LIMD2 Regulates Key Steps of Metastasis Cascade in Papillary Thyroid Cancer Cells via MAPK Crosstalk

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Cell Line and Culture
2.3. Western Blotting
2.4. RNA Extraction, cDNA Synthesis, and Quantitative RT-PCR
2.5. LIMD2 Copy Number Analysis by qPCR
2.6. Generation of Stable LIMD2 Knockout (KO) Papillary Thyroid Cell Lines with CRISPR/Cas9 System
2.7. Analysis of the KO of LIMD2 in Thyroid Cells by PCR Sequencing
2.8. Tracking of Indels by Decomposition (TIDE) Analysis
2.9. Isolation and Expansion of CRISPR/Cas9 Edited Single Clones Derived from BCPAP and TPC1 Cells
2.10. Analysis of the LIMD2 Editing Efficiencies Using Western Blot, Flow Cytometry, and High-Content Screening
2.11. Off-Target Analysis
2.12. Ultrastructure of Parental and Edited LIMD2 Cells via High Resolution Imaging Analysis
2.13. Monitoring Cell Invasion
2.14. Proteome Profiler Antibody Array
2.15. Statistical Analysis
3. Results
3.1. LIMD2 is Specifically Expressed in PTC Cells
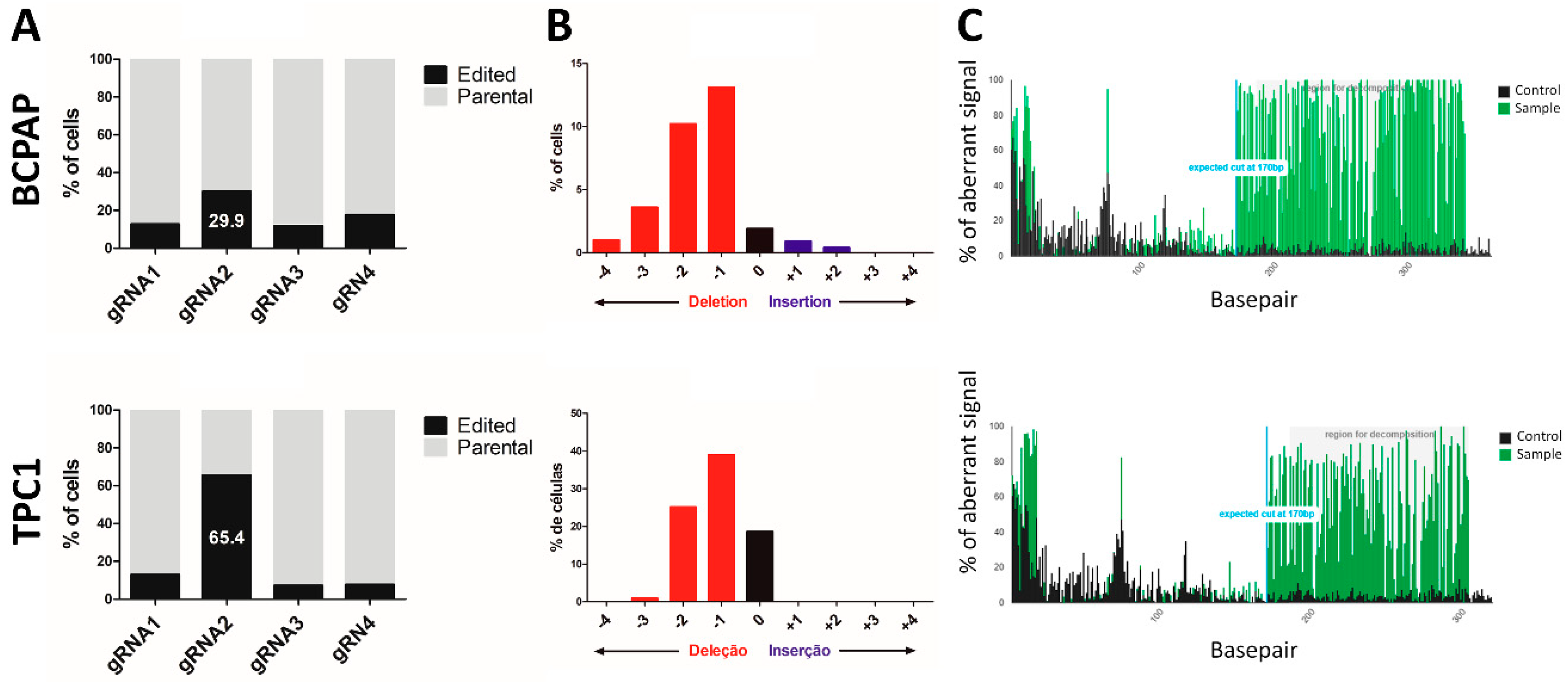
3.2. CRISPR/Cas9 System Successfully Promoted the LIMD2 KO in Thyroid Carcinoma Cell Lines
3.3. No Altered Sequence was Found in any of the Potential Off-Target Genes
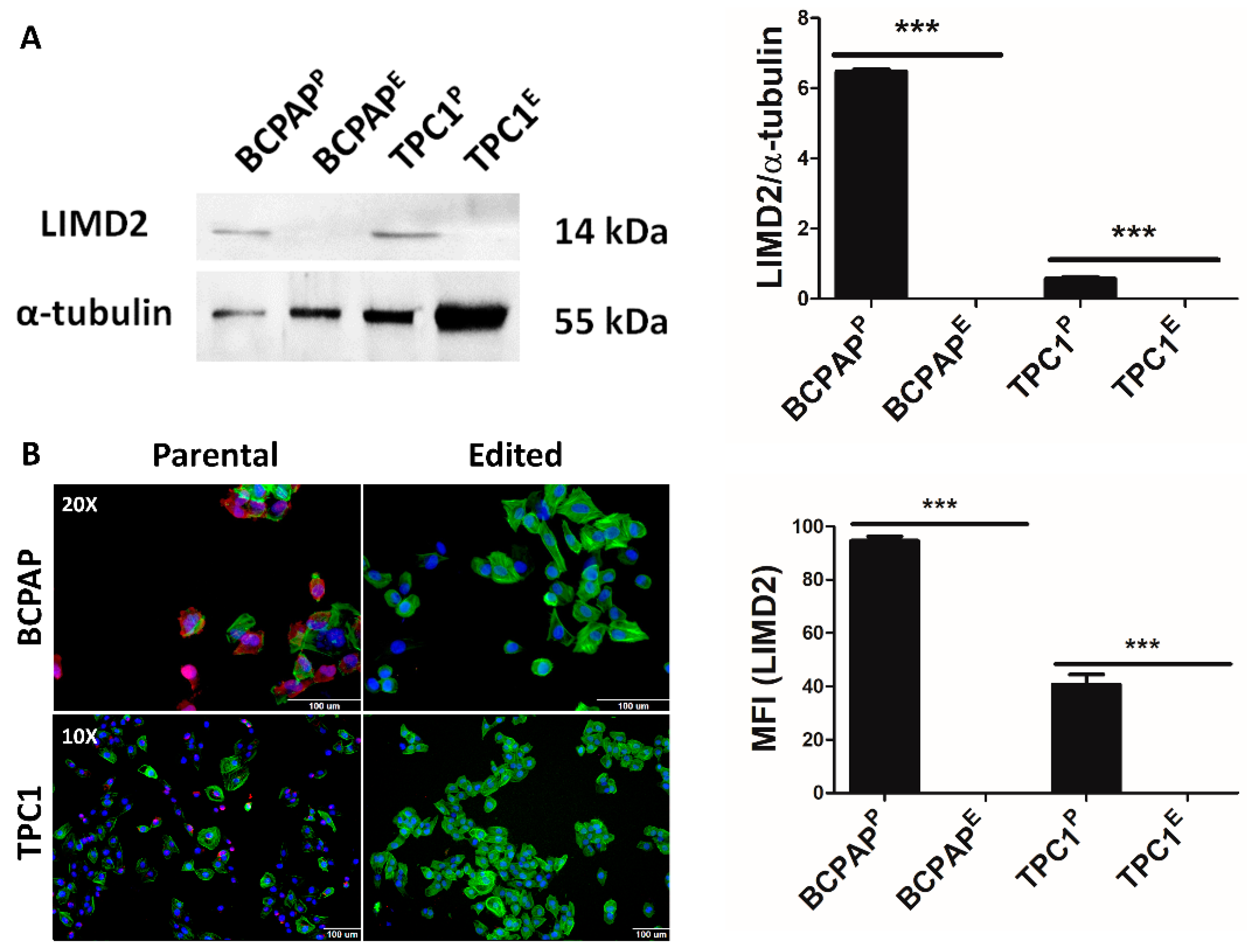
3.4. CRISPR/Cas9 System Promoted Functional KO of LIMD2
3.5. LIMD2 KO Restored Thyroid Cell Polarity and Mitochondria Morphology
3.6. LIMD2 KO Reduces the Cell Invasion
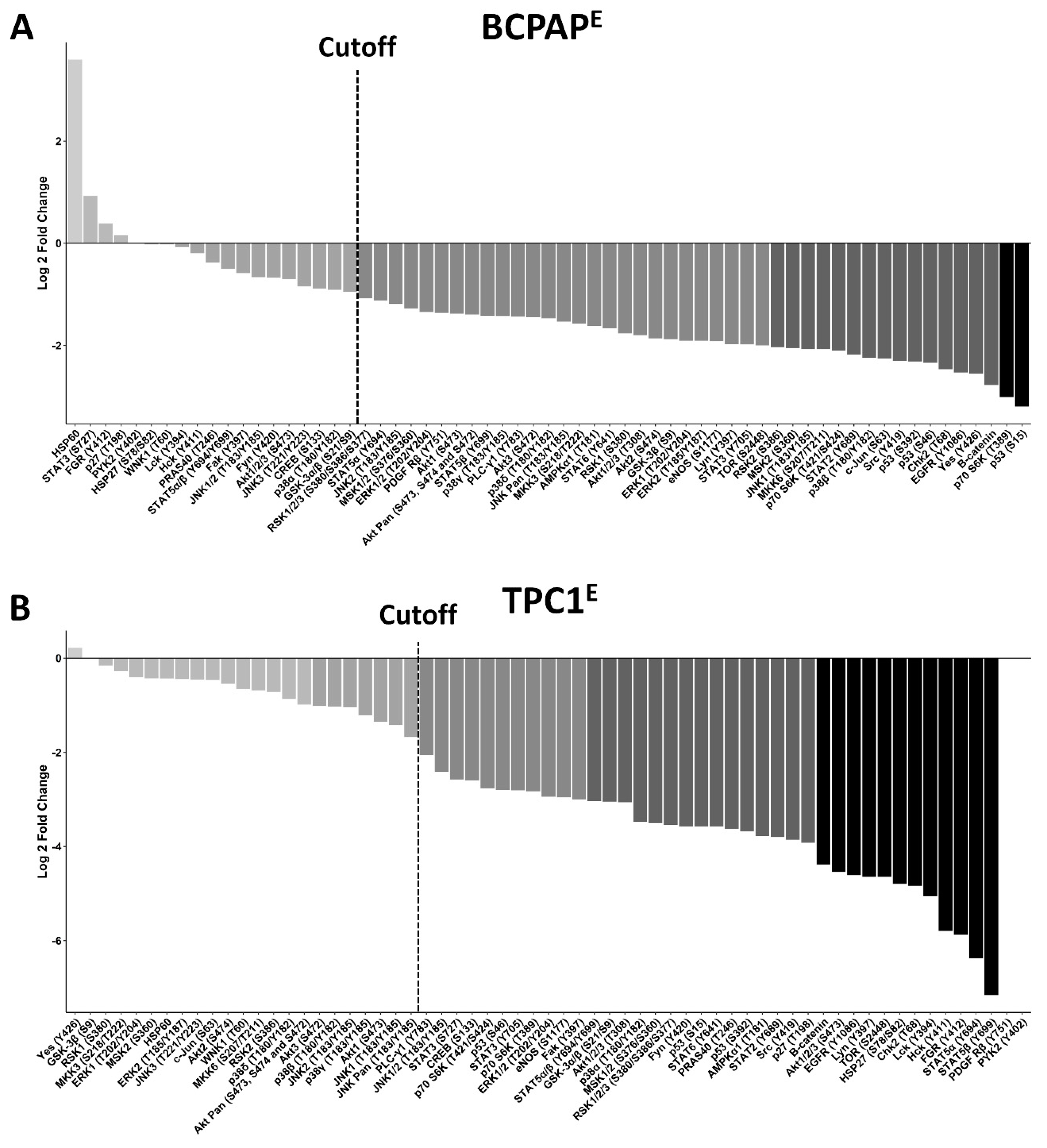
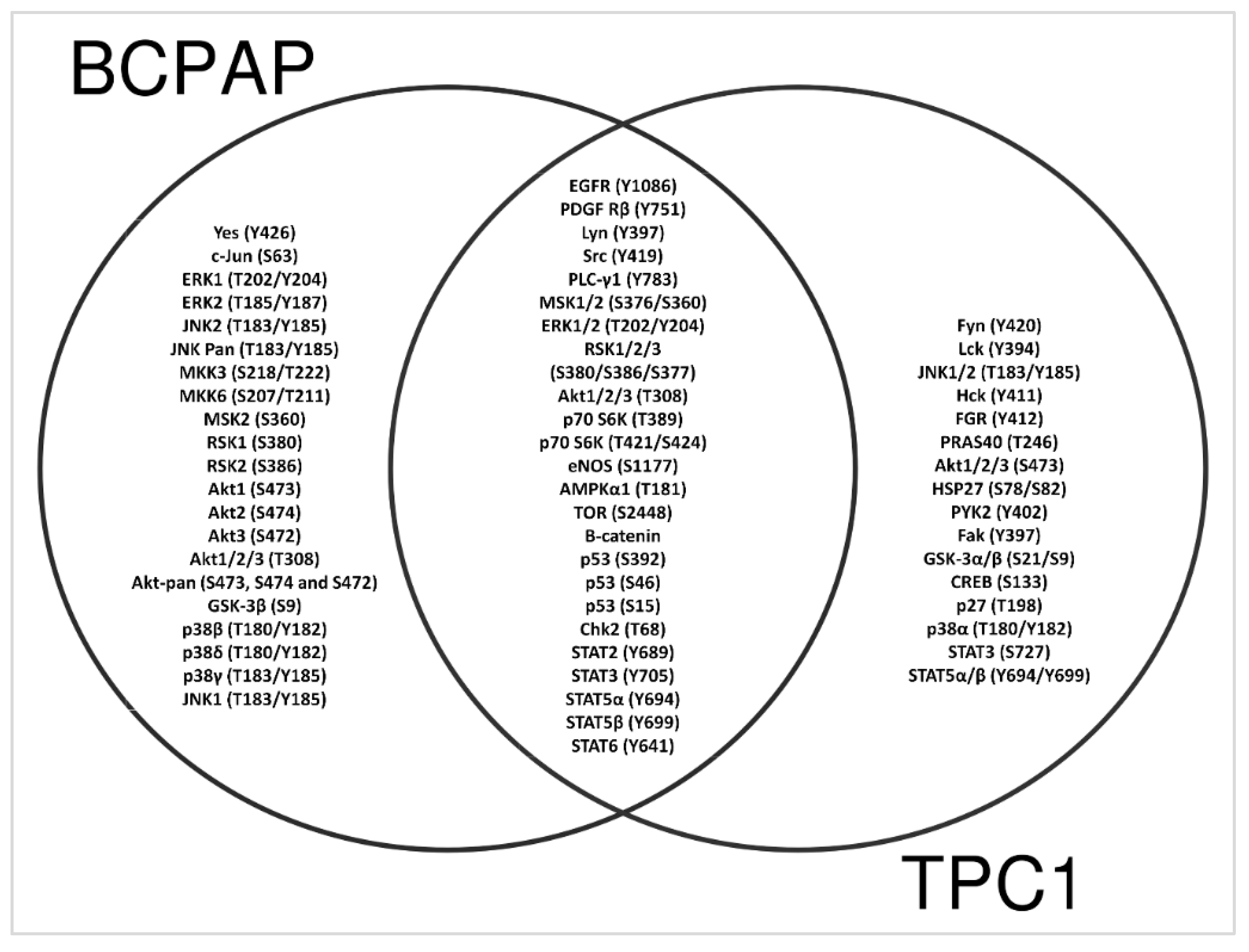
3.7. Decreased Serine/Threonine Kinase Phosphorylation in LIMD2 KO Cells
3.8. LIMD2 KO Reduces the Expression of Proteins Associated with Epithelial Mesenchymal Transition
