Norrin Protects Retinal Ganglion Cells from Excitotoxic Damage via the Induction of Leukemia Inhibitory Factor

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. NMDA-Induced Damage on Retinal Ganglion Cells
2.3. Overexpression and Purification of Human Recombinant Norrin
2.4. RNA Isolation, cDNA Synthesis and Real-Time rt-PCR Analysis
2.5. Light Microscopy
2.6. TUNEL Labeling
2.7. Statistical Analysis
3. Results
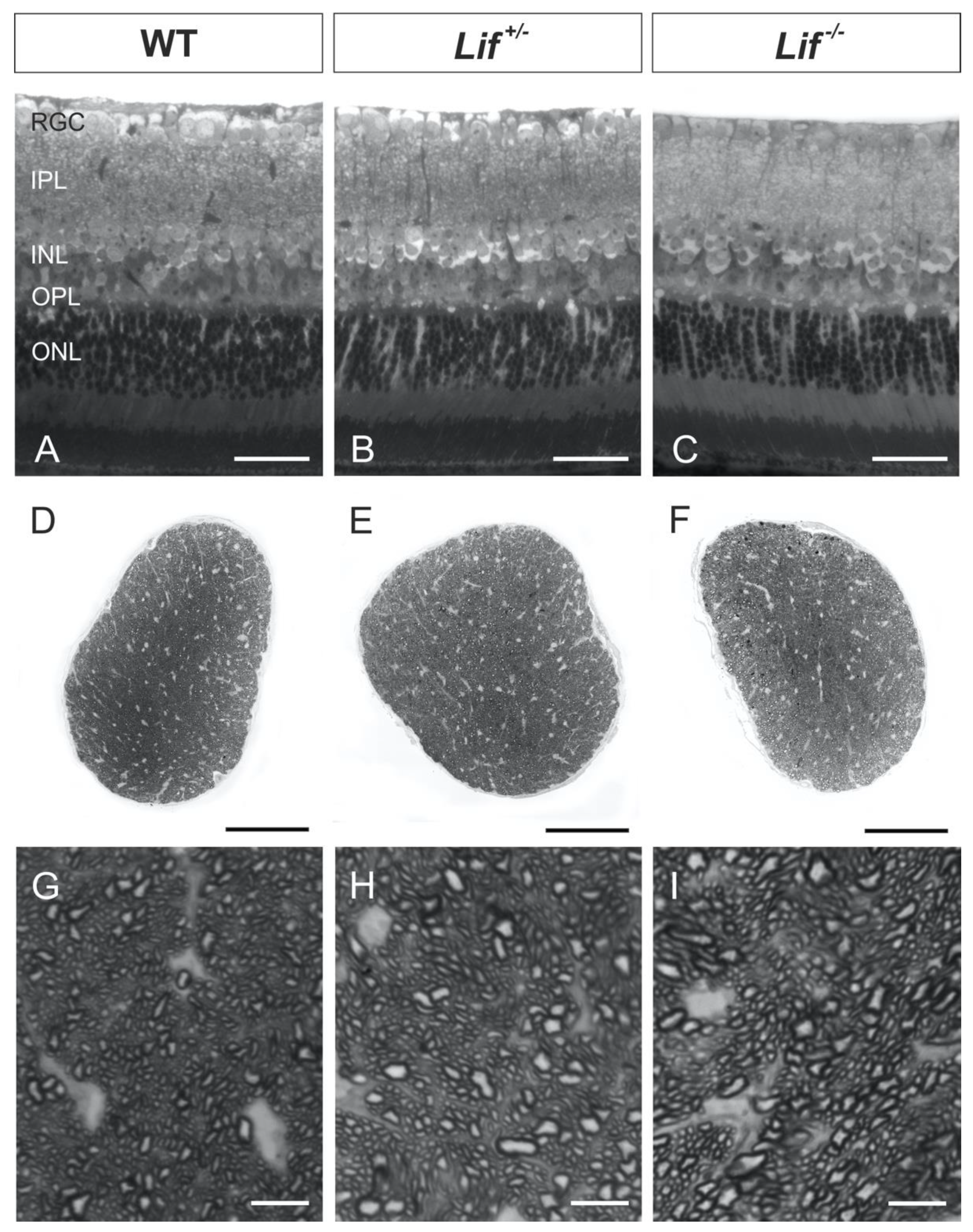
3.1. Lif-Deficient Mice do not Show an Obvious Retinal or Optic Nerve Phenotype
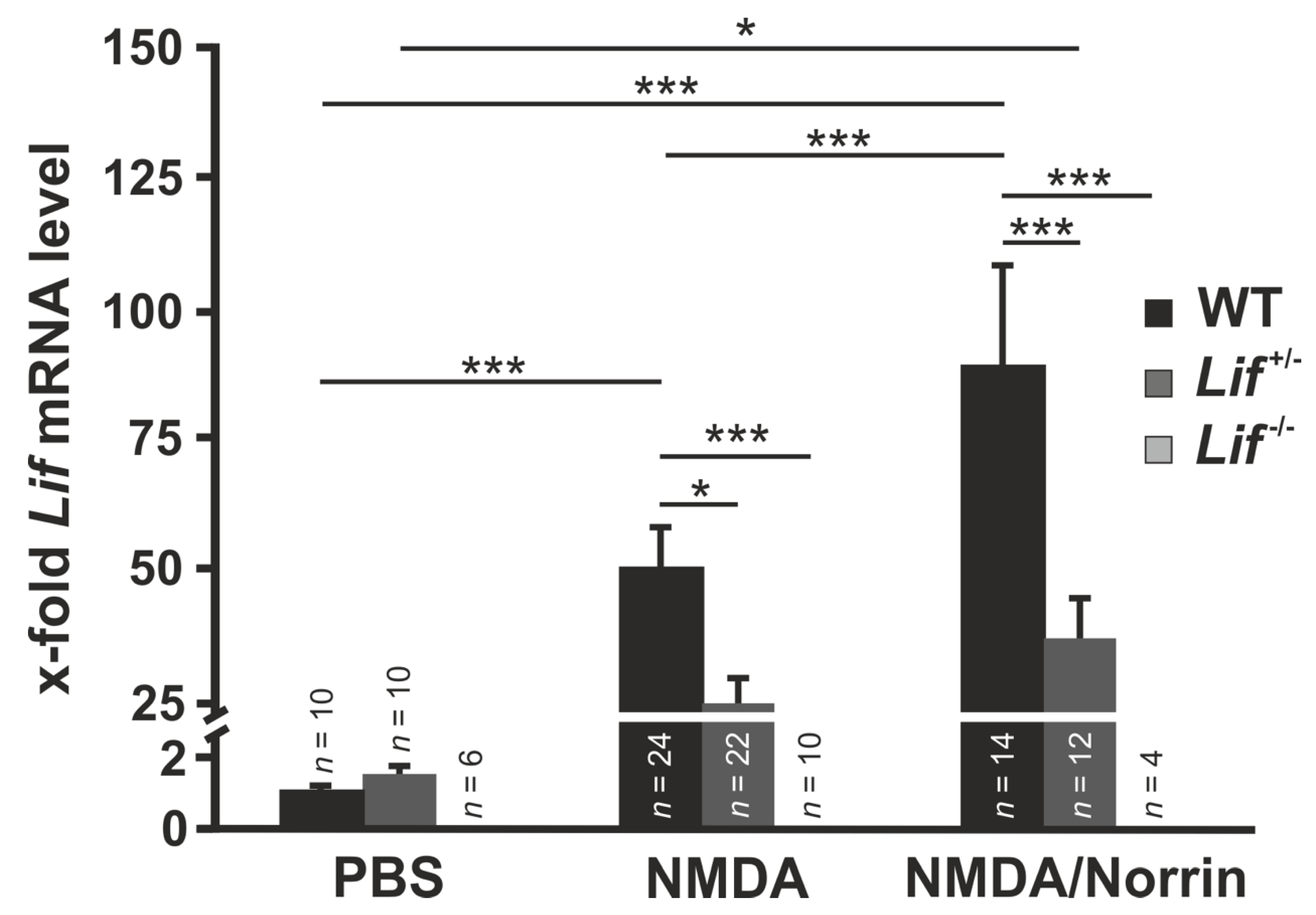
3.2. Norrin Induces Lif Expression in Retinae Following Excitotoxic Damage of Retinal Neurons
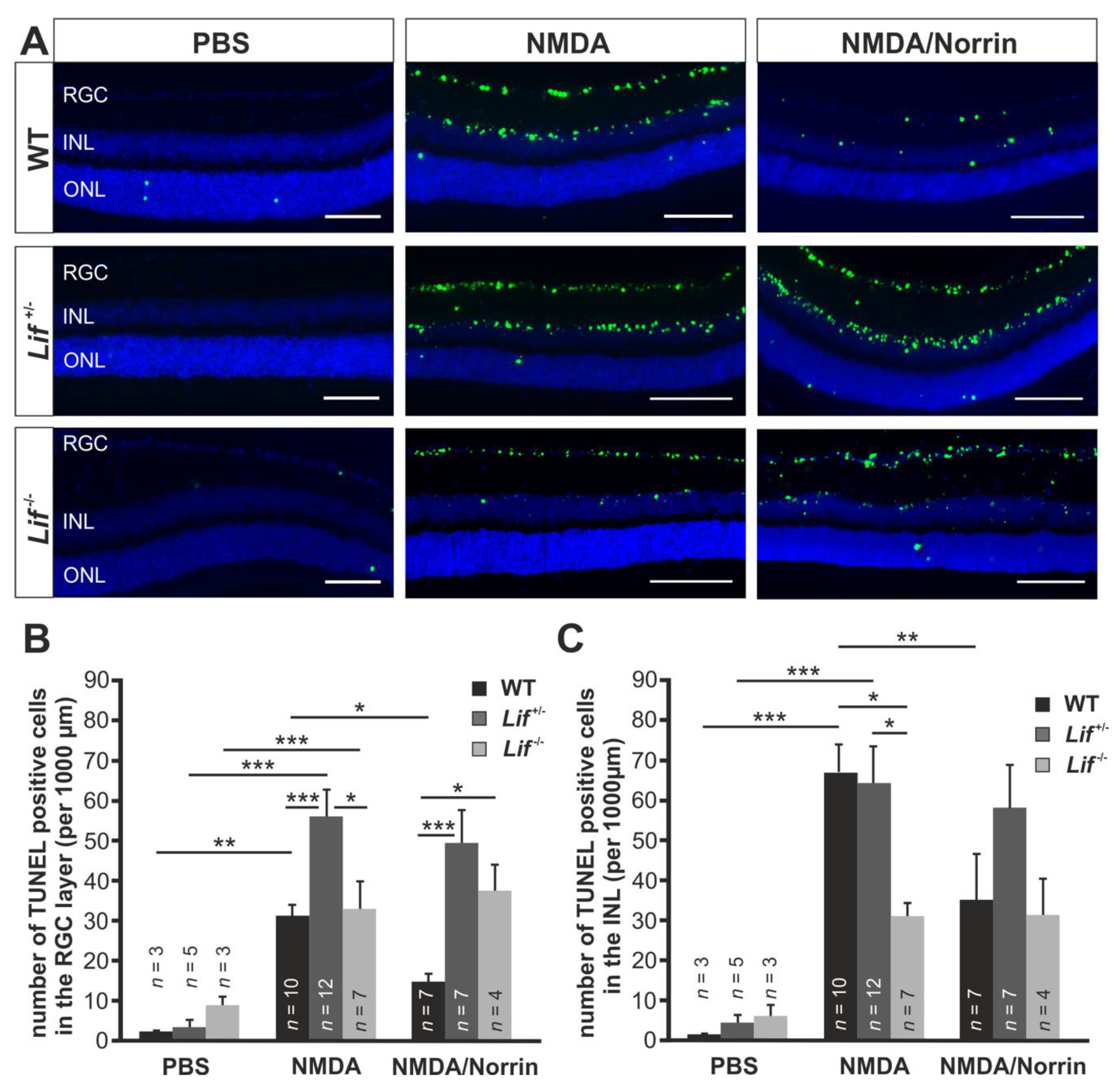
3.3. Norrin Mediates Its Neuroprotective Effect via an Induction of Lif
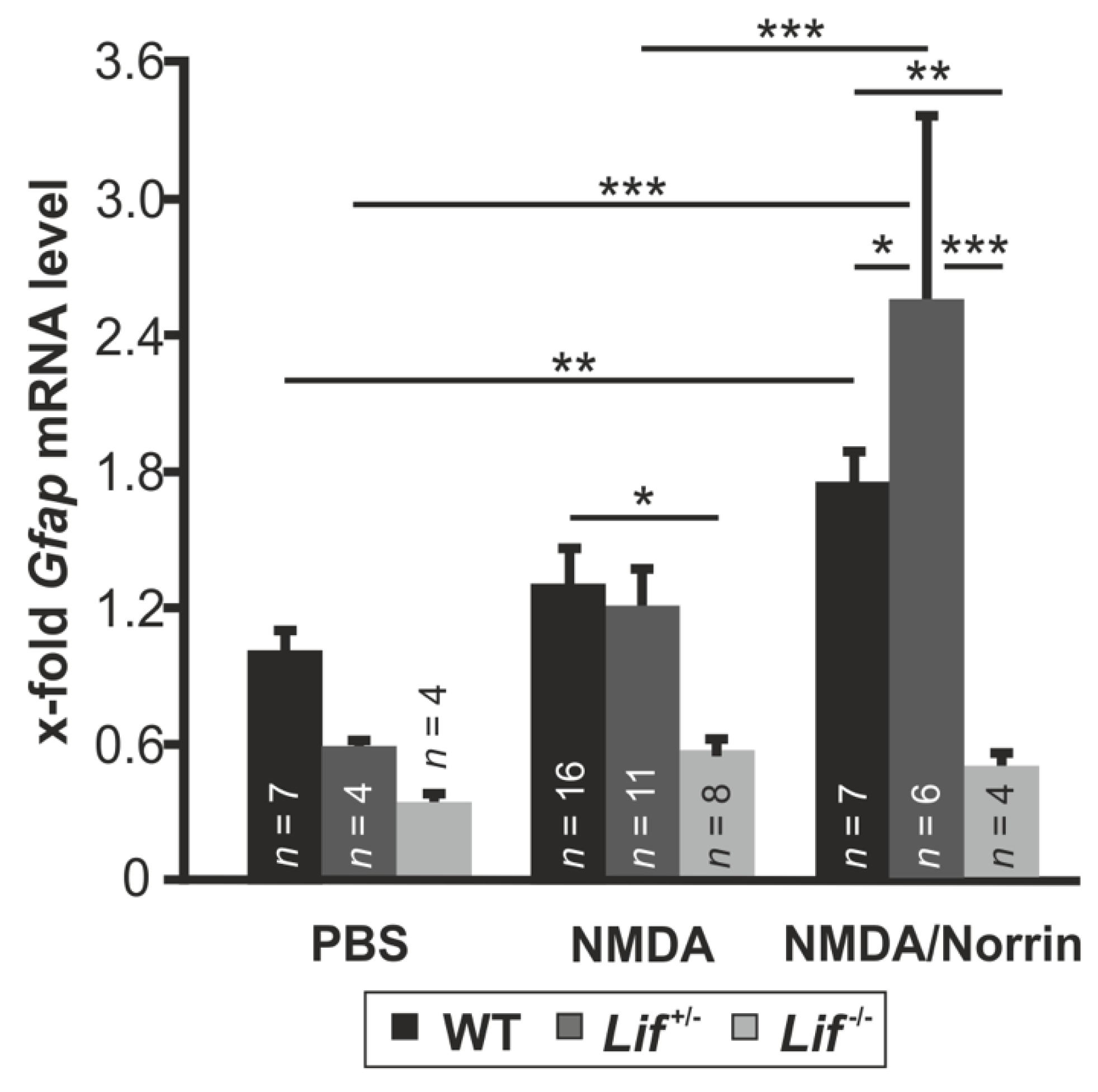
3.4. Norrin Mediates Müller Cell Gliosis via LIF Signaling
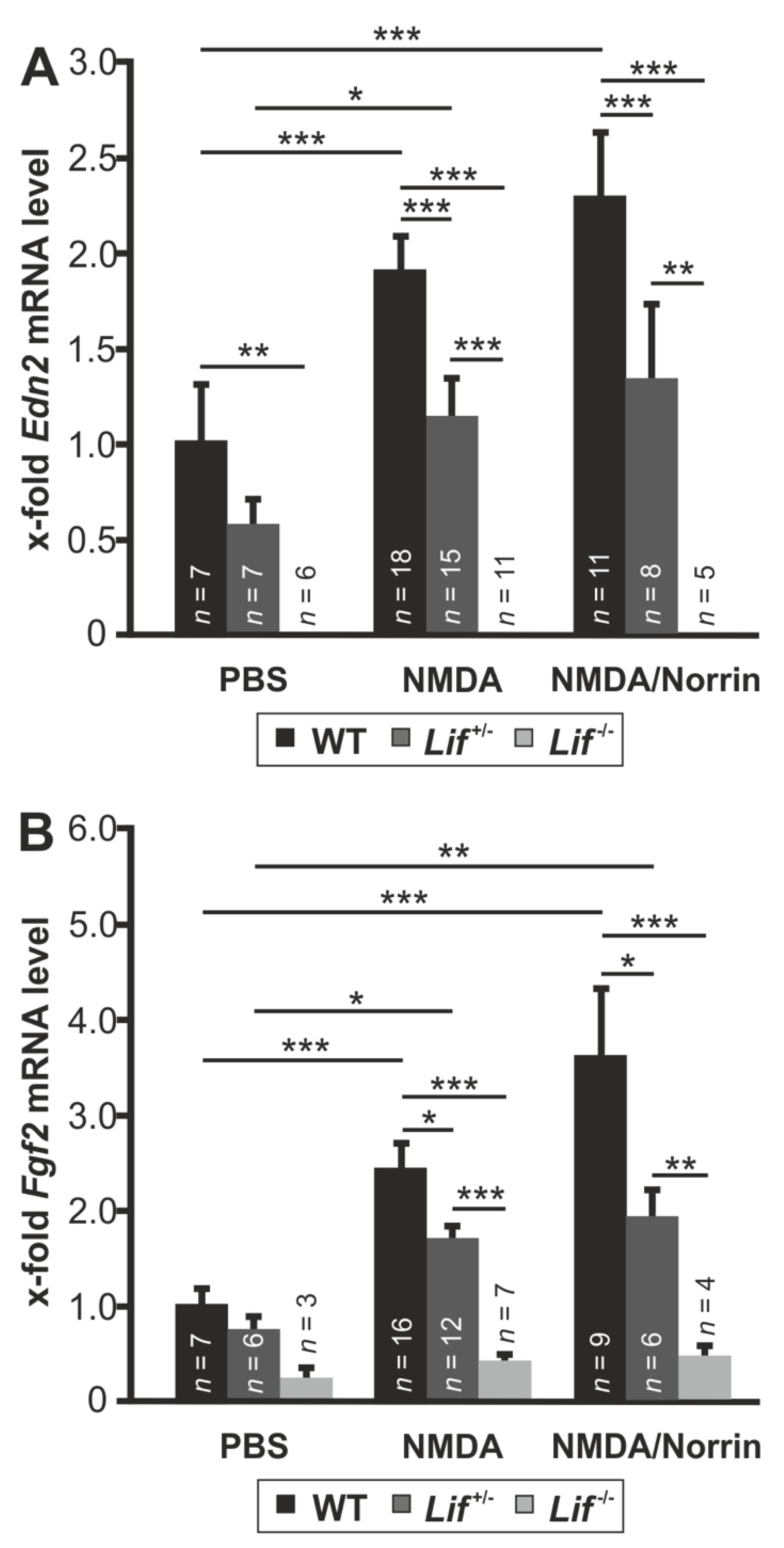
3.5. Norrin Induces the Expression of Edn2 and Fgf2 via Lif Signaling
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oster, S.F.; Deiner, M.; Birgbauer, E.; Sretavan, D.W. Ganglion cell axon pathfinding in the retina and optic nerve. Semin. Cell Dev. Biol. 2004, 15, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu-Wai-Man, P.; Turnbull, D.M.; Chinnery, P.F. Leber hereditary optic neuropathy. J. Med. Genet. 2002, 39, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Nucci, C.; Martucci, A.; Giannini, C.; Morrone, L.A.; Bagetta, G.; Mancino, R. Neuroprotective agents in the management of glaucoma. Eye 2018, 32, 938–945. [Google Scholar] [CrossRef]
