1. Introduction
The metabolic syndrome, which comprises obesity and diabetes, is a major public health problem and the comprehension of energy homeostasis control remains an important worldwide issue. The energy balance is finely regulated by the central nervous system (CNS), notably through neuronal networks, located in the hypothalamus and the DVC including the nucleus tractus solitarius (NTS), which integrate nutritional, humoral and nervous information from the periphery and in turn adjust energy expenditure and food intake. Decoding the precise mechanisms regulating energy balance remains a challenge necessary to efficiently treat metabolic disorders. Evidence showing contribution of glial cells in these processes are now mounting. Indeed, recent studies showed that glial cells in both the hypothalamus [
1,
2,
3] and brainstem [
4,
5,
6] interact with neurons and in turn finely tune energy homeostasis. However, the mechanisms of neuron-glia interactions in the context of food intake control remain to be deciphered. A typical feature of glial cells is their high expression level of Cx43, a protein able to form not only gap junction (GJ) channels, but also connexin 43 hemichannels (Cx43 HCs), in allowing the release of neuroactive substances.
Cxs proteins are highly conserved between species and are named according to their molecular weight, from 26 to 60 kDa. There are around 20 different genes (20 in mice and 21 in humans) coding for as many proteins [
7]. The Cxs are the connexon base subunits, a hexamer forming a hydrophilic channel 2 nm in diameter. When two connexons of different cells join together, they form an intercellular channel and several thousands of such channels form a GJ [
8]. In the central nervous system (CNS), glia and particularly astrocytes primarily and strongly express Cx43, Cx30 and to a lesser extent Cx26 [
9]. These Cxs GJ contribute to a cytoplasmic continuity, providing the structural basis for an extensive astroglial network [
10]. Alternatively, connexons not engaged in GJ form hemichannels (HCs), which are permeable to small molecules (less than 1.5 kDa) such as Ca
2+, glucose, adenosine tri-phosphate (ATP), D-serine and glutamate [
11]. The opening of these HCs is regulated by different factors such as Ca
2+, pH, hormones [
9]. The study of the functional role of these HCs is now possible thanks to the development of peptides specifically blocking their activity, such as TAT-GAP19, a mimetic peptide preventing interaction between the C-terminal part of Cx43 with the L2 loop [
12]. HCs contribute to the release of various neuroactive molecules and can thus modulate of neuronal networks and associated behaviors [
13,
14,
15,
16].
The aim of the present study was to determine the possible contribution of glial HCs in rodent’s energy balance. For this, we have characterized Cx43 expression and its variations during nutritional status changes in mouse hypothalamus and brainstem. Next, we investigated the structural relationship between Cx43 and synaptic elements in these brain regions and studied the impact of Cx43 HCs on energy balance and activation of food-intake-dedicated neuronal networks using pharmacological inhibition.
2. Materials and Methods
2.1. Animals
Adult male C57BL/6J mice were purchased from Charles River Laboratories (L’Abresle, France). For TAT-GAP19 or mutated TAT-GAP19-I14A injections, C57BL/6J adult male mice (8 to 10 weeks old) were housed in standard cages, maintained in a controlled environment (12/12 h light-dark cycle, 22 °C and 40–50% humidity) with free access to water and food (AO4 pellets, SAFE, Augy, France). Some mice received a high fat diet (HFD, 60% fat; SSNIFF GmbH, Germany). Body weight and food intake were measured every week. In the HFD group, only mice which gained more than 30% of weight mass after 15 weeks compared to mice fed with the standard food (NC) were considered as obese and conserved for the tissue analyzes. Both obese and lean mice were sacrificed after 15 weeks of diet and tissues samples were collected for further analysis. All experiments were conformed to EC Council Directive (2010/63/UE) and the French “Direction Départementale de la Protection des Populations des Bouches-du-Rhône” (animal housing facility accreditation: N° D13 0556; animal experimentation accreditation: APAFIS#2207-2015100819154639 v4).
2.2. Peptides
TAT-GAP19 and TAT-GAP19-I14A were synthesized by Genopshere Biotechnologies using the following amino acid sequences: YGRKKRRQRRRKQIEIKKFK. The mutated and noneffective peptide was strictly identical except for Alanine at 14th position instead of Isoleucine (mutation I130A in Cx43 protein). TAT peptide was added to GAP19 to allow for its cells penetration. To detect these peptides using histology, we used the same sequences coupled with N-terminal biotin.
2.3. Surgery and Intracerebroventricular Injection
Cannula implantation was performed as previously described by Gaigé and colleagues [
17]. One week post-surgery, mice were i.c.v. injected with 6 µL (2.5 µL/min) of physiological saline (NaCl 0.9%), TAT-GAP19 (12.5, 25 or 50 µg; 6 µL), or TAT-GAP19-I14A (25 µg) solutions at the beginning of the dark phase.
For acute injection of biotinylated TAT-GAP19 and TAT-GAP19-I14A, mice were anaesthetized by intraperitoneal (i.p.) injection of ketamine (100 mg/kg) and xylazine (16 mg/kg). Under stereotaxic control, mice were then i.c.v. injected with 6 µL (2.5 µL/min) of physiological saline (NaCl 0.9%), TAT-GAP19 (25 µg) or TAT-GAP19-I14A (25 µg) solutions. At different times after biotinylated mimetic peptides injection, anaesthetized animals were perfused intracardially with ice-cold 0.1M phosphate buffered saline (PBS, pH 7.4) and then with ice-cold, freshly prepared 4% paraformaldehyde (PFA) in 0.1M PB. The brains were immediately removed, post-fixed for 1h in 4% PFA at room temperature, rinsed overnight in PBS and then cryoprotected for 24–48 h in 30% sucrose at 4 °C. The brains were frozen in isopentane (−40 °C), and coronal sections (40 µm thick) of the brain were made with a cryostat (Leica CM3050, Leica Biosystems, Wetzlar, Germany) and collected serially in 0.1M PBS. Immunochemistry of biotin was then performed as described below.
2.4. Food Intake Measurements
For food consumptions, ad libitum fed mice received i.c.v. injections of NaCl (0.9%), TAT-GAP19 (12.5 or 25 µg) or TAT-GAP19-I14A (25 µg) 30 min prior to lights-off. Immediately after treatment, a fresh supply of pre-weighed powdered food was given. Continuous food intake measurements and meal pattern analyses following i.c.v. injections were performed using a previously described feeding station [
17]. Food intake was calculated as the difference between the pre-weighed and the remaining powder measured with a precision balance (0.01 g; Denver Instrument from Bioblock, San Diego, CA, USA). To analyze the meal pattern, we modified polypropylene cages (Cage S; Charles River Laboratories, Wilmington, MA, USA) with a feeding station allowing access to a food jar placed below the cage, and filled with powdered food (AO4). The feeding station was designed to avoid feces and urine dropping into the food jar and also to avoid food spillage. Analysis first proceeded through a MATLAB-developed software, filtering out high frequency oscillations in food jar weights and collecting the start time, end time and weight amplitude of the stepwise reductions in food jar weights. Analyzed data were exported to Excel for further analysis. Meals were defined by stepwise reductions in food jar weight of at least 0.02 g in amplitude (i.e., twice the balance precision) and 30 s in duration.
2.5. Indirect Calorimetry and Locomotor Activity
Indirect calorimetry recording was performed with an indirect open-circuit calorimeter Oxylet Physiocage system (Panlab, Cornella, Spain), equipped with a laser absorption O2 sensor, an infrared technology CO2 sensor, and weight transducers for the continuous assessment of the food and water consumption. Room air flowed through each chamber at a rate of 450 mL/min. The O2 and CO2 levels were measured during 3 min sampling periods every 30 min and data were analyzed with the METABOLISM software. Oxygen consumption (VO2), and carbon dioxide production (VCO2) were expressed in mL/min/kg0.75. The respiratory quotient (RQ) was determined by the ratio VCO2/VO2 and energy expenditure (EE) was calculated according to the formula EE (kcal/day/kg0.75) = VO2 _ 1.44 _ [3.815 + (1.232 _ RQ)]. Mice were habituated to the metabolic chambers for 24 h. The O2/CO2 analyzer was calibrated with purified gas standards and the food hoppers replenished every day at the beginning of the light phase. TAT-GAP19 (50 µg) or vehicle was administered just before the dark phase, on day 2 of a 2-day recording session. The mean values for VO2, VCO2, respiratory quotient (RQ) and energy expenditure (EE) were compared for each group between the dark phase before and after treatment. To measure locomotor activity, cages were kept on weight transducers allowing the detection of animal movements (1 cumulative measurement per 30 min period, Arbitrary Unit, AU).
