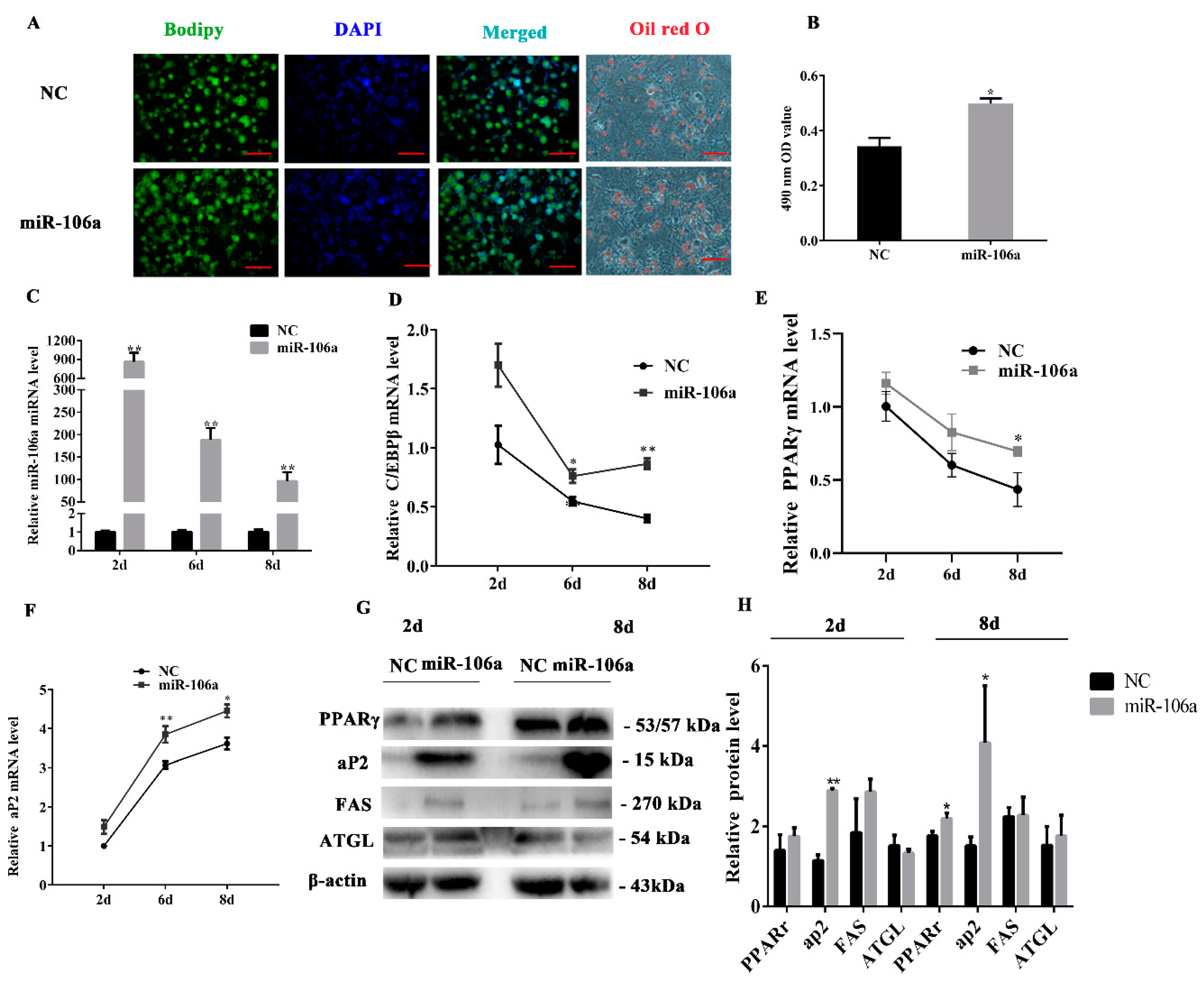
3.4. miR-106a Promotes the Differentiation of Porcine Preadipocytes
3.5. miR-106a Can Target BAMBI Instead of p21 during Porcine Preadipocytes Differentiation.