- Niwa, M.; Aoki, H.; Hirata, A.; Tomita, H.; Green, P.G.; Hara, A. Retinal Cell Degeneration in Animal Models. Int. J. Mol. Sci. 2016, 17, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Liu, X.L.; Yang, X.L. N-methyl-D-aspartate receptors in the retina. Mol. Neurobiol. 2006, 34, 163–179. [Google Scholar] [CrossRef]
- Lam, T.T.; Abler, A.S.; Kwong, J.M.; Tso, M.O. N-methyl-D-aspartate (NMDA)–induced apoptosis in rat retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2391–2397. [Google Scholar]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.M. Epithelial phenotype and the RPE: Is the answer blowing in the Wnt? Prog. Retin. Eye Res. 2008, 27, 579–595. [Google Scholar] [CrossRef] [Green Version]
- Drenser, K.A. Wnt signaling pathway in retinal vascularization. Eye Brain 2016, 8, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Wang, Y.; Dabdoub, A.; Smallwood, P.M.; Williams, J.; Woods, C.; Kelley, M.W.; Jiang, L.; Tasman, W.; Zhang, K.; et al. Vascular development in the retina and inner ear: Control by Norrin and Frizzled-4, a high-affinity ligand-receptor pair. Cell 2004, 116, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Wang, Y.; Cahill, H.; Yu, M.; Badea, T.C.; Smallwood, P.M.; Peachey, N.S.; Nathans, J. Norrin, frizzled-4 and Lrp5 signaling in endothelial cells controls a genetic program for retinal vascularization. Cell 2009, 139, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlmann, A.; Seitz, R.; Braunger, B.; Seitz, D.; Bosl, M.R.; Tamm, E.R. Norrin promotes vascular regrowth after oxygen-induced retinal vessel loss and suppresses retinopathy in mice. J. Neurosci. 