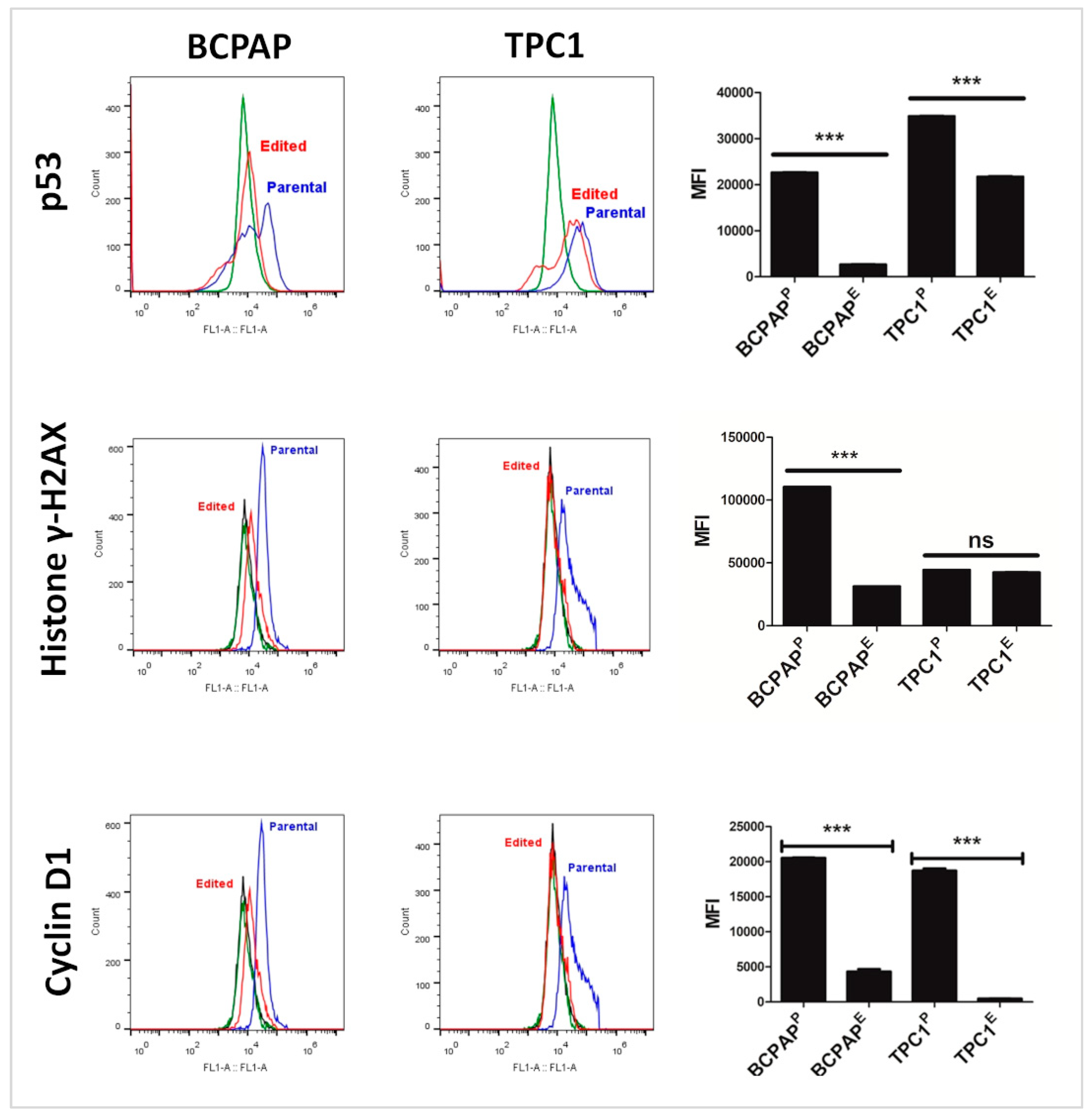
3.9. KO of LIMD2 Reduced Phosphorylation of Proteins that Function as a Guardian of the Genome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Maksimovic, S.; Jakovljevic, B.; Gojkovic, Z. Lymph node metastases papillary thyroid carcinoma and their importance in recurrence of disease. Med. Arch. 2018, 72, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohec, H.; Breuskin, I.; Hadoux, J.; Schlumberger, M.; Leboulleux, S.; Hartl, D.M. Occult contralateral lateral lymph node metastases in unilateral N1b papillary thyroid carcinoma. World J. Surg. 2019, 43, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Pazaitou-Panayiotou, K.; Kaprara, A.; Chrisoulidou, A.; Boudina, M.; Georgiou, E.; Patakiouta, F.; Drimonitis, A.; Vainas, I. Cerebellar metastasis as first metastasis from papillary thyroid carcinoma. Endocr. J. 2005, 52, 653–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid 2016, 26, 1–133. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, R.M.; Haugen, B.; Perrier, N.D. Updated American Joint Committee on Cancer/Tumor-Node-Metastasis Staging System for Differentiated and Anaplastic Thyroid Cancer (Eighth Edition): What Changed and Why? Thyroid 2017, 27, 751–756. [Google Scholar] [CrossRef]
- Cerutti, J.; Oler, G.; Michaluart, P.; Delcelo, R.; Beaty, R.; Shoemaker, J.; Riggins, G. Molecular profiling of matched samples identifies biomarkers of papillary thyroid carcinoma lymph node metastasis. Cancer Res. 2007, 67, 7885–7892. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.J.C.P.; Bastos, A.U.; Costa, V.R.; Delcelo, R.; Lindsey, S.C.; Colozza-Gama, G.A.; Peng, H.; Rauscher, F.J.; Oler, G.; Cerutti, J.M. LIMD2 Is Overexpressed in BRAF V600E-Positive Papillary Thyroid Carcinomas and Matched Lymph Node Metastases. Endocr. Pathol. 2018, 29, 222–230. [Google Scholar] [CrossRef]
- Peng, H.; Talebzadeh-Farrooji, M.; Osborne, M.; Prokop, J.; McDonald, P.; Karar, J.; Hou, Z.; He, M.; Kebebew, E.; Orntoft, T.; et al. LIMD2 is a small lim-only protein overexpressed in metastatic lesions that regulates cell motility and tumor progression by directly binding to and activating the integrin-linked kinase. Cancer Res. 2014, 74, 1390–1403. [Google Scholar] [CrossRef] [Green Version]
- Araldi, R.P.; Cerutti, J.M. Novel biotechnological opportunities in thyroid cancer metastasis based on LIMD2 differential expression. Trend Cancer Res. Chemother. 2018, 1, 1–6. [Google Scholar] [CrossRef]
- Koch, B.J.; Ryan, J.F.; Baxevanis, A.D. The diversification of the lim superclass at the base of the metazoa increased subcellular complexity and promoted multicellular specialization. PLoS ONE 2012, 7, e33261. [Google Scholar] [CrossRef] [Green Version]
- Landa, I.; Pozdeyev, N.; Korch, C.; Marlow, L.A.; Smallridge, R.C.; Copland, J.A.; Henderson, Y.C.; Lai, S.Y.; Clayman, G.L.; Onoda, N.; et al. Comprehensive genetic characterization of human thyroid cancer cell lines: A validated panel for preclinical studies. Clin. Cancer Res. 2019, 25, 3141–3151. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.N.L.; Hemerly, J.P.; Bastos, A.U.; Tamanaha, R.; Latini, F.R.M.; Camacho, C.P.; Impellizzeri, A.; Maciel, R.M.B.; Cerutti, J.M. The RET p.G533C mutation confers poredisposition to multiple endocrine neoplasia type 2A in a Brazilian kindred and is able to induce a malignant phenotype in vitro and in vivo. Thyroid 2011, 21, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Moraes, L.; Zanchin, N.I.T.; Cerutti, J.M. ABI3, a component of the WAVE2 complex, is potentially regulated by PI3K/AKT pathway. Oncotarget 2017, 8, 67769–67781. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Desai, S.P.; Bhatia, S.N.; Toner, M.; Irimia, D.; Chase, C. Mitochondrial localization and the persistent migration of epithelial cancer cells. Biophys. J. 2013, 104, 2077–2088. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Li, Z.; Xu, L.; Li, Y.; Li, Y.; Zhang, X.; Wang, Y.; Liu, D. LIMD2 targeted by miR‑34a promotes the proliferation and invasion of non‑small cell lung cancer cells. Mol. Med. Rep. 2018, 18, 4760–4766. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Qin, S.; Xiao, X.; Tan, Y.; Hao, P.; Xu, Y. Overexpression of LIMD2 promotes the progression of non-small cell lung cancer. Oncol. Lett. 2019, 18, 2073–2081. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Hu, H.; Hong, P.; Zhang, Q.; Xu, W.W.; He, Q.; Li, B. Significance of integrin-linked kinase ( ILK ) in tumorigenesis and its potential implication as a biomarker and therapeutic target for human cancer. Am. J. Cancer Res. 2019, 9, 186–197. [Google Scholar] [PubMed]
- Shirley, L.A.; McCarty, S.; Yang, M.C.; Saji, M.; Zhang, X.; Phay, J.; Ringel, M.D.; Chen, C.S. Integrin-linked kinase affects signaling pathways and migration in thyroid cancer cells and is a potential therapeutic target. Surgery 2016, 159, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Huang, C. Regulation of EMT by STAT3 in gastrointestinal cancer (Review). Int. J. Oncol. 