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mamudu, H.M.; Alamian, A.; Paul, T.; Subedi, P.; Wang, L.; Jones, A.; Alamin, A.E.; Stewart, D.; Blackwell, G.; Budoff, M. Diabetes, subclinical atherosclerosis and multiple cardiovascular risk factors in hard-to-reach asymptomatic patients. Diabetes Vasc. Dis. Res. 2018, 15, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasky, T.M.; Kabat, G.C.; Ho, G.Y.F.; Thomson, C.A.; Nicholson, W.K.; Barrington, W.E.; Bittoni, M.A.; Wassertheil-Smoller, S.; Rohan, T.E. C-reactive protein concentration and risk of selected obesity-related cancers in the Women’s Health Initiative. Cancer Causes Control 2018, 29, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Drummen, M.; Tischmann, L.; Gatta-Cherifi, B.; Adam, T.; Westerterp-Plantenga, M. Dietary Protein and Energy Balance in Relation to Obesity and Co-morbidities. Front. Endocrinol. (Lausanne) 2018, 9, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavo, L.; Busetto, L.; Cesaretti, M.; Zelber-Sagi, S.; Deutsch, L.; Iannelli, A. Nutritional issues in patients with obesity and cirrhosis. World J. Gastroenterol. 2018, 24, 3330–3346. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.R.; Baas, T.J.; Stalder, K.J. Results from six generations of selection for intramuscular fat in Duroc swine using real-time ultrasound. II. Genetic parameters and trends. J. Anim. Sci. 2010, 88, 69–79. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef]
- Xu, D.; Sun, L. Role of microRNA biogenesis in adipocyte and lipodystrophy. Adipocyte 2015, 4, 222–224. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Zhang, Z.; Qiu, X.; Chu, H.; Sun, X.; Zheng, X.; Zhao, J.; Li, Q. Analysis of the miRNA and mRNA involved in osteogenesis of adipose-derived mesenchymal stem cells. Exp. Ther. Med. 2018, 16, 1111–1120. [Google Scholar] [CrossRef]
- Mieczkowska, A.; Schumacher, A.; Filipowicz, N.; Wardowska, A.; Zielinski, M.; Madanecki, P.; Nowicka, E.; Langa, P.; Deptula, M.; Zielinski, J.; et al. Immunophenotyping and transcriptional profiling of in vitro cultured human adipose tissue derived stem cells. Sci. Rep. 2018, 8, 11339. [Google Scholar] [CrossRef]
- Mudhasani, R.; Imbalzano, A.N.; Jones, S.N. An essential role for Dicer in adipocyte differentiation. J. Cell. Biochem. 2010, 110, 812–816. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Tang, J.; Hu, X.; Bao, P.; Pan, J.; Chen, Z.; Xian, J. MiR-27b Impairs Adipocyte Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells by Targeting LPL. Cell. Physiol. Biochem. 2018, 47, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y.O.; Garufi, G.; Pasarica, M.; Seyhan, A.A. Elevated and Correlated Expressions of miR-24, miR-30d, miR-146a, and SFRP-4 in Human Abdominal Adipose Tissue Play a Role in Adiposity and Insulin Resistance. Int. J. Endocrinol. 2018, 2018, 7351902. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.; Wang, Q.; Wang, J.; Huang, J.; Jiang, S.; Xiao, R.; Liu, Z.; Yang, F. Expression Pattern and Regulatory Role of microRNA-23a in Conjugated Linoleic Acids-Induced Apoptosis of Adipocytes. Cell. Physiol. Biochem. 2016, 40, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, Y.; Zhang, Y.; Zhang, Y.; Chen, L.; Mo, D. Up-regulated miR-145 expression inhibits porcine preadipocytes differentiation by targeting IRS1. Int. J. Biol. Sci. 2012, 8, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Qiu, Y.; Liu, S.; Dong, P.; Ning, X.; Li, Y.; Yang, G.; Sun, S. Over-expressed MiR-103 promotes porcine adipocyte differentiation. Sheng Wu Gong Cheng Xue Bao 2012, 28, 927–936. [Google Scholar] [PubMed]