2010, 30, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlmann, A.; Scholz, M.; Goldwich, A.; Chauhan, B.K.; Hudl, K.; Ohlmann, A.V.; Zrenner, E.; Berger, W.; Cvekl, A.; Seeliger, M.W.; et al. Ectopic norrin induces growth of ocular capillaries and restores normal retinal angiogenesis in Norrie disease mutant mice. J. Neurosci. 2005, 25, 1701–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Gottanka, J.; May, C.A.; Welge-Lussen, U.; Berger, W.; Lutjen-Drecoll, E. Retinal vasculature changes in Norrie disease mice. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2450–2457. [Google Scholar]
- Miller, S.J.; Philips, T.; Kim, N.; Dastgheyb, R.; Chen, Z.; Hsieh, Y.C.; Daigle, J.G.; Datta, M.; Chew, J.; Vidensky, S.; et al. Molecularly defined cortical astroglia subpopulation modulates neurons via secretion of Norrin. Nat. Neurosci. 2019, 22, 741–752. [Google Scholar] [CrossRef]
- Dailey, W.A.; Drenser, K.A.; Wong, S.C.; Cheng, M.; Vercellone, J.; Roumayah, K.K.; Feeney, E.V.; Deshpande, M.; Guzman, A.E.; Trese, M.; et al. Norrin treatment improves ganglion cell survival in an oxygen-induced retinopathy model of retinal ischemia. Exp. Eye Res. 2017, 164, 129–138. [Google Scholar] [CrossRef]
- Seitz, R.; Hackl, S.; Seibuchner, T.; Tamm, E.R.; Ohlmann, A. Norrin mediates neuroprotective effects on retinal ganglion cells via activation of the Wnt/beta-catenin signaling pathway and the induction of neuroprotective growth factors in Muller cells. J. Neurosci. 2010, 30, 5998–6010. [Google Scholar] [CrossRef] [Green Version]
- Braunger, B.M.; Ohlmann, A.; Koch, M.; Tanimoto, N.; Volz, C.; Yang, Y.; Bosl, M.R.; Cvekl, A.; Jagle, H.; Seeliger, M.W.; et al. Constitutive overexpression of Norrin activates Wnt/beta-catenin and endothelin-2 signaling to protect photoreceptors from light damage. Neurobiol. Dis. 2013, 50, 1–12. [Google Scholar] [CrossRef]