2.6. Oral Glucose Tolerance Test and Glycaemia Measurements
Glycaemia measurements were acquired by collecting blood samples from THE tail vein. A section of tail vein was pierced by a needle, and blood was directly applied to a strip for glucose detection using a glucose meter (Accu-Chek Performa nano, Roche Diagnostics Corporation, Indianapolis, IN, USA). Glycaemia was determined before the TAT-GAP19 injection and then at 10, 20, 30, 60, 90, 120, 180 min after its injection. For oral glucose tolerance test, i.c.v. TAT-GAP19 treatment or vehicle was performed 30 min before animals received a bolus of glucose per os (1.5 g/kg). Glycaemia was then determined before glucose administration and 5, 10, 20, 30, 60, 90, 120 and 180 min after its administration.
2.7. Tissue Collection
Mice were anaesthetized using i.p. injection of ketamine (100 mg/kg) and xylazine (16 mg/kg) and sacrificed by decapitation. The prefrontal cortex, hypothalamus and DVC were collected, rinsed in saline solution (NaCl 0.9%), snap-frozen in liquid nitrogen and stored at ‒80 °C until further analysis.
2.8. Real-Time PCR Analysis
Animals used for reverse transcriptase polymerase chain reaction (RT-PCR) analysis were sacrificed at the end of the treatment period. mRNA extraction and RT were made as described previously [
17]. Briefly, total RNAs were extracted from frozen organs using TRI Reagent
® (Sigma-Aldrich, St. Louis, MA, USA) according to the manufacturer’s instructions. RT was realized using Moloney Murine Leukemia Virus Reverse Transcriptase in the presence of random hexamer primers (Promega, Madison, WI, USA). Gene expression analysis by real time PCR was performed using the LightCycler
® 480 System (Roche Applied Science, Penzberg, Germany). The equivalent of 40 ng initial RNAs were subjected to PCR amplification in a 10 μL final volume using specific primers 0.25 μM (
Table 1) and KAPA SYBR
® FAST Master Mix (2X) optimized for Roche LightCycler
® 480 (CliniSciences, KAPABiosystems, Wilmington, MA, USA). The generation of specific PCR products was confirmed by melting-curve analysis. Glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) was used as internal reference gene.
2.9. Western Blotting
Hypothalamus tissues were homogenized in Radioimmunoprecipitation Assay (RIPA) buffer (50 mM Tris pH 8.0, 0.1% sodium dodecyl sulfate, 1% Triton X- 100, 0.5% sodium deoxycholate, 150 mM NaCl, and 1 mM EDTA) supplemented with protease and phosphatase inhibitors cocktail (
bimake.com, UK) then maintained under constant agitation in ice for 1 h. Extracts were centrifuged at 12,000×
g for 20 min at 4 °C to remove tissue debris. The supernatants were collected for assessment of protein concentration by Bicinchoninic Acid (BCA) Protein Assay method (Novagen, Germany). Then equivalent amounts of protein (30 μg) were separated on 12% SDS-PAGE gel and transferred onto nitrocellulose membranes (Amersham, Germany) to form blots.
Anti-connexin 43: The blots were blocked for 30 min with 2% casein in PBS- 0,1% Tween 20 and incubated for 2 h at room temperature with rabbit polyclonal antibody against mouse monoclonal anti-Cx43 antibody (1/13000; Sigma Aldrich, St. Louis, MO, USA). Blots were then incubated for 1 h at room temperature with anti-mouse immunoglobulin G (IgG) (Fab specific)-horseradish peroxidase (HRP) (1:2500; Sigma Aldrich, St. Louis, MO, USA).
Anti-phospho-connexin 43: The protein blots were blocked in 5% BSA in Tris-buffered saline Tween 20 (TBST) for 1h and then incubated with phospho-Cx43/GJA1 (Ser368) rabbit polyclonal antibody (1:1000 in TBST; ABCAM, UK) at 4 °C overnight, followed by incubation with HRP-conjugated goat anti-rabbit secondary antibody (1:10000; Bethyl, Montgomery, TX, USA) in 5% non-fat milk-TBST at room temperature for 1h. For loading control, blots were further stripped and re-probed with GAPDH mouse monoclonal antibody (1:10000; Proteintech, Rosemont, IL, USA) followed by HRP-conjugated goat anti-mouse secondary antibody (1:2500; Sigma Aldrich, St. Louis, MO, USA). Bands were visualized using the colorimetric system 3,3’,5,5’-tetramethylbenzidine (TMB) one-component HRP membrane substrate (SurModics, Eden Praire, MN, USA) and quantified by densitometry using ImageJ software (National Institute of Health, NIH, Bethesda, MD, USA).
2.10. Immunohistochemistry
Mice were anaesthetized using i.p. injection of ketamine (100 mg/kg) and xylazine (16 mg/kg) and then perfused intracardially with ice-cold 0.1M PBS pH 7.4 and then with ice-cold freshly prepared solution of 4% paraformaldehyde (PFA) in 0.1M PB. The brains were immediately removed, post-fixed for 1 h in 4% PFA at room temperature, rinsed overnight in PBS and then cryoprotected for 24–48 h in 30% sucrose at 4 °C. The brains were frozen in isopentane (‒40 °C) and coronal sections (40 µm thick) of the hypothalamus or brainstem were made with a cryostat (Leica CM3050, France) and collected serially in 0.1M PBS.
For Glial Fibrillary Acidic Protein (GAFP), Ionized Ca2+ Binding Adapter Molecule 1 (IBA1) and Cx43 immunostainings, sections were incubated for 1 h saturation buffer containing 0.3% triton X-100 in 1X PBS buffer and respectively 3% normal goat serum (NGS for GFAP), 3% horse serum (HS for IBA1) and 3% NGS coupled with 1% bovine serum albumin (NGS + BSA for Cx43). Primary antibodies against GFAP were used at 1/1000 (mouse #G3893, Sigma Aldrich, St. Louis, MO, USA), against IBA1 at 1/500 (rabbit #019-19741, Wako, Richmond, VA, USA) and against Cx43 at 1/3000 (rabbit #ACC-201, Alomone Labs, Jerusalem, Israel) or 1/2000 (mouse #C8093, Sigma Aldrich, St. Louis, MO, USA). After one night of incubation at 4 °C, labelings were revealed with corresponding secondary antibody coupled with Alexa Fluor 488 or 594 (Invitrogen, Carlsbad, CA, USA), used at 1/400 during 2 h incubation at room temperature.
Detection of biotinylated TAT-GAP19 and TAT-GAP19-I14A was performed after 1 h saturation of slices in 3% NGS and 0.3% triton PBS buffer and 2 h of incubation with avidine FITC (Vector) in the same PBS buffer.
For c-Fos immunohistochemistry, sections were incubated for 10 min in 0.1 M PBS containing 1.5% H2O2 for quenching of endogenous peroxidase activity. After 1 h in saturation PBS buffer containing 3% NGS and 0.3% triton X-100, sections were incubated for 48 h at 4 °C with a rabbit anti-c-Fos antibody (1/10 000, #PC38, Calbiochem). A biotinylated goat anti-rabbit IgG (1/400, #BA1000 Vector) was used as a secondary antibody (incubated for 1 h 30 at room temperature). Peroxidase activity was revealed using the avidin-biotin complex (1/200, Vector Labs, Burlingame, CA, USA) and diaminobenzidine as chromogen. Non-specific labelling was observed on adjacent sections that were treated identically but without the primary antibody. The reaction was closely monitored and terminated by washing the sections in distilled water when optimum intensity was obtained (3–5 min).
After rinse in PBS, slices were mounted on gelatin-coated slides and coverslipped with Mowiol mounting medium.
2.11. Image Acquisition and Data Processing
Fluorescence images were acquired by confocal microscopy (Zeiss LSM 710). In double-labeling experiments, images were sequentially acquired. All images were further processed in Adobe Photoshop 6.0, with only contrast and brightness adjustments. Photomicrographs labeling were taken on a Nikon Eclipse E600 light microscope using a DXM 1200 Camera equipped with ACT-1 software. The microscope was set at a specific illumination level, as was the camera exposure time. c-Fos-positive nuclei were counted using NIH ImageJ software. Labeling and counting were performed on every fourth coronal section throughout the rostro ¬¬–¬caudal axis from the brainstem to the hypothalamus.
