Nrp1 is Activated by Konjac Ceramide Binding-Induced Structural Rigidification of the a1a2 Domain

 , and
, and
Abstract
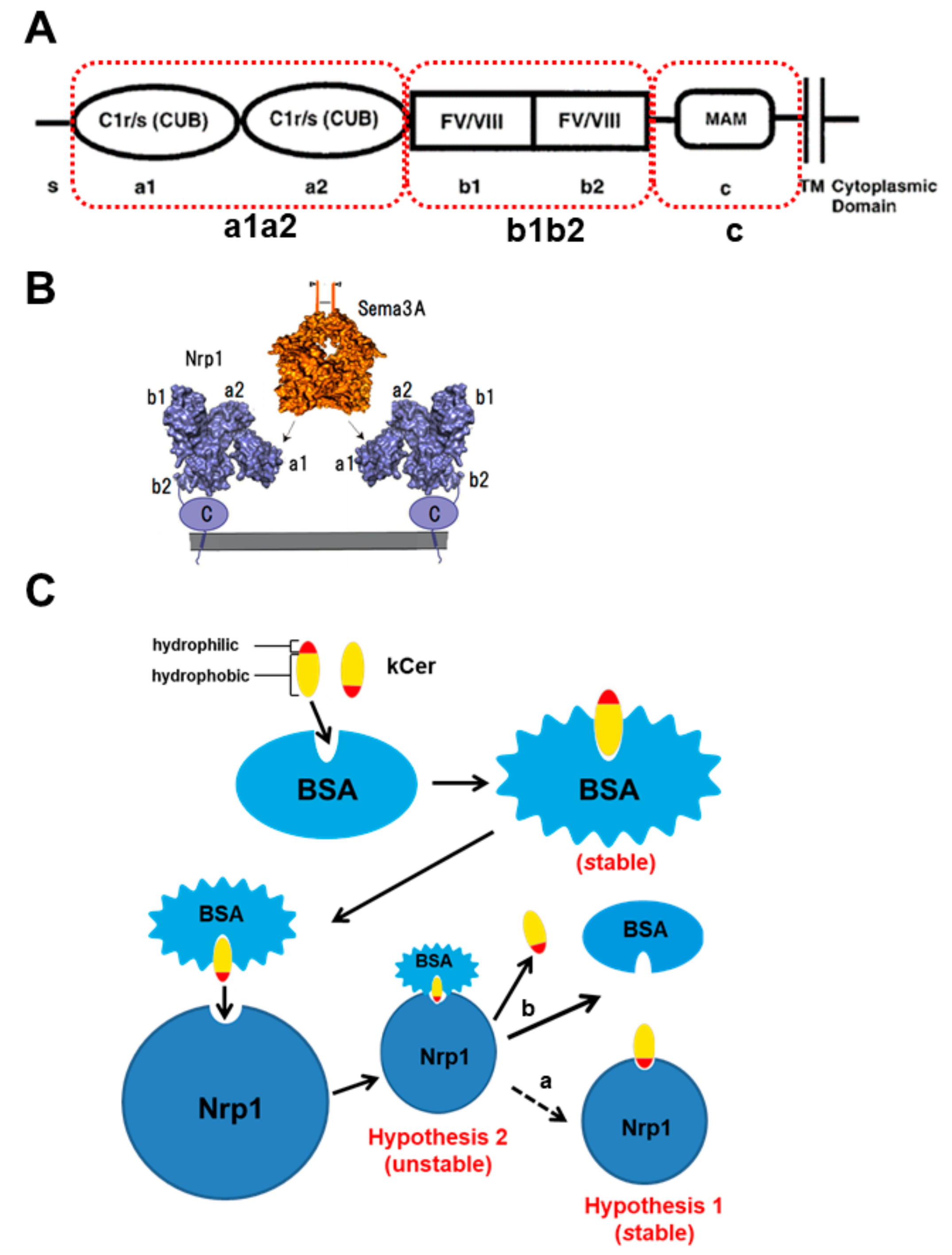
:1. Introduction
2. Materials and Methods
2.1. Preparation of Chemicals
2.1.1. Konjac Ceramide (kCer)
2.1.2. Sphingadienine, d4t,8t-, and 4t,8c-C16Cer
2.1.3. NBD-ceramides (NBD-Cers)
2.1.4. Rhod-Bovine Serum Albumin (Fatty Acid-Free)
2.2. Preparation of Recombinant Proteins
2.3. Preparation of Gene-Silencing Cells
2.4. Co-Immunoprecipitation (Co-IP) of Sema3A and Nrp1 Domain Proteins
2.5. Quantitative Dot Blot Analysis
2.6. Time Course of Fluorescence Intensity of NBD-Cer and Rhod-BSA on the Cell Surface
2.7. Differential Scanning Calorimetry (DSC)
2.8. Circular Dichroism (CD) Spectroscopy
2.9. Molecular Docking
2.10. Statistical Analysis
2.11. Research Rules and Regulation
3. Results
3.1. Co-Immunoprecipitation of AP-Sema3A and Nrp1 Domain Proteins
3.2. Dot-Blotting and Binding Parameters
3.3. Dissociation Time Course for Cell Surface Binding of NBD-Cer and Rhod-BSA
3.4. Comparison of Differential Scanning Calorimetry Thermograms and Circular Dichroism Spectra Between Control and kCer
3.5. Molecular Docking Simulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NGF | nerve growth factor |
| Cer | ceramide |
| GlcCer | glucosylceramide |
| kCer | konjac ceramide |
| EGCase I | endoglycoceramidase I |
| CRMP2 | collapsin response mediator protein 2 |
| pCRMP2 | phospho-collapsin response mediator protein 2 |
| Sema3A | semaphorin 3A |
| AP-Sema3A | alkaline phosphatase-fused Sema3A |
| BSA | bovine serum albumin |
| Rhod-BSA | rhodamine-labeled BSA |
| PBS | phosphate-buffered saline |
| DMEM | Dulbecco’s modified Eagle’s medium |
| IDR | intrinsically disordered region |
| d4t,8t-kCer | ceramides containing d4t,8t-sphingadienine of kCer |
| d4t,8c-kCer | ceramides containing d4t,8c-sphingadienine of kCer |
| NBD-Cer | fluorescent ceramide derivative comprising NBD-dodecanoic acid and long-chain bases (sphingadienine, sphingosine, phytosphingosine) |
| Co-IP | co-immunoprecipitation |
References
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Goshima, Y.; Yamashita, N.; Nakamura, F.; Sasaki, Y. Regulation of dendritic development by semaphorin 3A through novel intracellular remote signaling. Cell Adh. Migr. 2016, 10, 627–640. [Google Scholar] [CrossRef]
- Tamagnone, L.; Comoglio, P.M. Signalling by semaphorin receptors: Cell guidance and beyond. Trends Cell Biol. 2000, 10, 377–383. [Google Scholar] [CrossRef]
- Martin-Satue, M.; Blanco, J. Identification of semaphorin E gene expression in metastatic human lung adenocarcinoma cells by mRNA differential display. J. Surg. Oncol. 1999, 72, 18–23. [Google Scholar] [CrossRef]