- Joly, S.; Lange, C.; Thiersch, M.; Samardzija, M.; Grimm, C. Leukemia inhibitory factor extends the lifespan of injured photoreceptors in vivo. J. Neurosci. 2008, 28, 13765–13774. [Google Scholar] [CrossRef] [Green Version]
- Nicola, N.A.; Babon, J.J. Leukemia inhibitory factor (LIF). Cytokine Growth Factor Rev. 2015, 26, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.M.; Pennypacker, K.R. The role of the leukemia inhibitory factor receptor in neuroprotective signaling. Pharmacol. Ther. 2018, 183, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.D.; Yang, X.J. Function and mechanism of CNTF/LIF signaling in retinogenesis. Adv. Exp. Med. Biol. 2010, 664, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agca, C.; Grimm, C. Leukemia inhibitory factor signaling in degenerating retinas. Adv. Exp. Med. Biol. 2014, 801, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.F.; Huang, Y.X.; Lan, M.; Zhang, T.R.; Zhou, J. Protective Effects of Leukemia Inhibitory Factor on Retinal Vasculature and Cells in Streptozotocin-induced Diabetic Mice. Chin. Med. J. 2018, 131, 75–81. [Google Scholar] [CrossRef]
- Escary, J.L.; Perreau, J.; Dumenil, D.; Ezine, S.; Brulet, P. Leukaemia inhibitory factor is necessary for maintenance of haematopoietic stem cells and thymocyte stimulation. Nature 1993, 363, 361–364. [Google Scholar] [CrossRef]
- Schultze, W.H. Über das Paraphenylendiamin in der histologischen Färbetechnik und über eine neue Schnellfärbemethode der Nervenmarkscheiden am Gefrierschnitt. Zent. Pathol. 1917, 36, 639–640. [Google Scholar]