2.12. Stereological Estimation of Cx43/Bassoon Apposition
Immunohistochemistry for 1st order stereologic quantifications: The PFA perfusion, cryoprotection and cryosection were performed as previously described [
3]. Five animals (15 weeks C57Bl6 mice) per condition at least (ad libitum food and water) were used to perform these quantifications. The DVC and the arcuate nucleus (ARC) in the hypothalamus were cut in 40 µm sections and immunohistochemistry was performed on free-floating sections [
3]; saturation solution for 1 h at room temperature, 2% BSA and 0.3% Triton X-100, then incubations using mouse primary antibody against Bassoon (#SAP7F407 Enzo Life Sciences, Farmingdale, NY, USA) at 1/3000 during 3 days at 4 °C, then secondary antibody (goat) directed against mouse IgG and coupled with Alexa 488 (1/400, overnight, 4 °C). Once again, the same saturation solution was used before the Cx43 rabbit primary antibody (#ACC-201 Alomone Labs 1/3000, Jerusalem, Israel) and goat anti-rabbit-IgG secondary antibody (1/400) on the same conditions. The long incubation times are necessary to obtain a strong and constant fluorescent signal throughout the entire section. The sections were serially mounted on slides, using Fluoroshield mounting medium (#104136, Abcam, Cambridge, UK) and then coverslipped.
Cavalieri method: the tissue was serially cut with uniform thickness of 40 µm. All the sections were taken into account. An entire DVC theoretically comprises 32 sections from rostral to caudal axis. NTS was present on all slices. In the hypothalamus, only the ARC including the median eminence (ME) is considered. It represents a mean number of 12 sections to match the entire nucleus. The total volumes of NTS and ARC were estimated by manually delimiting areas on section images. The reliability of these reference volume estimations was assessed by calculating a coefficient of error (CE) for each NTS and ARC of each mouse as described by Gundersen and Jensen [
18]. The estimation of the reference volume was considered accurate if the CE is lower than 7%.
Optical fractionator method: Depending on the average volume of the DVC, an x =2 00 µm and y = 300 µm grid was created to capture circa 150 voxels per animal. This grid was adapted for the ARC (200 µm × 200 µm). Confocal mosaic images were taken and this grid was randomly applied on each mosaic and every intersection within the considered structures was recorded as a voxel position. Voxels of 64 µm
3 (4 × 4 × 4 µm, z stacks=13) included a 27 µm
3 zone (3 × 3 × 3 µm) in the center of the voxels. Bassoon fluorescent signals were counted inside this zone. In order to restrict any overestimation, the points limiting the lower and right edges, the corners except the upper left one, the 2 first z-stacks and 2 last z-stacks (over 14) were excluded. The highlighted Bassoon fluorescent signal population was then split into 2: one with every Bassoon signal closes enough (less than 0.5 µm in every dimension of pure black) from a Cx43 signal and the other population with the remaining Bassoon signals. This counting includes the Cx43 signal out of the 27 µm
3 zone. We considered a point more than 0.1 µm-diameter as a significant fluorescent signal labeling of proteins. To avoid overestimation, two points should not have had any fluorescent connections in the 3 dimensions and must have had at least one z-stack of no-signal black between them. For each structure, a shrinkage coefficient was applied on voxels’ data according to the confocal measured thicknesses of sections. Note that for rostral levels of DVC, where the NTS is in touch with fourth ventricle, another coefficient reported the shrinkage differences between the voxels in the voxels’ thickness depending on their distances from ventricle. These coefficients were calculated after establishing the average thickness profiles of DVC or ARC sections for each animal and then averaging for all. The estimation of the total number of Bassoon signal close to Cx43 signal described previously was carried out according to West and colleagues [
19].
2.13. Ethidium Bromide Uptake and Quantification
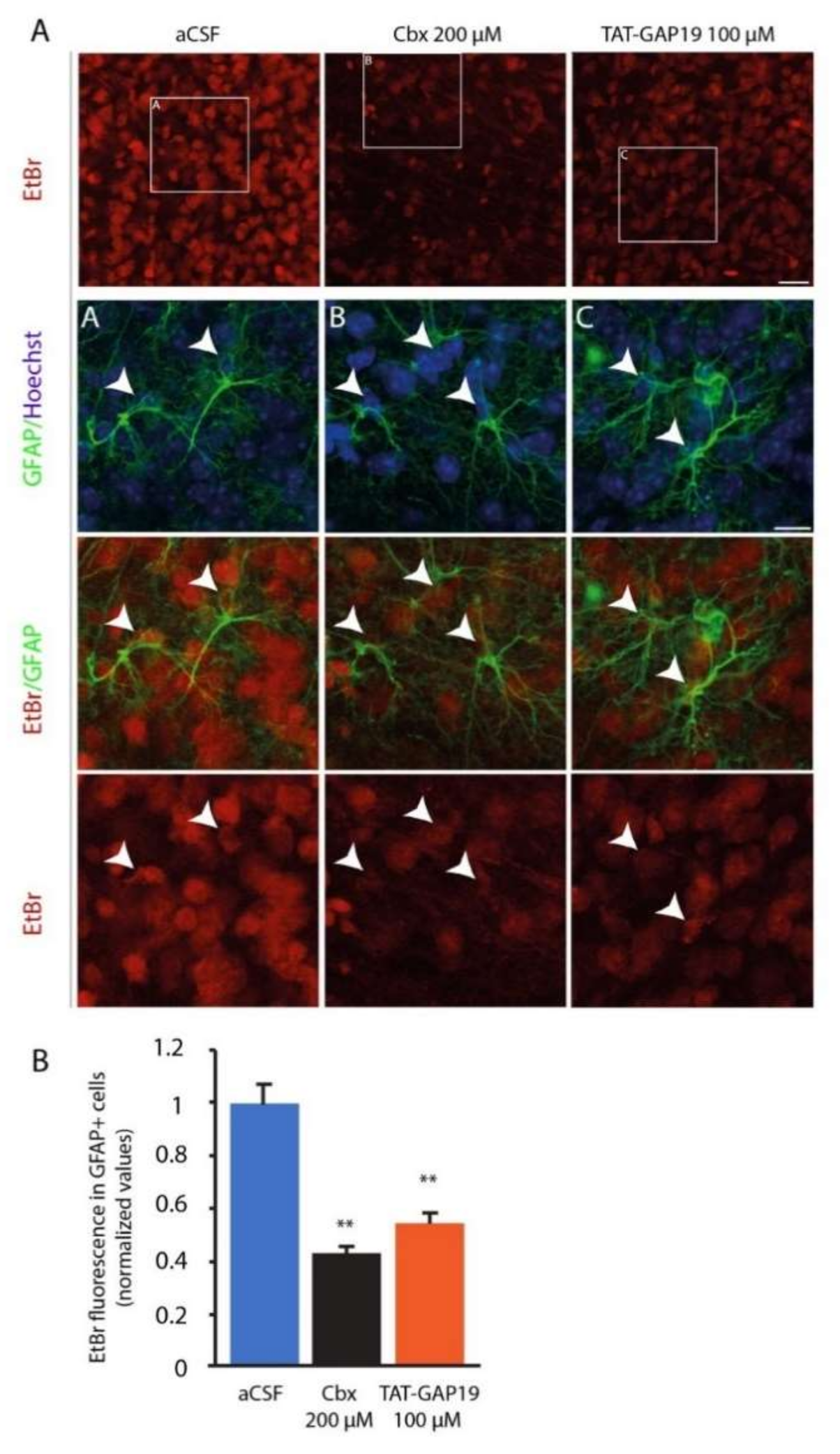
Ethidium Bromide Uptake was performed on acute hypothalamic slices prepared from 7-week-old mice fed ad libitum. Mice were euthanized by cervical dislocation followed by decapitation. Brains were removed and placed in ice-cold oxygenated (95% O2; 5% CO2) sucrose-enriched artificial cerebrospinal fluid containing (aCSF; in mM): 87 NaCl; 75 sucrose; 25 NaHCO3; 10 Glucose; 7 MgCl2, 2.5 KCl, 1 Na2HPO4; 0.5 CaCl2; 320–330 mOsm. The lateral borders of the brain were removed and coronal slices (300 µM thick) containing the mediobasal hypothalami were cut with a vibratome (VT2000S; Leica, Wetzlar, Germany). Slices were transferred to a constantly oxygenated (95% O2; 5% CO2) holding chamber containing sucrose-enriched aCSF for 30 min, then at least 30 min in the recording aCSF used for dye uptake assay containing: 119 NaCl; 26.2 NaHCO3; 11 Glucose; 2.5KCl; 2.5 CaCl2; 1.3 MgCl2; 1 Na2HPO4; 320–330 mOsm. Then, the slices were then randomly distributed in 10 mL submerged chambers and incubated for 15 min in recording aCSF, or in aCSF supplemented with carbenoxolone 200 µM or TAT-GAP19 100 µM before and during the application of EtBr (314 Da, 4 µM, 10 min). Slices were then washed for 20 min in aCSF and fixed for 1 h 30 in PBS containing 4% PFA before immunohistochemistry. Slices were treated with PBS containing 3% NGS, 1% Triton X-100 to block nonspecific binding sites. Slices were then incubated overnight with rabbit polyclonal anti-GFAP (1/1000, #Z03334, Dako, Santa Clara, CA, USA), washed in PBS and incubated for 1 h 30 with secondary antibody goat α-Rabbit-Alexa Fluor 488 (1/400, #A-11034, Invitrogen, Carlsbad, CA, USA). After rinse in PBS, slices were incubated 1 min with Hoechst 1/10000, rinsed, and mounted on gelatin-coated slides and coverslipped with Mowiol mounting medium. Images were acquired on a confocal microscope (ZEISS LSM 710) using the 488-nm (Alexa), 561-nm (BET) and 405 nm band. Dye uptake was evaluated in GFAP labelled cells and expressed as the difference between the fluorescence measured in GFAP+ astrocyte nuclei and the background fluorescence measured where no nuclei were detected. Values of fluorescence in different experimental conditions were normalized relative to the control level.