- Tseng, C.H.; Murray, K.D.; Jou, M.F.; Hsu, S.M.; Cheng, H.J.; Huang, P.H. Sema3E/plexin-D1 mediated epithelial-to-mesenchymal transition in ovarian endometrioid cancer. PLoS ONE 2011, 6, e19396. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.Q.; Zhou, S.L.; Zhou, Z.J.; Luo, C.B.; Chen, E.B.; Zhan, H.; Wang, P.C.; Dai, Z.; Zhou, J.; Fan, J.; et al. Overexpression of semaphorin 3A promotes tumor progression and predicts poor prognosis in hepatocellular carcinoma after curative resection. Oncotarget 2016, 7, 51733–51746. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, G.; Kumar, S.; Mishra, R.; Patil, T.V.; Kundu, G.C. Semaphorin 3A suppresses tumor growth and metastasis in mice melanoma model. PLoS ONE 2012, 7, e33633. [Google Scholar] [CrossRef]
- Tischoff, I.; Markwarth, A.; Witzigmann, H.; Uhlmann, D.; Hauss, J.; Mirmohammadsadegh, A.; Wittekind, C.; Hengge, U.R.; Tannapfel, A. Allele loss and epigenetic inactivation of 3p21.3 in malignant liver tumors. Int. J. Cancer 2005, 115, 684–689. [Google Scholar] [CrossRef]
- Usuki, S.; Tamura, N.; Yuyama, K.; Tamura, T.; Mukai, K.; Igarashi, Y. Konjac Ceramide (kCer) Regulates NGF-Induced Neurite Outgrowth via the Sema3A Signaling Pathway. J. Oleo Sci. 2018, 67, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Usuki, S.; Tamura, N.; Tamura, T.; Mukai, K.; Igarashi, Y. Characterization of Konjac Ceramide (kCer) Binding to Sema3A Receptor Nrp1. J. Oleo Sci. 2018, 67, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Kou, K.; Nakamura, F.; Aihara, M.; Chen, H.; Seto, K.; Komori-Yamaguchi, J.; Kambara, T.; Nagashima, Y.; Goshima, Y.; Ikezawa, Z. Decreased expression of semaphorin-3A, a neurite-collapsing factor, is associated with itch in psoriatic skin. Acta Derm. Venereol. 2012, 92, 521–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tominaga, M.; Takamori, K. Itch and nerve fibers with special reference to atopic dermatitis: Therapeutic implications. J. Dermatol. 2014, 41, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Ito, Y.; Kawai-Yamada, M. Molecular characterization and targeted quantitative profiling of the sphingolipidome in rice. Plant J. 2016, 88, 681–693. [Google Scholar] [CrossRef]
- Shimada, E.; Aida, K.; Sugawara, T.; Hirata, T. Inhibitory effect of topical maize glucosylceramide on skin photoaging in UVA-irradiated hairless mice. J. Oleo Sci. 2011, 60, 321–325. [Google Scholar] [CrossRef]