- Richardson, K.C.; Jarett, L.; Finke, E.H. Embedding in epoxy resins for ultrathin sectioning in electron microscopy. Stain Technol. 1960, 35, 313–323. [Google Scholar] [CrossRef]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Kubota, Y.; Hirashima, M.; Kishi, K.; Stewart, C.L.; Suda, T. Leukemia inhibitory factor regulates microvessel density by modulating oxygen-dependent VEGF expression in mice. J. Clin. Investig. 2008, 118, 2393–2403. [Google Scholar] [CrossRef] [Green Version]
- Pannicke, T.; Wagner, L.; Reichenbach, A.; Grosche, A. Electrophysiological characterization of Muller cells from the ischemic retina of mice deficient in the leukemia inhibitory factor. Neurosci. Lett. 2018, 670, 69–74. [Google Scholar] [CrossRef]
- Barbaric, I.; Miller, G.; Dear, T.N. Appearances can be deceiving: Phenotypes of knockout mice. Brief. Funct. Genom. 2007, 6, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Sendtner, M.; Gotz, R.; Holtmann, B.; Escary, J.L.; Masu, Y.; Carroll, P.; Wolf, E.; Brem, G.; Brulet, P.; Thoenen, H. Cryptic physiological trophic support of motoneurons by LIF revealed by double gene targeting of CNTF and LIF. Curr. Biol. 1996, 6, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Leibinger, M.; Muller, A.; Andreadaki, A.; Hauk, T.G.; Kirsch, M.; Fischer, D. Neuroprotective and axon growth-promoting effects following inflammatory stimulation on mature retinal ganglion cells in mice depend on ciliary neurotrophic factor and leukemia inhibitory factor. J. Neurosci. 