2.14. Statistical Analysis
All results are presented as mean ± SEM. Statistical analysis was performed using GraphPad Prism 6. Comparison between two groups was performed using unpaired two-tailed student’s t-test. One-way ANOVA was performed for comparisons between mice treated with different conditions followed by Dunnett’s post-test. A repeated-measure ANOVA was performed when needed.
4. Discussion
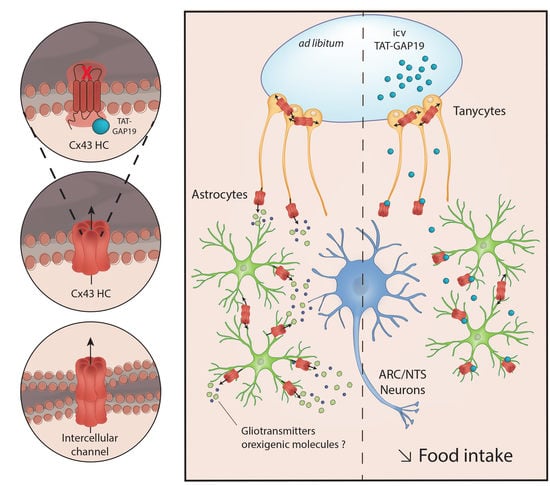
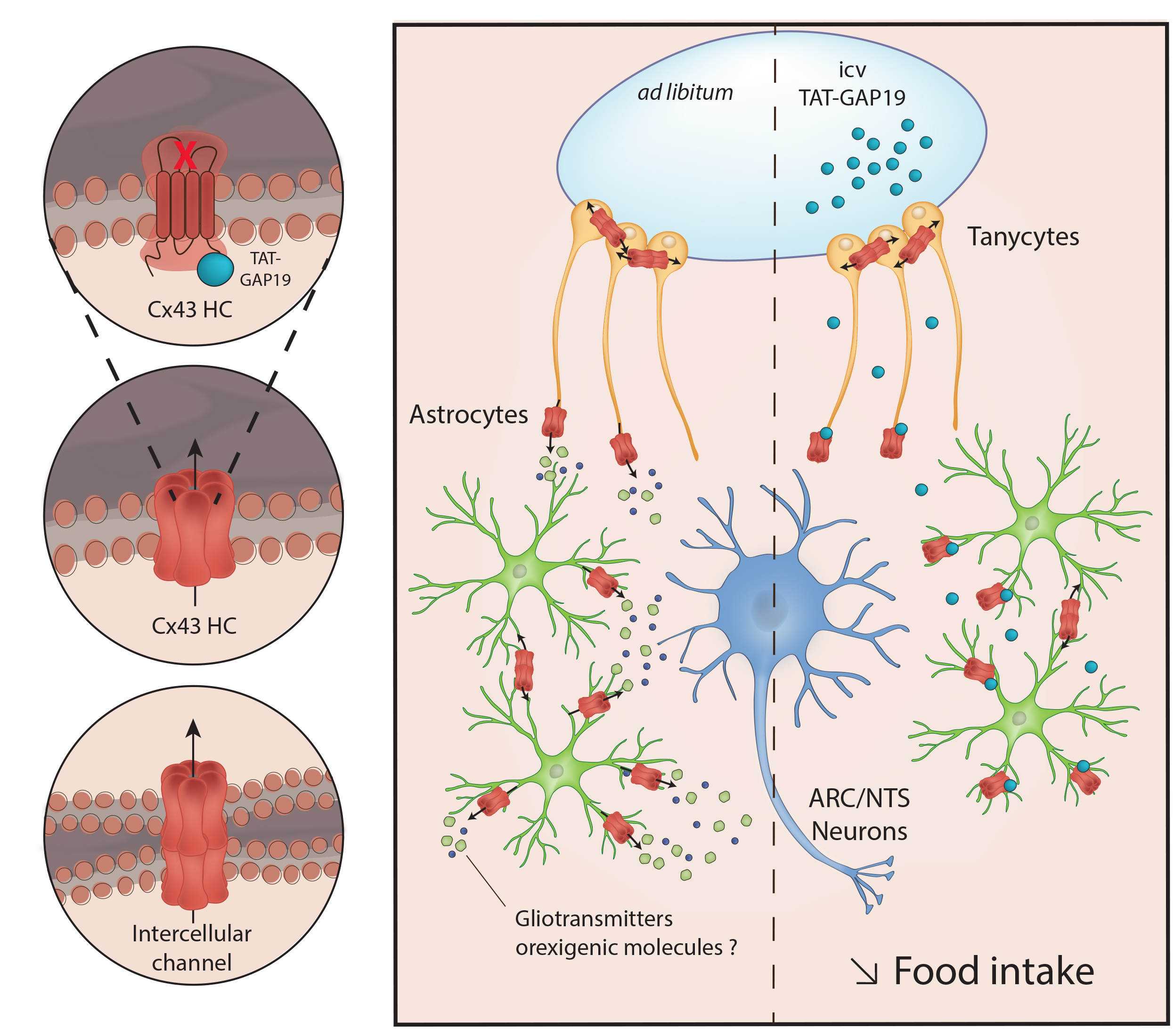
The present work shows that the two main structures involved in homeostatic regulation of energy balance are characterized by a notable expression of Cx43 associated with high glial density. Interestingly in the hypothalamus Cx43 expression is sensitive to nutritional status changes (fasting, obesity). In addition, a strong basal HCs activity in the ARC is suggested by the EtBr uptake assay. This HC activity is at least partially dependent on Cx43. Furthermore, the central administration of TAT-GAP19, a permeant peptide, that inhibits specifically the opening of Cx43 HCs, induces a decrease in food intake, without affecting locomotor activity. Finally, i.c.v. TAT-GAP19 administration induces cellular activation attested by c-Fos expression specifically in the hypothalamus and the DVC. Altogether, these results constitute the first demonstration that glial Cx43 HCs are involved in the regulation of food intake.