2017, 50, 753–767. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Ren, C.; Wang, J.; Wang, S.; Yang, L.; Han, X.; Chen, Y.; Tong, G.; Yang, G. The crosstalk between STAT3 and p53/RAS signaling controls cancer cell metastasis and cisplatin resistance via the Slug/MAPK/PI3K/AKT-mediated regulation of EMT and autophagy. Oncogenesis 2019, 8, 59. [Google Scholar] [CrossRef] [Green Version]
- Medici, D.; Hay, E.; Olsen, B. Snail and Slug promote epithelial-mesenchymal transition through B-catenin-T-cell factor-4-dependent expression of transforming growth fator-B3. Mol. Biol. Cell 2008, 19, 4875–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visciano, C.; Liotti, F.; Prevete, N.; Cali’, G.; Franco, R.; Collina, F.; de Paulis, A.; Marone, G.; Santoro, M.; Melillo, R.M. Mast cells induce epithelial-to-mesenchymal transition and stem cell features in human thyroid cancer cells through an IL-8–Akt–Slug pathway. Oncogene 2015, 34, 5175–5186. [Google Scholar] [CrossRef] [PubMed]
- Buehler, D.; Hardin, H.; Shan, W.; Montemayor-Garcia, C.; Rush, P.; Asioli, S.; Chen, H.; Lloyd, R. Expression of epithelial-mesenchymal transition regulators SNAI2 and TWIST1 in thyroid carcinomas. Mod. Pathol. 2013, 26, 54–61. [Google Scholar] [CrossRef]
- Gasparotto, D.; Polesel, J.; Marzotto, A.; Colladel, R.; Piccinin, S.; Modena, P.; Grizzo, A.; Sulfaro, S.; Barzan, L.; Doglioni, C.; et al. Overexpression of TWIST2 correlates with poor prognosis in head and neck squamous cell carcinomas. Oncotarget 2011, 2, 1165–1175. [Google Scholar] [CrossRef] [Green Version]