2009, 29, 14334–14341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, J.; Cayouette, M.; Gravel, C. The CNTF/LIF signaling pathway regulates developmental programmed cell death and differentiation of rod precursor cells in the mouse retina in vivo. Dev. Biol. 2006, 300, 583–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgi, S.; Samardzija, M.; Grimm, C. Endogenous leukemia inhibitory factor protects photoreceptor cells against light-induced degeneration. Mol. Vis. 2009, 15, 1631–1637. [Google Scholar] [PubMed]
- Brandstatter, J.H.; Hartveit, E.; Sassoe-Pognetto, M.; Wassle, H. Expression of NMDA and high-affinity kainate receptor subunit mRNAs in the adult rat retina. Eur. J. Neurosci. 1994, 6, 1100–1112. [Google Scholar] [CrossRef]
- Grunder, T.; Kohler, K.; Kaletta, A.; Guenther, E. The distribution and developmental regulation of NMDA receptor subunit proteins in the outer and inner retina of the rat. J. Neurobiol. 2000, 44, 333–342. [Google Scholar] [CrossRef]
- Luo, X.; Baba, A.; Matsuda, T.; Romano, C. Susceptibilities to and mechanisms of excitotoxic cell death of adult mouse inner retinal neurons in dissociated culture. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4576–4582. [Google Scholar] [CrossRef]
- Ullian, E.M.; Barkis, W.B.; Chen, S.; Diamond, J.S.; Barres, B.A. Invulnerability of retinal ganglion cells to NMDA excitotoxicity. Mol. Cell Neurosci. 2004, 26, 544–557. [Google Scholar] [CrossRef]
- Fischer, A.J.; Schmidt, M.; Omar, G.; Reh, T.A. BMP4 and CNTF are neuroprotective and suppress damage-induced proliferation of Muller glia in the retina. Mol. Cell Neurosci. 2004, 27, 531–542. [Google Scholar] [CrossRef]