4.1. Cx43 Expression within the Hypothalamus and DVC and Modulation by Nutritional Status
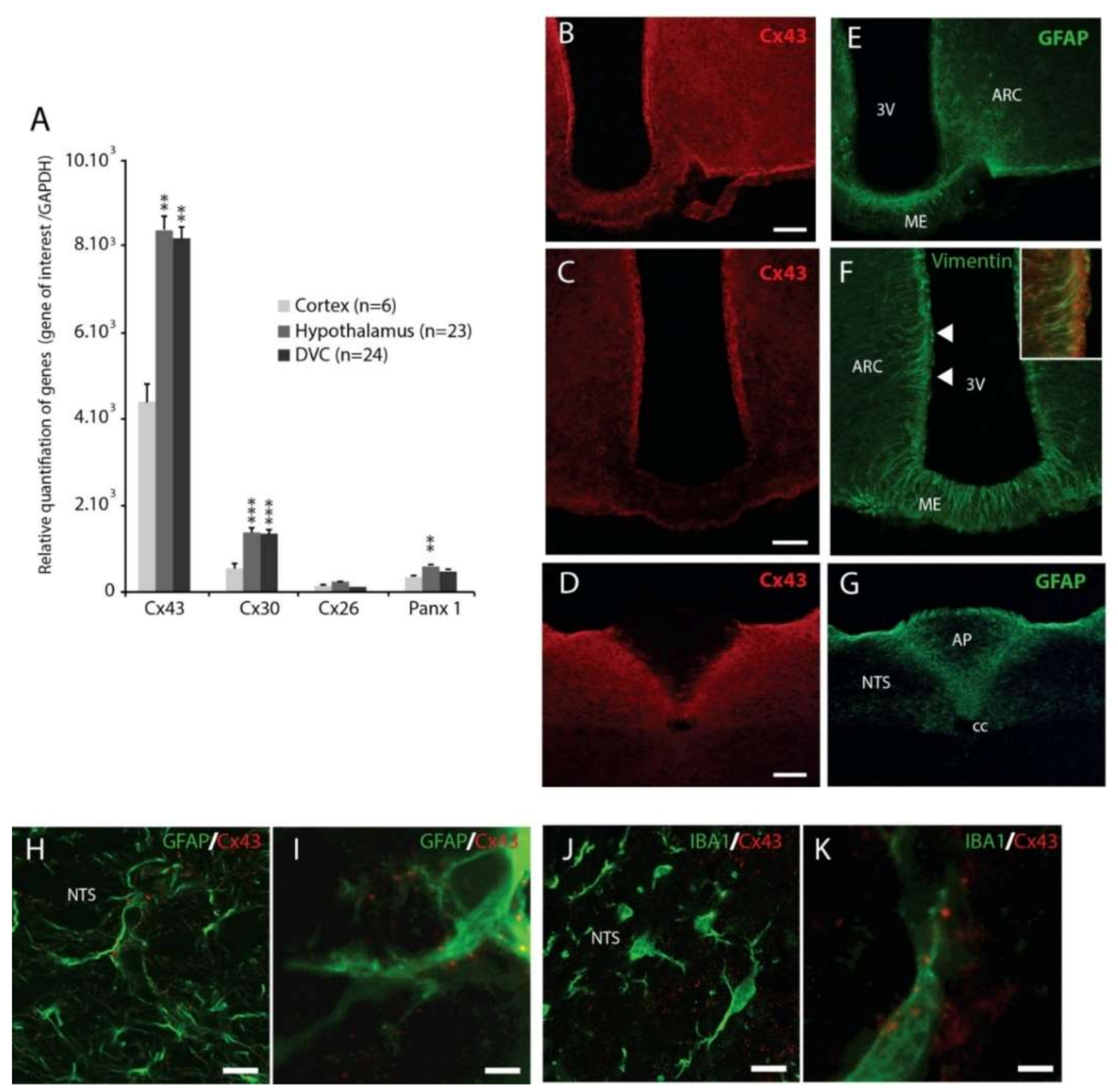
Cx43 is a ubiquitous protein found in almost all tissues of the body except red cells, sperm and skeletal muscle cells [
7]. Within the CNS, this protein is expressed by glia, mainly astrocytes and to a lesser extent microglia [
9]. Exceptionally, neuronal Cx43 expression has been reported in olfactory bulb neurons [
23] and spinal cord motor neurons [
24], where Cx43 is involved in the formation of electrical synapses. We first investigated Cx43 expression in the hypothalamus and DVC, two major centers liable for the regulation of the energy balance including homeostatic control of food intake. An analysis of transcripts encoding proteins known for their ability to form GJs and/or HCs in glial cells (Cx43, Cx30, Cx26 and Panx1) has shown that Cx43 is the most widely expressed in the hypothalamus and DVC. These structures are enriched in Cx43 mRNA when compared to the cortex, a structure taken here as a reference and reported to express high level of Cx43 in the adulthood [
25]. This strong Cx43 expression in the hypothalamus and DVC has been confirmed at the protein level by immunohistochemistry. These two structures are distinguished by a higher Cx43 immunoreactivity than surrounding structures. At the brainstem level, the Cx43 labelling clearly delimits the contours of the NTS. This high density Cx43 expression was found to a lesser extent at the hypothalamic level around the third ventricle and in the ARC. At higher magnifications, we confirmed the presence of Cx43 in membranes of GFAP + astrocytes. As previously described [
26,
27], tanycytes and more precisely β-tanycytes expressed Cx43. Interestingly, a population of tanycytes-like cells located within the DVC and originally described by Pecchi and colleagues [
20] also expressed Cx43. Finally, it should also be noted that some Cx43 labeling is also associated with IBA1
+ microglial cells. This cellular localization reported for the first time in the hypothalamus and DVC is consistent with the results of the literature obtained in other structures from the CNS [
9].
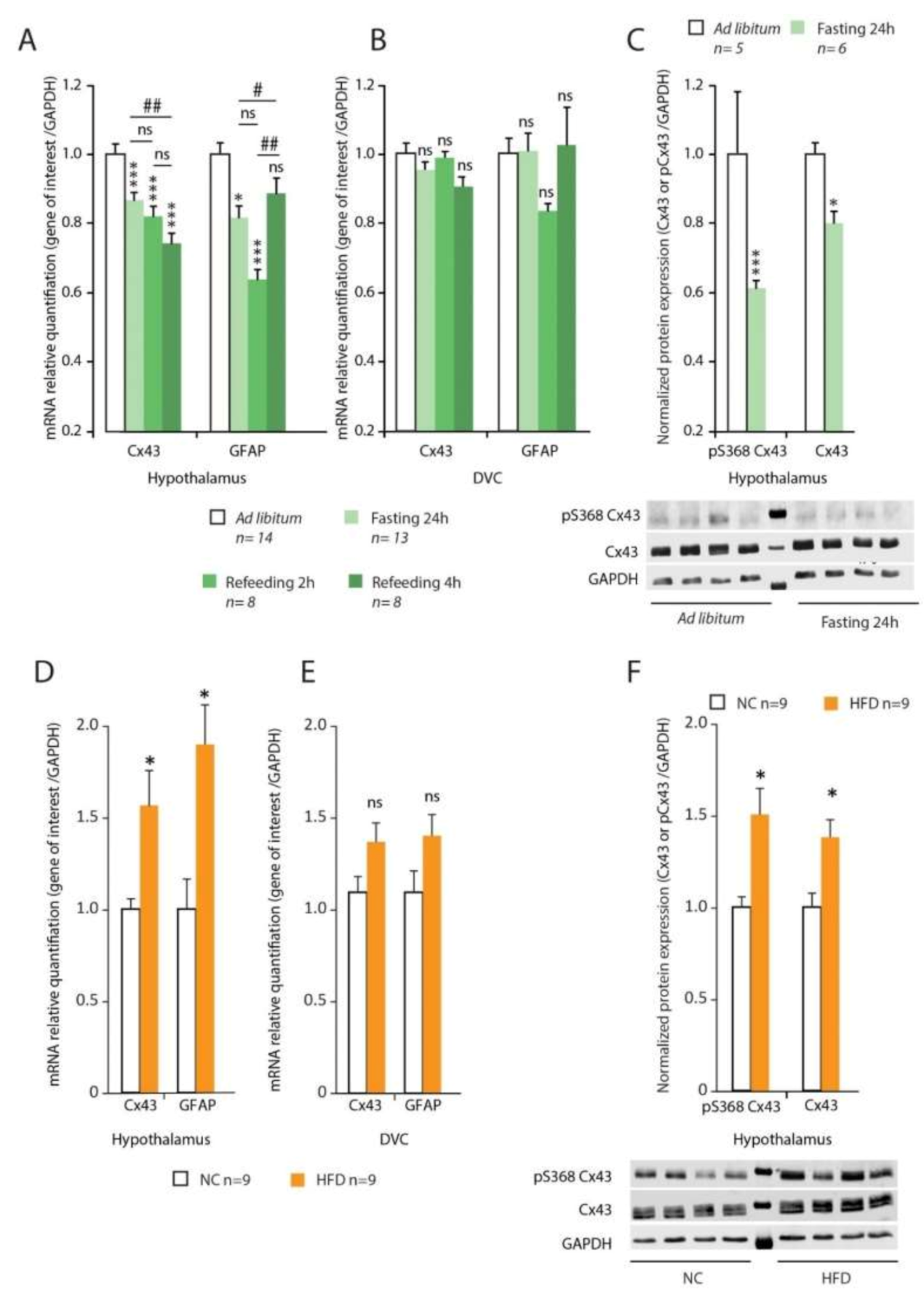
We then investigated whether Cx43 expression (mRNAs and protein) can be modified by drastic changes in energy balance. Control of Cx expression occurs mainly at the transcriptional level [
7] and the short Cx half-life, of the order of only a few hours (1.5 to 4 h for the Cx43, [
28], implies a permanent synthesis. We first chose to test a 24 h fast, known to cause a change in the energy substrates used. This change in metabolism is accompanied by a decrease in insulinemia and leptinemia and an increased secretion of ghrelin [
29]. Under these conditions, we observed a decrease in Cx43 expression in the hypothalamus. No change in Cx43 expression was observed in the DVC. The important contribution of the hypothalamus to counter-regulatory mechanisms observed during a fast [
30] may explain this result. Refeeding (2 and 4 h) did not restore the initial levels of Cx43. Secondly, we looked for variations in Cx43 expression in diet-induced obese (DIO) animals. We observed an upregulation of Cx43 expression in the hypothalamus of DIO animals, while again no modification was observed within the DVC. The turnover, internalization and degradation of Cxs including Cx43 are highly associated with phosphorylation/dephosphorylation events and can be triggered by a variety of stimuli (e.g., growth factors, extracellular matrix interactions, ischemia, inflammation, see [
28] for review). Cx43 phosphorylation by protein kinases especially at serine 368 has been associated to i/ GJ internalization, ii/ changes in GJ intercellular communication and iii/ reduced Cx43 half-life [
31]. Interestingly, we observed a marked decrease in phosphorylated Cx43 levels in the hypothalamus during fasting while during induced obesity, the level of phosphorylated Cx43 increased.