- Pan, S.; Yang, X.; Jia, Y.; Li, R.; Zhao, R. Microvesicle-shuttled miR-130b reduces fat deposition in recipient primary cultured porcine adipocytes by inhibiting PPAR-g expression. J. Cell. Physiol. 2014, 229, 631–639. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nakajima, I.; Chikuni, K.; Kojima, M.; Awata, T.; Mikawa, S. MicroRNA-33b downregulates the differentiation and development of porcine preadipocytes. Mol. Biol. Rep. 2014, 41, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Bahari, F.; Emadi-Baygi, M.; Nikpour, P. miR-17-92 host gene, uderexpressed in gastric cancer and its expression was negatively correlated with the metastasis. Indian J. Cancer 2015, 52, 22–25. [Google Scholar] [CrossRef]
- Li, Y.; Lauriola, M.; Kim, D.; Francesconi, M.; D’Uva, G.; Shibata, D.; Malafa, M.P.; Yeatman, T.J.; Coppola, D.; Solmi, R.; et al. Adenomatous polyposis coli (APC) regulates miR17-92 cluster through β-catenin pathway in colorectal cancer. Oncogene 2016, 35, 4558–4568. [Google Scholar] [CrossRef]
- Meng, W.J.; Yang, L.; Ma, Q.; Zhang, H.; Adell, G.; Arbman, G.; Wang, Z.Q.; Li, Y.; Zhou, Z.G.; Sun, X.F. MicroRNA Expression Profile Reveals miR-17-92 and miR-143-145 Cluster in Synchronous Colorectal Cancer. Medince (Baltim.) 2015, 94, e1297. [Google Scholar] [CrossRef]
- Yu, G.; Tang, J.Q.; Tian, M.L.; Li, H.; Wang, X.; Wu, T.; Zhu, J.; Huang, S.J.; Wan, Y.L. Prognostic values of the miR-17-92 cluster and its paralogs in colon cancer. J. Surg. Oncol. 2012, 106, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.J.; Grandy, W.A.; Balwierz, P.J.; Dimitrova, Y.A.; Pachkov, M.; Ciaudo, C.; Nimwegen, E.; Zavolan, M. Embryonic stem cell-specific microRNAs contribute to pluripotency by inhibiting regulators of multiple differentiation pathways. Nucleic Acids Res. 2014, 42, 9313–9326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic stem cell-specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Judson, R.L.; Babiarz, J.E.; Venere, M.; Blelloch, R. Embryonic stem cell-specific microRNAs promote induced pluripotency. Nat. Biotechnol. 2009, 27, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Z.Y.; Chou, C.H.; Lu, C.Y.; Singh, S.; Yu, S.L.; Li, S.S. Proteomic comparison of human embryonic stem cells with their differentiated fibroblasts: Identification of 206 genes targeted by hES cell-specific microRNAs. Kaohsiung J. Med. Sci. 2011, 27, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, F.T.; Mountain, A.J.; Kelly, M.P.; Hall, C.; Rigopoulos, A.; Johns, T.G.; Smyth, F.E.; Brechbiel, M.W.; Nice, E.C.; Burgess, A.W.; et al. Enhanced efficacy of radioimmunotherapy with 90Y-CHX-A’’-DTPA-hu3S193 by inhibition of epidermal growth factor receptor (EGFR) signaling with EGFR tyrosine kinase inhibitor AG1478. Clin. Cancer Res. 2005, 11, 7080s–7086s. [Google Scholar] [CrossRef]
- Xu, Y.; Niu, J.; Xi, G.; Niu, X.; Wang, Y.; Guo, M.; Yangzong, Q.; Yao, Y.; Sizhu, S.L.; Tian, J. TGF-β1 resulting in differential microRNA expression in bovine granulosa cells. Gene 2018, 663, 88–100. [Google Scholar] [CrossRef]
- Huang, Q.; Ma, Q. MicroRNA-106a inhibits cell proliferation and induces apoptosis in colorectal cancer cells. Oncol. Lett. 2018, 15, 8941–8944. [Google Scholar] [CrossRef]
- Luo, B.; Kang, N.; Chen, Y.; Liu, L.; Zhang, Y. Oncogene miR-106a promotes proliferation and metastasis of prostate cancer cells by directly targeting PTEN in vivo and in vitro. Minerva Med. 2018, 109, 24–30. [Google Scholar] [CrossRef]