- Flachsbarth, K.; Jankowiak, W.; Kruszewski, K.; Helbing, S.; Bartsch, S.; Bartsch, U. Pronounced synergistic neuroprotective effect of GDNF and CNTF on axotomized retinal ganglion cells in the adult mouse. Exp. Eye Res. 2018, 176, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, A.; Iandiev, I.; Pannicke, T.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Osborne, N.N.; Reichenbach, A. Cellular signaling and factors involved in Muller cell gliosis: Neuroprotective and detrimental effects. Prog. Retin. Eye Res. 2009, 28, 423–451. [Google Scholar] [CrossRef] [PubMed]
- LaVail, M.M.; Unoki, K.; Yasumura, D.; Matthes, M.T.; Yancopoulos, G.D.; Steinberg, R.H. Multiple growth factors, cytokines and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc. Natl. Acad. Sci. USA 1992, 89, 11249–11253. [Google Scholar] [CrossRef] [Green Version]
- LaVail, M.M.; Yasumura, D.; Matthes, M.T.; Lau-Villacorta, C.; Unoki, K.; Sung, C.H.; Steinberg, R.H. Protection of mouse photoreceptors by survival factors in retinal degenerations. Investig. Ophthalmol. Vis. Sci. 1998, 39, 592–602. [Google Scholar]
- Leopold, S.A.; Zeilbeck, L.F.; Weber, G.; Seitz, R.; Bosl, M.R.; Jagle, H.; Fuchshofer, R.; Tamm, E.R.; Ohlmann, A. Norrin protects optic nerve axons from degeneration in a mouse model of glaucoma. Sci. Rep. 2017, 7, 14274. [Google Scholar] [CrossRef] [Green Version]
- Graf, U.; Casanova, E.A.; Cinelli, P. The Role of the Leukemia Inhibitory Factor (LIF)—Pathway in Derivation and Maintenance of Murine Pluripotent Stem Cells. Genes 2011, 2, 280–297. [Google Scholar] [CrossRef] [Green Version]