Using EtBr uptake as an index of HCs activity, we observed a high HCs permeability within the ARC of ad libitum-fed mice. This HCs activity relied significantly on Cx43 HCs, especially in GFAP+ astrocytes, since it was inhibited by TAT-GAP19, a mimetic peptide preventing interaction between the C-terminal part of Cx43 with the L2 loop and thus causing the specific closure of the Cx43 HCs [
12]. It is noteworthy that TAT-GAP19 has no influence on communication via the GJ made up of Cx43 [
12]. HCs are known to mediate the transfer of signaling molecules i.e., ATP and glutamate, between the cytoplasm and the extracellular space, constituting a non-vesicular route for gliotransmitters release (see [
32] for review). In agreement with this concept, blockade of Cx43 HCs with GAP26 resulted in reduced frequency of excitatory post-synaptic currents (fEPSPs) recorded in hippocampal CA1 neurons after Schaffer collateral stimulation via ATP signaling [
33]. Furthermore, it was demonstrated that glial Cx43 modulates glutamatergic synaptic activity of CA1 pyramidal cells via changes in synaptically released glutamate [
34].
In the hypothalamus and the DVC, Cx43 labelling was often found in close proximity, even juxtaposed, to bassoon positive elements. Bassoon is a very large scaffolding proteins of the cytomatrix assembled at the active zone where neurotransmitters are released [
21]. Our stereological study revealed that a high proportion of Cx43 signal (~80–85%) was found in the close environment of the synaptic elements (<1 µm). This result is quite different from a pioneer ultrastructural work by Sipe and Moore [
35] performed at the lateral hypothalamic area level reporting that about 90% of astrocytic GJ, identified on the basis of morphological criteria, was positioned within a distance of 15–20 µm from a synaptic terminal. On the other hand, our observation is to be compared to the work carried out at the barrel cortex where the average distance between an excitatory synapse and an astroglial Cx43 GJ, identified on electron-microscopic images, was about 0.7 µm [
36]. Nevertheless, contrary to cited works, our stereological analysis is not based on the morphological identification of GJ but on Cx43 immunofluorescence staining, which allows the detection of connexons involved in both GJ and HCs. Thus, we can assume that a proportion of Cx43 that we detected in the immediate vicinity of the synapse is not engaged in GJ but could constitute HCs. The high HCs activity we observed in hypothalamus slices support this assumption.
Altogether, these results suggest that, in structures involved in homeostatic regulation of food intake, Cx43 expression and Cx43 GJ and/or HCs gating can be regulated by hormonal and/or metabolic signals associated with nutritional status. In turn, these modulations could result in a disturbance of astrocytic and tanycytic GJ networks and/or the release of neuroactive compounds through Cx43 HCs.
4.2. The Inhibition of Cx43 HCs Diminishes Food Intake
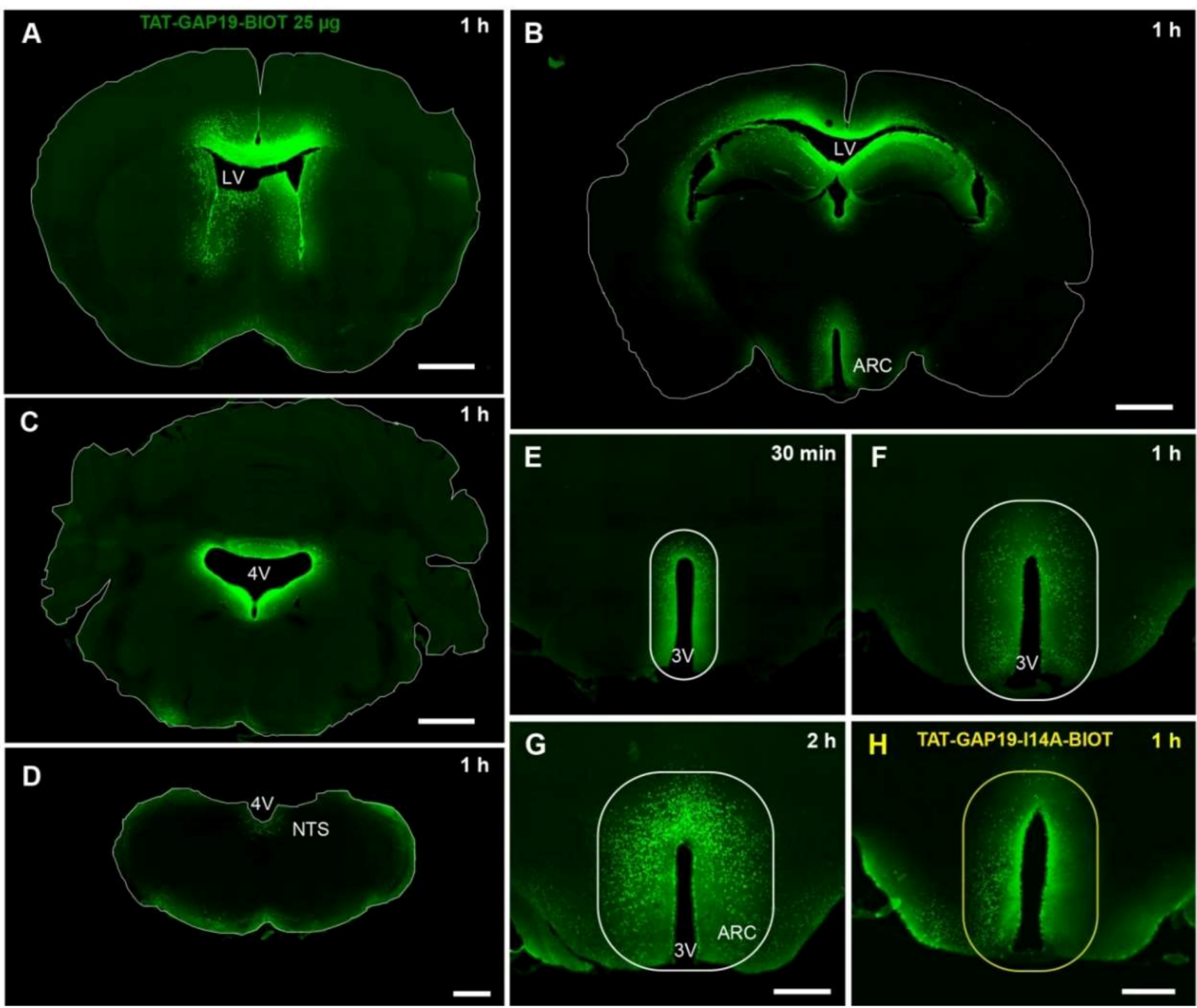