- Arai, K.; Mizobuchi, Y.; Tokuji, Y.; Aida, K.; Yamashita, S.; Ohnishi, M.; Kinoshita, M. Effects of Dietary Plant-Origin Glucosylceramide on Bowel Inflammation in DSS-Treated Mice. J. Oleo Sci. 2015, 64, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, T.; Nakano, Y.; Ueda, O.; Mori, H.; Nakashima, M.; Noda, A.; Ishizaki, C.; Mizoguchi, M. Oral Intake of Glucosylceramide Improves Relatively High level of Transepideramal Water Loss in Mice and Healthy Human Subjects. J. Health Sci. 2008, 54, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Usuki, S.; Tamura, N.; Sakai, S.; Tamura, T.; Mukai, K.; Igarashi, Y. Chemoenzymatically preapred konjac ceramide inhibits NGF-induced neurite outgrowth by a semaphorin 3A-like action. Biochem. Biophys. Rep. 2016, 5, 160–167. [Google Scholar]
- Usuki, S.; Tamura, N.; Tamura, T.; Tanji, K.; Mikami, D.; Mukai, K.; Igarashi, Y. Neurite Outgrowth and Morphological Changes Induced by 8-trans Unsaturation of Sphingadienine in kCer Molecular Species. Int. J. Mol. Sci. 2019, 20, E2116. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.C.; Kreusch, A.; McMullan, D.; Ng, K.; Spraggon, G. Crystal structure of the human neuropilin-1 b1 domain. Structure 2003, 11, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Strittmatter, S.M. Plexina1 autoinhibition by the plexin sema domain. Neuron 2001, 29, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.F.; Vander Kooi, C.W. Neuropilin Functions as an Essential Cell Surface Receptor. J. Biol. Chem. 2015, 290, 29120–29126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, D.; Sakai, S.; Sasaki, S.; Igarashi, Y. Effects of Asterias amurensis-derived Sphingoid Bases on the de novo Ceramide Synthesis in Cultured Normal Human Epidermal Keratinocytes. J. Oleo Sci. 2016, 65, 671–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T.; Kinoshita, M.; Ohnishi, M.; Nagata, J.; Saito, M. Digestion of maize sphingolipids in rats and uptake of sphingadienine by Caco-2 cells. J. Nutr. 2003, 133, 2777–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usuki, S.; Tamura, N.; Tamura, T.; Higashiyama, S.; Tanji, K.; Mitsutake, S.; Inoue, A.; Aoki, J.; Mukai, K.; Igarashi, Y. Konjac ceramide (kCer) regulates keratinocyte migration by Sema3A-like repulsion mechanism. Biochem. Biophys. Rep. 2019, 17, 132–138. [Google Scholar] [CrossRef]
- Saitoh, T.; Ikegami, T.; Nakayama, M.; Teshima, K.; Akutsu, H.; Hase, T. NMR study of the electron transfer complex of plant ferredoxin and sulfite reductase: mapping the interaction sites of ferredoxin. J. Biol. Chem. 2006, 281, 10482–10488. [Google Scholar] [CrossRef] [Green Version]