- Bremnes, R.; Veve, R.; Hirsch, F.; Franklin, W. The E-cadherin cell–cell adhesion complex and lung cancer invasion, metastasis, and prognosis. Lung Cancer 2002, 36, 115–124. [Google Scholar] [CrossRef]
- Hajra, K.; Chen, D.; Fearon, E. Advances in brief the SLUG zinc-finger protein represses E-cadherin in breast cancer 1. Cancer Res. 2002, 62, 1613–1618. [Google Scholar]
- Cox, M.L.; Meek, D.W. Phosphorylation of serine 392 in p53 is a common and integral event during p53 induction by diverse stimuli. Cell. Signal. 2010, 22, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Maclaine, N.J.; Hupp, T.R. The regulation of p53 by phosphorylation: A model for how distinct signals integrate into the p53 pathway. Aging 2009, 1, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX phosphorylation: A marker for DNA damage. Methods Mol. Biol. 2012, 920, 613–626. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Histological Type | Medium | Source | Mutation Identified 1 |
|---|---|---|---|---|
| BCPAP | PTC | RPMI 1640 medium, supplemented with 5% FBS | DSMZ Cat No. ACC 273 | BRAF V600E, TP53 D258Y, TERT c.-124C>T and c.-125C>T |
| TPC1 | PTC | RPMI 1640 medium, supplemented with 10% FBS | João R. Maciel, UNIFESP, SP, Brazil | RET/PTC1, TERT c.-124C>T |
| FTC133 | FTC | DMEM and Ham’s F12 (1:1) medium, supplemented with 10% FBS | ECACC cat no. 94060901 | NF1 C167*, PTEN R130*, TP53 R273H, TERT c.-124C>T, RB1*, MSH6* |
| FTC236 | FTC | DMEM and Ham’s F12 (1:1) medium, supplemented with 10% FBS, 1 mU/mL TSH, and 10 µg/mL insulin | ECACC cat. no. 06030202 | PTEN R130*, TP53 R273H, TERT c.-124C>T, RB1*, MSH6* and others |