- Li, H.; Li, T.; Wang, S.; Wei, J.; Fan, J.; Li, J.; Han, Q.; Liao, L.; Shao, C.; Zhao, R.C. miR-17-5p and miR-106a are involved in the balance between osteogenic and adipogenic differentiation of adipose-derived mesenchymal stem cells. Stem Cell Res. 2013, 10, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Chen, F.F.; Ge, J.; Zhu, J.Y.; Shi, X.E.; Li, X.; Yu, T.Y.; Chu, G.Y.; Yang, G.S. miR-429 Inhibits Differentiation and Promotes Proliferation in Porcine Preadipocytes. Int. J. Mol. Sci. 2016, 17, 2047. [Google Scholar] [CrossRef]
- Deiuliis, J.A. MicroRNAs as regulators of metabolic disease: Pathophysiologic significance and emerging role as biomarkers and therapeutics. Int. J. Obes. (Lond.) 2016, 40, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, M.; Zhu, H.; Zhang, W.; He, S.; Hu, C.; Quan, L.; Bai, J.; Xu, N. miR-106a is frequently upregulated in gastric cancer and inhibits the extrinsic apoptotic pathway by targeting FAS. Mol. Carcinog. 2013, 52, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, F.; Sheng, X.G.; Zhang, S.Q.; Chen, Y.T.; Liu, B.W. MicroRNA-106a regulates phosphatase and tensin homologue expression and promotes the proliferation and invasion of ovarian cancer cells. Oncol. Rep. 2016, 36, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Huo, Z.; Song, X.; Chen, X.; Tian, X.; Wang, X. mir-106a regulates cell proliferation and apoptosis of colon cancer cells through targeting the PTEN/PI3K/AKT signaling pathway. Oncol. Lett. 2018, 15, 3197–3201. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.C.; Wang, J.; Kong, J.; Qi, Y.; Quigg, R.J.; Li, X. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc. Natl. Acad. Sci. USA 2008, 105, 2889–2894. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Wang, X.; Chen, X.; Cai, R.; Chen, F.; Dong, W.; Yang, G.; Pang, W. MicroRNA-432 targeting E2F3 and P55PIK inhibits myogenesis through PI3K/AKT/mTOR signaling pathway. RNA Biol. 2017, 14, 347–360. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, H.; He, X.; Song, H.; Qiang, Y.; Li, Y.; Gao, J.; Wang, Z. miR-106a* inhibits the proliferation of renal carcinoma cells by targeting IRS-2. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 8389–8398. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, M.; Zhu, H.; Zhang, W.; He, S.; Hu, C.; Quan, L.; Bai, J.; Xu, N. Suppression of p21 by c-Myc through members of miR-17 family at the post-transcriptional level. Int. J. Oncol. 2010, 37, 1315–1321. [Google Scholar] [Green Version]
- Gibcus, J.H.; Kroesen, B.J.; Koster, R.; Halsema, N.; de Jong, D.; de Jong, S.; Poppema, S.; Kluiver, J.; Diepstra, A.; van den Berg, A. MiR-17/106b seed family regulates p21 in Hodgkin’s lymphoma. J. Pathol. 2011, 225, 609–617. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol. Biol. 2017, 1509, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Li, J.; Liu, H.; Zhou, F.; Liu, M. MiR-106a promotes tumor growth, migration, and invasion by targeting BCL2L11 in human endometrial adenocarcinoma. Am. J. Transl. Res. 2017, 9, 4984–4993. [Google Scholar] [PubMed]
- Luo, X.; Hutley, L.J.; Webster, J.A.; Kim, Y.H.; Liu, D.F.; Newell, F.S.; Widberg, C.H.; Bachmann, A.; Turner, N.; Schmitz-Peiffer, C.; et al. Identification of BMP and activin membrane-bound inhibitor (BAMBI) as a potent negative regulator of adipogenesis and modulator of autocrine/paracrine adipogenic factors. Diabetes 2012, 61, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, C.; Liao, B.; Cao, J.; Liang, C.; He, X. BAMBI inhibits inflammation through the activation of autophagy in experimental spinal cord injury. Int. J. Mol. Med. 2017, 39, 423–429. [Google Scholar] [CrossRef]