- Tochio, N.; Umehara, K.; Uewaki, J.I.; Flechsig, H.; Kondo, M.; Dewa, T.; Sakuma, T.; Yamamoto, T.; Saitoh, T.; Togashi, Y.; et al. Non-RVD mutations that enhance the dynamics of the TAL repeat array along the superhelical axis improve TALEN genome editing efficacy. Sci. Rep. 2016, 6, 37887. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.J.; Malinauskas, T.; Weir, G.A.; Cader, M.Z.; Siebold, C.; Jones, E.Y. Neuropilins lock secreted semaphorins onto plexins in a ternary signaling complex. Nat. Struct. Mol. Biol. 2012, 19, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- ADT/AutoDockTools. Available online: http://autodock.scripps.edu/resources/adt/ (accessed on 21 February 2020).
- Schrödinger, L.L.C. The PyMOL Molecular Graphics System. Available online: https://pymol.org/2/ (accessed on 21 February 2020).
- Thudichum, J.L.W. Reports of the Medical Officer of Privy Council and Local Government Board; Harvard University Press: Cambridge, MA, USA, 1874. [Google Scholar]
- Obeid, L.M.; Linardic, C.M.; Karolak, L.A.; Hannun, Y.A. Programmed cell death induced by ceramide. Science 1993, 259, 1769–1771. [Google Scholar] [CrossRef]
- von Haefen, C.; Wieder, T.; Gillissen, B.; Starck, L.; Graupner, V.; Dorken, B.; Daniel, P.T. Ceramide induces mitochondrial activation and apoptosis via a Bax-dependent pathway in human carcinoma cells. Oncogene 2002, 21, 4009–4019. [Google Scholar] [CrossRef] [Green Version]
- Chalfant, C.E.; Spiegel, S. Sphingosine 1-phosphate and ceramide 1-phosphate: expanding roles in cell signaling. J. Cell Sci. 2005, 118, 4605–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihara, A. Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog. Lipid Res. 2016, 63, 50–69. [Google Scholar] [CrossRef] [PubMed]
- Brodowicz, J.; Przegalinski, E.; Muller, C.P.; Filip, M. Ceramide and Its Related Neurochemical Networks as Targets for Some Brain Disorder Therapies. Neurotox. Res. 2018, 33, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohm, B.; Ottemeyer, A.; Lohrum, M.; Puschel, A.W. Plexin/neuropilin complexes mediate repulsion by the axonal guidance signal semaphorin 3A. Mech. Dev. 2000, 93, 95–104. [Google Scholar] [CrossRef]
- Wilson, S.M.; Xiong, W.; Wang, Y.; Ping, X.; Head, J.D.; Brittain, J.M.; Gagare, P.D.; Ramachandran, P.V.; Jin, X.; Khanna, R. Prevention of posttraumatic axon sprouting by blocking collapsin response mediator protein 2-mediated neurite outgrowth and tubulin polymerization. Neuroscience 2012, 210, 451–466. [Google Scholar] [CrossRef] [Green Version]
- Kurschat, P.; Bielenberg, D.; Rossignol-Tallandier, M.; Stahl, A.; Klagsbrun, M. Neuron restrictive silencer factor NRSF/REST is a transcriptional repressor of neuropilin-1 and diminishes the ability of semaphorin 3A to inhibit keratinocyte migration. J. Biol. Chem. 2006, 281, 2721–2729. [Google Scholar] [CrossRef] [Green Version]