| FTC238 | FTC | DMEM and Ham’s F12 (1:1) medium, supplemented with 10% FBS | ECACC cat no. 94060902 | PTEN R130*, TP53 R273H, TERT c.-124C>T, RB1*, MSH6* |
| WRO | FTC | DMEM medium, supplemented with 10% FBS | Alfredo Fusco, Università degli Studi di Napoli Federico II, Italy | TP53 P223L, BRAF wild type, TERT promoter wild type |
| TT | MTC | F-12K medium, supplemented with 10% FBS | ECACC cat no. 92050721 | RET C634W |
| Target | m/pAb | Specie | Source | Cat. Number | Dilution | Assay |
|---|---|---|---|---|---|---|
| LIMD2 1 | pAb | Rabbit | Peng et al. [8] | - | 1: 1000 | WB |
| LIMD2 2 | pAb | Rabbit | Abcam | Ab221110 | 1:33 | WB, HCS |
| Α-tubulin | mAb | Mouse | Sigma-Aldrich | T9026 | 1:10,000 | WB |
| F-actin 3 | - | - | Thermo Fisher Scientific | A12379 | 1:300 | HCS |
| Histone-γH2AX | pAb | Rabbit | Novus Biologicals | NB100-384 | 1:100 | FC |
| P53 | pAb | Rabbit | Abcam | ab131442 | 1:100 | FC |
| β-catenin | mAb | Mouse | Santa Cruz Biotechnology | Sc-376841 | 1:100 | FC, HCS |
| STAT3 (Y705) | pAb | Rabbit | R&D Systems | AF4607 | 1:100 | FC, HCS |
| STAT5α | mAb | Mouse | Santa Cruz Biotechnology | Sc-166479 | 1:200 | FC, HCS |
| SLUG | pAb | Rabbit | Santa Cruz Biotechnology | sc-15391 | 1:100 | FC, HCS |
| TWIST2 | mAb | Mouse | Abcam | Ab50887 | 1:50 | FC, HCS |
| N-cadherin | mAb | Mouse | Dako | M3613 | 1:100 | FC, HCS |
| FITC-conjugated goat anti-rabbit IgG | pAb | Goat | Thermo Fisher Scientific | AB42978 | 1:1000 | FC, HCS |
| FITC-conjugated goat anti-mouse IgG | pAb | Goat | Thermo Fisher Scientific | AB2533946 | 1:10,000 | FC, HCS |