- Todd, L.; Palazzo, I.; Suarez, L.; Liu, X.; Volkov, L.; Hoang, T.V.; Campbell, W.A.; Blackshaw, S.; Quan, N.; Fischer, A.J. Reactive microglia and IL1beta/IL-1R1-signaling mediate neuroprotection in excitotoxin-damaged mouse retina. J. Neuroinflamm. 2019, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Tsoka, P.; Barbisan, P.R.; Kataoka, K.; Chen, X.N.; Tian, B.; Bouzika, P.; Miller, J.W.; Paschalis, E.I.; Vavvas, D.G. NLRP3 inflammasome in NMDA-induced retinal excitotoxicity. Exp. Eye Res. 2019, 181, 136–144. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (Forward) | Sequence (Reverse) |
|---|---|---|
| Mouse primers | ||
| Edn2 | 5′-ACCTCCTCCGAAAGCTGAG-3′ | 5′-TTTCTTGTCACCTCTGGCTGTA-3° |
| Fgf2 | 5′-CGGCTCTACTGCAAGAACG-3′ | 5′-TGCTTGGAGTTGTAGTTTGACG-3′ |
| Gfap | 5′-ACAGACTTTCTCCAACCTCCAG-3′ | 5′-CCTTCTGACACGGATTTGGT-3′ |
| Gnb2l | 5′-TCTGCAAGTACACGGTCCAG-3′ | 5′-GAGACGATGATAGGGTTGCTG-3′ |
| Lif | 5′-AAACGGCCTGCATCTAAGG-3′ | 5′-AGCAGCAGTAAGGGCACAAT-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kassumeh, S.; Leopold, S.; Fuchshofer, R.; Thomas, C.N.; Priglinger, S.G.; Tamm, E.R.; Ohlmann, A. Norrin Protects Retinal Ganglion Cells from Excitotoxic Damage via the Induction of Leukemia Inhibitory Factor. Cells 2020, 9, 277. https://doi.org/10.3390/cells9020277
Kassumeh S, Leopold S, Fuchshofer R, Thomas CN, Priglinger SG, Tamm ER, Ohlmann A. Norrin Protects Retinal Ganglion Cells from Excitotoxic Damage via the Induction of Leukemia Inhibitory Factor. Cells. 2020; 9(2):277. https://doi.org/10.3390/cells9020277
Chicago/Turabian StyleKassumeh, Stefan, Stephanie Leopold, Rudolf Fuchshofer, Carina N. Thomas, Siegfried G. Priglinger, Ernst R. Tamm, and Andreas Ohlmann. 2020. "Norrin Protects Retinal Ganglion Cells from Excitotoxic Damage via the Induction of Leukemia Inhibitory Factor" Cells 9, no. 2: 277. https://doi.org/10.3390/cells9020277