All of the results discussed above prompted us to test the effect of Cx43 HCs inhibition on food intake of ad libitum-fed mice. We postulated that Cx43 HCs activity could allow the release of neuroactive compounds and hence constitute a tonic modulation of neuronal networks dedicated to the control of food intake. To specifically target Cx43 HCs, we used GAP19 as previously done with EtBr uptake assays. Nonetheless, to facilitate GAP19 diffusion and cell penetration in vivo, we coupled GAP19 to a TAT sequence, derived from HIV. The TAT sequence facilitates cellular internalization of the peptide by a macropinocytosis mechanism [
37]. Moreover, we have chosen to administer this peptide by i.c.v. injection in lateral ventricles in order to reach both the hypothalamus and DVC, which border the third and fourth ventricles, respectively. This route of administration, which is not totally specific for the targeted structures, can potentially induce diffusion of the peptide in other brain structures and lead to unspecific behavioral effects. However, based on our previous results [
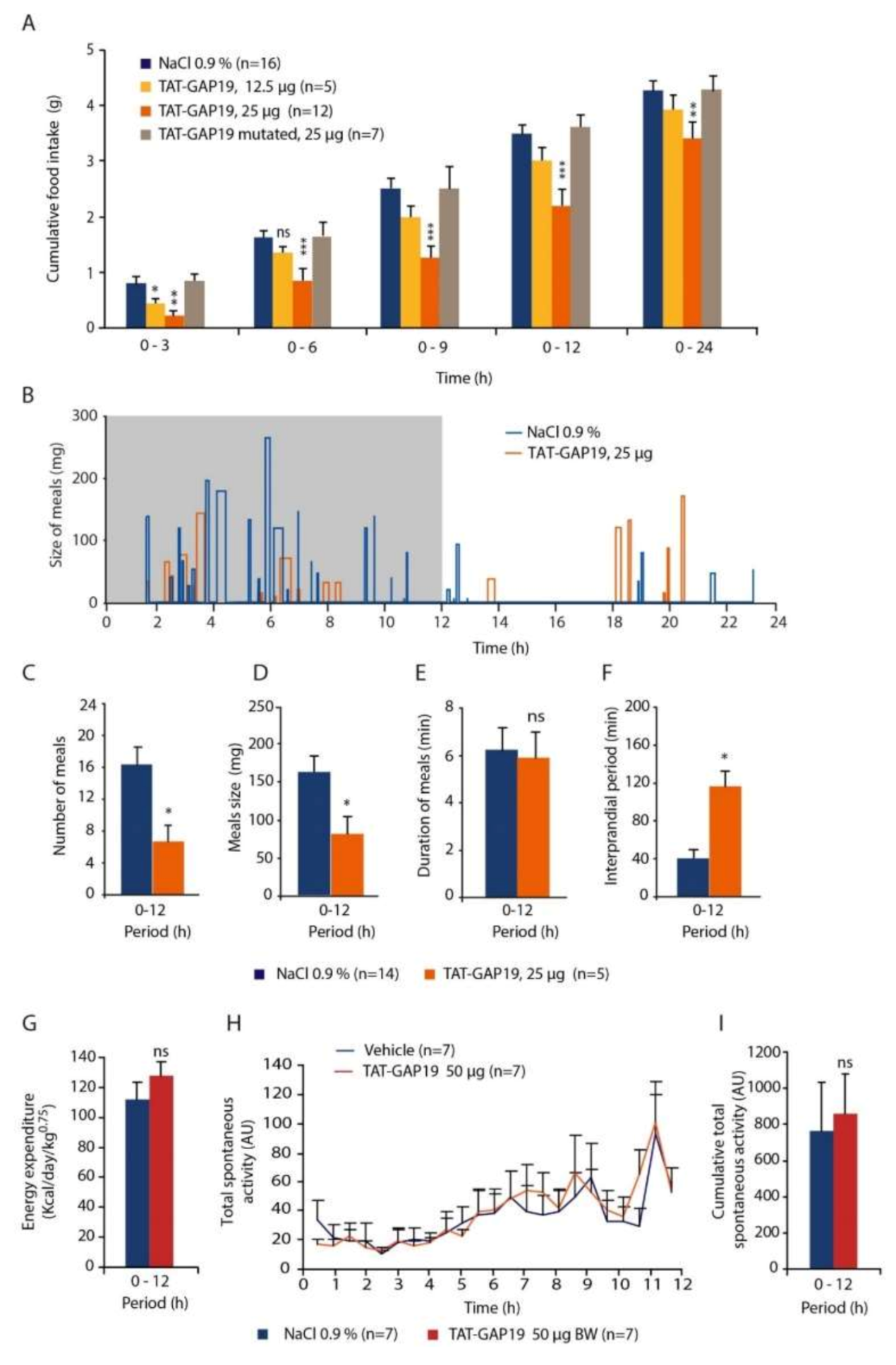
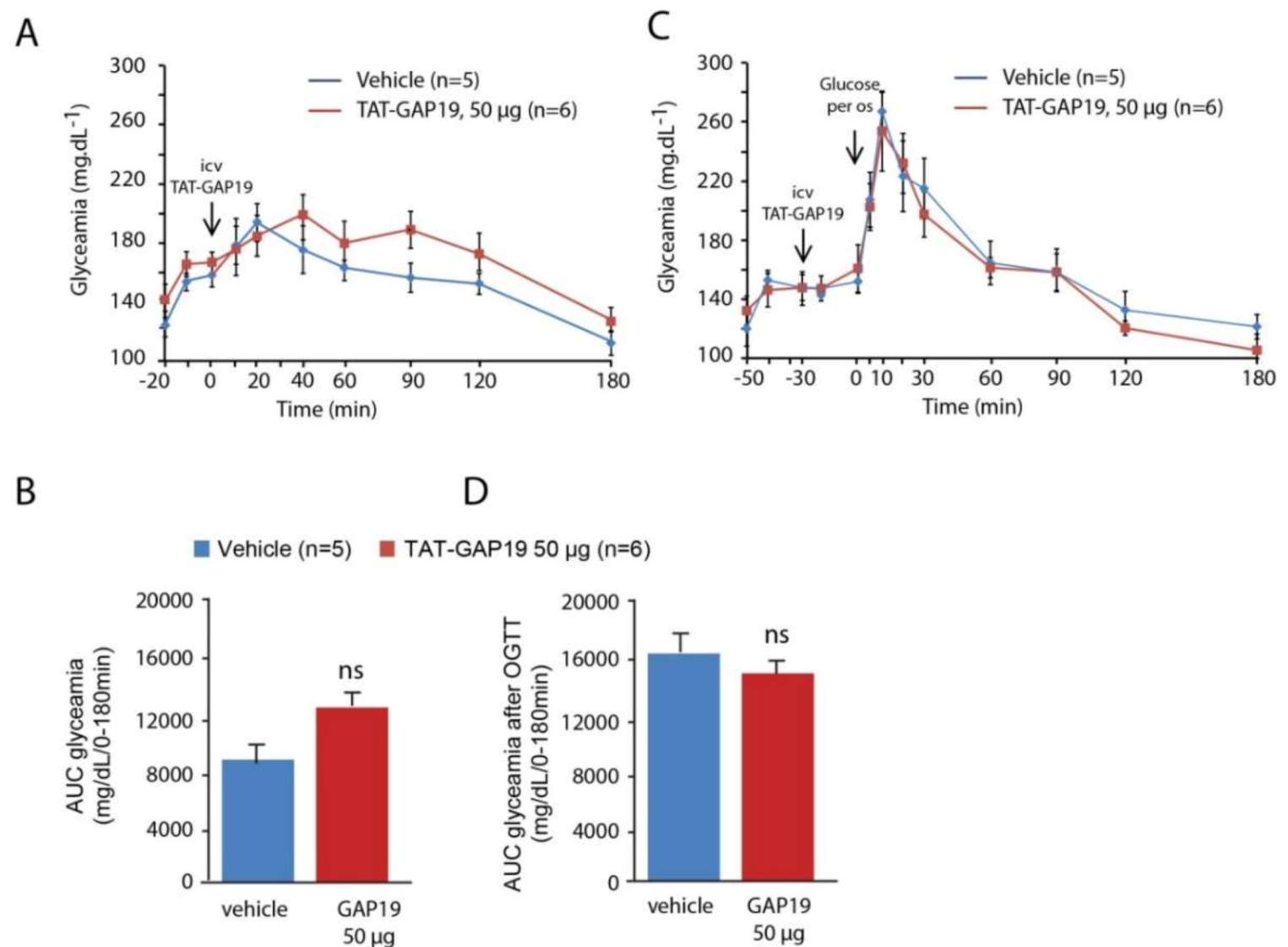
38], we postulated that this would not be the case because of the facilitated peptide diffusion from the ventricles to the ARC and NTS thanks to the proximity of the organs circumventricular, namely the median eminence and the area postrema. FITC labeling of biotinylated TAT-GAP19 or TAT-GAP19-I14A performed at different time points after their i.c.v. administration confirmed a privileged presence of the peptide in the ARC and NTS after its ventricular injection. Thus, we next evaluated the impact of i.c.v. TAT-GAP19 administration performed at the beginning of the mouse activity period. TAT-GAP19 rapidly (<1.5 h) diminished food intake, and this effect persisted throughout the dark phase. A meal microstructure analysis revealed that TAT-GAP19 causes both meal frequency and size decrease. While meal frequency reduction is evocative of a sickness behavior [
39], the decrease in meal size refereed to a more physiological process since endogenous substances regulating food intake, i.e., cholecystokinin, leptin, are known to modify the meal size. The possible nonspecific action of TAT-GAP19 inducing a broad distress resulting in anorexia seems unlikely since we did not observe signs of suffering. In accordance, i.c.v. TAT-GAP injection did not modify locomotor activity. In addition to food intake, we looked for a possible effect of TAT-GAP19 on carbohydrate regulation. The permeability of these HCs for the glucose suggests a role for Cx43 in central glucodetection, and it was previously reported that the inhibition of Cx43 synthesis, by interfering RNAs (siRNAs) injected into the rats hypothalamus parenchyma, decreases hypothalamic glucose sensitivity and glucose sensing-induced insulin secretion [
40]. Here, the TAT-GAP19 peptide did not alter glycaemia or insulinemic regulation following a glucose overload. The differences in the protocols used, i.e., a reduction of Cx43 expression by siRNAs and acute inhibition of Cx43 HCs activity could explain this apparent discrepancy. Finally, mapping of cellular activation using c-Fos immunolabelling [
41] was carried out on sections of the bulbar region and of the entire diencephalon. At a time where anorexia is ongoing, the i.c.v. TAT-GAP19 peptide injection induced cellular activation only in the hypothalamus and DVC. This activation profile is characteristic of cellular activation obtained after i.c.v. administration of appetite suppressant compounds [
3,
5,