- Zhang, J.C.; Chen, G.; Chen, L.; Meng, Z.J.; Xiong, X.Z.; Liu, H.J.; Jin, Y.; Tao, X.N.; Wu, J.H.; Sun, S.W. TGF-β/BAMBI pathway dysfunction contributes to peripheral Th17/Treg imbalance in chronic obstructive pulmonary disease. Sci. Rep. 2016, 6, 31911. [Google Scholar] [CrossRef]
- Legg, K. Autoimmunity: A controlled performance by BAMBI. Nat. Rev. Rheumatol. 2016, 12, 72. [Google Scholar] [CrossRef]
- Marwitz, S.; Depner, S.; Dvornikov, D.; Merkle, R.; Szczygiel, M.; Muller-Decker, K.; Lucarelli, P.; Wasch, M.; Mairbaurl, H.; Rabe, K.F.; et al. Downregulation of the TGF-β pseudoreceptor BAMBI in non-small cell lung cancer enhances TGF-β signaling and invasion. Cancer Res. 2016, 76, 3785–3801. [Google Scholar] [CrossRef]
- Lin, Z.; Gao, C.; Ning, Y.; He, X.; Wu, W.; Chen, Y.G. The pseudoreceptor BMP and activin membrane-bound inhibitor positively modulates Wnt/β-catenin signaling. J. Biol. Chem. 2008, 283, 33053–33058. [Google Scholar] [CrossRef]
- Yang, X.Z.; Cheng, T.T.; He, Q.J.; Lei, Z.Y.; Chi, J.; Tang, Z.; Liao, Q.X.; Zhang, H.; Zeng, L.S.; Cui, S.Z. LINC01133 as ceRNA inhibits gastric cancer progression by sponging miR-106a-3p to regulate APC expression and the Wnt/β-catenin pathway. Mol. Cancer 2018, 17, 126. [Google Scholar] [CrossRef]
- Fedeli, M.; Riba, M.; Garcia Manteiga, J.M.; Tian, L.; Vigano, V.; Rossetti, G.; Pagani, M.; Xiao, C.; Liston, A.; Stupka, E.; et al. miR-17 approximately 92 family clusters control iNKT cell ontogenesis via modulation of TGF-β signaling. Proc. Natl. Acad. Sci. USA 2016, 113, E8286–E8295. [Google Scholar] [CrossRef]
- Mai, Y.; Zhang, Z.; Yang, H.; Dong, P.; Chu, G.; Yang, G.; Sun, S. BMP and activin membrane-bound inhibitor (BAMBI) inhibits the adipogenesis of porcine preadipocytes through Wnt/β-catenin signaling pathway. Biochem. Cell Biol. 2014, 92, 172–182. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Length | Temp (°C) |
|---|---|---|---|
| PPARγ | F: AGGACTACCAAAGTGCCATCAAA R: GAGGCTTTATCCCCACAGACAC | 164 | 60 |
| aP2 | F: GAGCACCATAACCTTAGATGGA R: AAATTCTGGTAGCCGTGACA | 142 | 60 |
| C/EBPβ | F: GCACAGCGACGAGTACAAGA R: TATGCTGCGTCTCCAGGTTG | 98 | 60 |
| p21 | F:ACGTCTCAGGAGGACCATGT R:AGAAGATCAGCCGGCGTTTG | 166 | 60 |
| BAMBI | F:AGGACAAGGCAACAGGTATTAGC R: GAACCACAACTCTTTGGAGGAAG | 96 | 60 |
| β-actin | F: GGACTTCGAGCAGGAGATGG R: AGGAAGGAGGGCTGGAAGAG | 138 | 60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, K.; Shi, X.; Wang, J.; Yao, Y.; Peng, Y.; Chen, X.; Li, X.; Yang, G. Upregulated microRNA-106a Promotes Porcine Preadipocyte Proliferation and Differentiation by Targeting Different Genes. Genes 2019, 10, 805. https://doi.org/10.3390/genes10100805
Huang K, Shi X, Wang J, Yao Y, Peng Y, Chen X, Li X, Yang G. Upregulated microRNA-106a Promotes Porcine Preadipocyte Proliferation and Differentiation by Targeting Different Genes. Genes. 2019; 10(10):805. https://doi.org/10.3390/genes10100805
Chicago/Turabian StyleHuang, Kuilong, Xin’e Shi, Jie Wang, Ying Yao, Ying Peng, Xiaochang Chen, Xiao Li, and Gongshe Yang. 2019. "Upregulated microRNA-106a Promotes Porcine Preadipocyte Proliferation and Differentiation by Targeting Different Genes" Genes 10, no. 10: 805. https://doi.org/10.3390/genes10100805