| gRNA | crRNA | Off-Targets | Exon |
|---|---|---|---|
| 1 | GGCAACTACGACGAGGGGTTTGG | 0 | 5 |
| 2 | CCAGAAGACCGTGTACCCCATGG | 5 | 4 |
| 3 | GTTTGGCCGCAAGCAGCACAAGG | 2 | 5 |
| 4 | AGCAAAGGCAACTACGACGAAGG | 2 | 5 |
| Sequence | PAM | Coordinate | Gene | Locus |
|---|---|---|---|---|
| CCAAAAGACCGGATACCCCA | CGG | chr15[74825883] | - | - |
| CCAGAAGACCGTGCAGCCCC | TGG | chrX[132498708] | - | - |
| GCAGAAGTCCGTGAACCCCA | TGG | chr3[153321832] | - | - |
| CCAGAAGTCTGTGTTCCCCA | TGG | chr13[24452558] | PARP4 | 13q12.12 |
| CCAGGAGACCCTGGACCCCA | AGG | chr6[70199715] | COL19A1 | 6q13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araldi, R.P.; de Melo, T.C.; Levy, D.; de Souza, D.M.; Maurício, B.; Colozza-Gama, G.A.; Bydlowski, S.P.; Peng, H.; Rauscher, F.J., III; Cerutti, J.M. LIMD2 Regulates Key Steps of Metastasis Cascade in Papillary Thyroid Cancer Cells via MAPK Crosstalk. Cells 2020, 9, 2522. https://doi.org/10.3390/cells9112522
Araldi RP, de Melo TC, Levy D, de Souza DM, Maurício B, Colozza-Gama GA, Bydlowski SP, Peng H, Rauscher FJ III, Cerutti JM. LIMD2 Regulates Key Steps of Metastasis Cascade in Papillary Thyroid Cancer Cells via MAPK Crosstalk. Cells. 2020; 9(11):2522. https://doi.org/10.3390/cells9112522
Chicago/Turabian StyleAraldi, Rodrigo Pinheiro, Thatiana Correa de Melo, Débora Levy, Dener Madeiro de Souza, Beatriz Maurício, Gabriel Avelar Colozza-Gama, Sergio Paulo Bydlowski, Hongzhuang Peng, Frank J. Rauscher, III, and Janete Maria Cerutti. 2020. "LIMD2 Regulates Key Steps of Metastasis Cascade in Papillary Thyroid Cancer Cells via MAPK Crosstalk" Cells 9, no. 11: 2522. https://doi.org/10.3390/cells9112522