- Reichert, S.; Scheid, S.; Roth, T.; Herkel, M.; Petrova, D.; Linden, A.; Weberbauer, M.; Esser, J.; Diehl, P.; Grundmann, S.; et al. Semaphorin 3F Promotes Transendothelial Migration of Leukocytes in the Inflammatory Response After Survived Cardiac Arrest. Inflammation 2019, 42, 1252–1264. [Google Scholar] [CrossRef]
- Smith, D.A.; Di, L.; Kerns, E.H. The effect of plasma protein binding on in vivo efficacy: Misconceptions in drug discovery. Nat. Rev. Drug Discov. 2010, 9, 929–939. [Google Scholar] [CrossRef]
- Curry, S.; Brick, P.; Franks, N.P. Fatty acid binding to human serum albumin: New insights from crystallographic studies. Biochim. Biophys. Acta 1999, 1441, 131–140. [Google Scholar] [CrossRef]
- Al-Husseini, J.; Stanton, N.J.; Selassie, C.R.D.; Johal, M.S. The Binding of Drug Molecules to Serum Albumin: The Effect of Drug Hydrophobicity on Binding Strength and Protein Desolvation. Langmuir 2019, 17054–17060. [Google Scholar] [CrossRef]
- Sugase, K.; Dyson, H.J.; Wright, P.E. Mechanism of coupled folding and binding of an intrinsically disordered protein. Nature 2007, 447, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Sako, Y. Single-Molecule Forster Resonance Energy Transfer Measurement Reveals the Dynamic Partially Ordered Structure of the Epidermal Growth Factor Receptor C-Tail Domain. J. Phys. Chem. B 2019, 123, 571–581. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNTL | kCer | kGlcCer | C24Cer | C18Cer | C16Cer | C8Cer | C2Cer |
|---|---|---|---|---|---|---|---|
| 61.4 °C | 63.1 °C | 61.0 °C | 61.9 °C | 61.9 °C | 61.9 °C | 62.5 °C | 64.1 °C |
| Common element/Additive | α-helices | β-sheets | β-turns | Random Coil | Total |
|---|---|---|---|---|---|
| CNTL | 15.5% | 43.4% | 5.2% | 35.0% | 100.0±0.6% |
| kCer | 16.9% | 44.8% | 3.5% | 34.8% | 100.0±0.4% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usuki, S.; Yasutake, Y.; Tamura, N.; Tamura, T.; Tanji, K.; Saitoh, T.; Murai, Y.; Mikami, D.; Yuyama, K.; Monde, K.; et al. Nrp1 is Activated by Konjac Ceramide Binding-Induced Structural Rigidification of the a1a2 Domain. Cells 2020, 9, 517. https://doi.org/10.3390/cells9020517
Usuki S, Yasutake Y, Tamura N, Tamura T, Tanji K, Saitoh T, Murai Y, Mikami D, Yuyama K, Monde K, et al. Nrp1 is Activated by Konjac Ceramide Binding-Induced Structural Rigidification of the a1a2 Domain. Cells. 2020; 9(2):517. https://doi.org/10.3390/cells9020517
Chicago/Turabian StyleUsuki, Seigo, Yoshiaki Yasutake, Noriko Tamura, Tomohiro Tamura, Kunikazu Tanji, Takashi Saitoh, Yuta Murai, Daisuke Mikami, Kohei Yuyama, Kenji Monde, and et al. 2020. "Nrp1 is Activated by Konjac Ceramide Binding-Induced Structural Rigidification of the a1a2 Domain" Cells 9, no. 2: 517. https://doi.org/10.3390/cells9020517