17,
39,
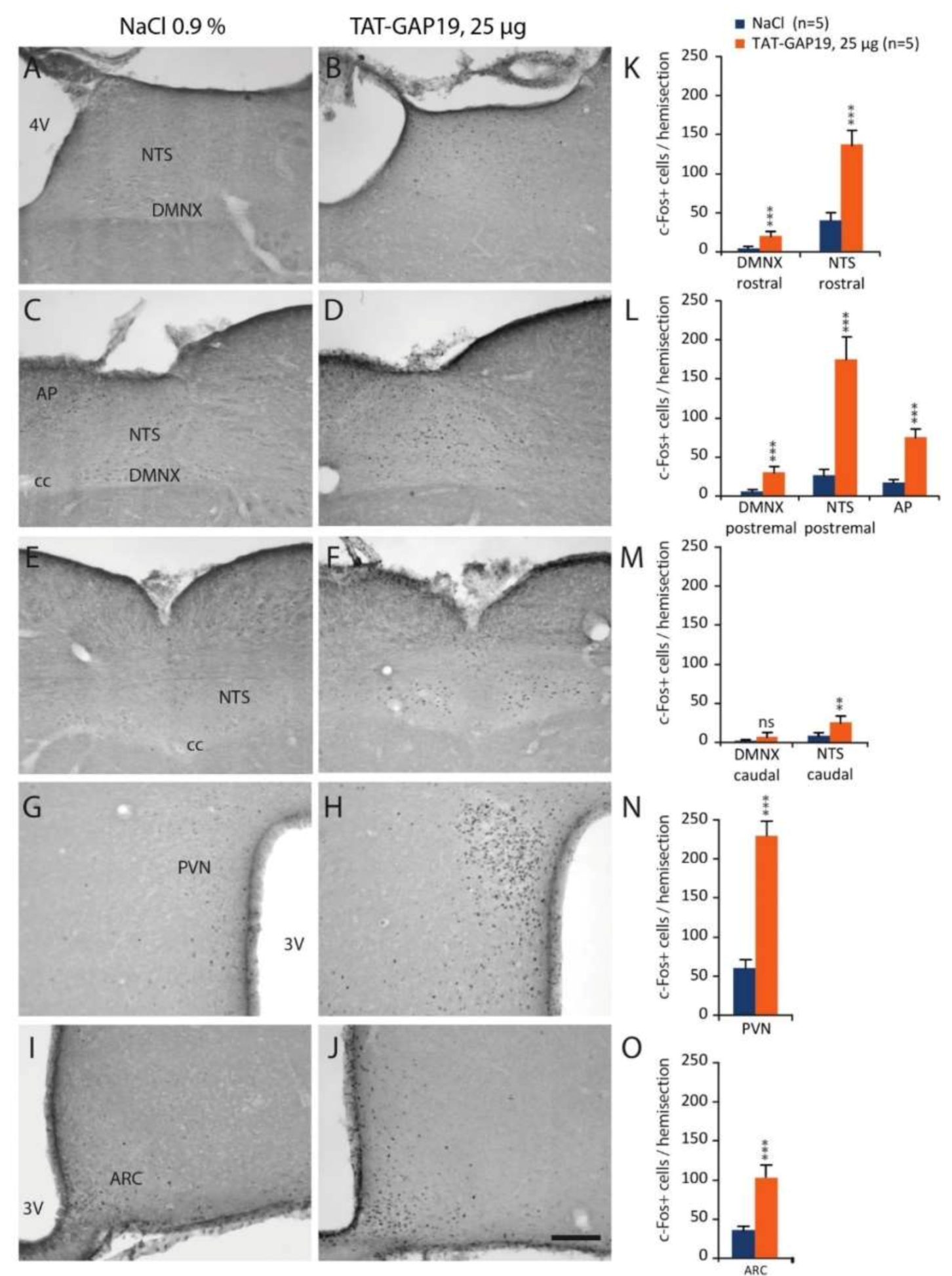
42]. This result shows that inhibition of Cx43 HCs within the hypothalamus and DVC modifies the activity of circuitries potentially involved in central appetite control, which could explain the diminution of meal number and size observed in the behavioral study.
4.3. Possible Mechanisms Underlying the Reduction of Food Intake Induced by Cx43 HCs Inhibition
The release of gliotransmitters and their involvement in brain functions are illustrated by numerous results obtained on different physiological and pathological models (see for reviews [
43,
44]). In this context, the release of neuroactive substances by Cx HCs has been largely proposed (see for review [
11]). Astrocytic release of ATP through the Cx26 HCs contributes to the control of breathing [
45]. Stehberg and colleagues [
13] have shown that the release of neuroactive substances via the astrocytic Cx43 HCs is necessary for long-term memorization of fear conditioning. Indeed, injection of TAT-Cx43L2 or GAP27 into the basolateral amygdala resulted in amnesia to fear conditioning training. Interestingly, co-administration of these blocking peptides with a gliotransmitter cocktail including glutamate, glutamine, lactate, D-serine, glycine, and ATP restored fear conditioning memory. Nevertheless, this study did not identify a specific gliotransmitter in mediating the effects of HCs on learning and memory. Similarly, the nature of neuroactive substance(s), released by hypothalamic and brainstem glial cells through Cx43 HCs, capable of modulating food intake remains to be identified. This issue appears as a puzzle since most of the compounds transported through Cx43 HCs have a profound impact on food intake. ATP, amino acids and their metabolites including glutamine glutamate, aspartate, glycine or D-serine can directly or indirectly (via modulation of neurotransmitters release) modify feeding behavior [
46]. Interestingly, most of them were known for their orexigenic action. For instance, glutamate was reported to stimulate feeding behavior by acting at the hypothalamic level [
47]. ATP appears also as a potential candidate. Its release through Cx43 HCs has been widely reported [
11], and this compound exerts orexigenic effects thanks to its action on P2X2 [
48] and P2Y [
49] receptors.
Another intriguing question that remains open is that of the glial cells identity at the origin of food intake regulation via HC Cx43. Presumably, GFAP + astrocytes are the primary cell type involved in such regulation given i/ their involvement in the tripartite synapse and astrocyte-neuron communications (see for review [
50]) and ii/ the strong Cx43 expression that we observed in GFAP+ cells within the hypothalamus and the DVC. However, tanycytes should not be overlooked. Indeed, we observed that Cx43 is expressed by a subpopulation of tanycytes both within the hypothalamus and DVC. Given their contribution to the regulation of food intake that has emerged in recent years [
1,
3,
51,
52], we can assume that tanycytes are susceptible to release, via Cx43 HCs, which are neuroactive substances acting on energy balance regulation. This is often the approach we used, i.e., icv injection of TAT-GAP19, as is it can clearly inhibit Cx43 HCs activity in the cells bordering both the third and fourth ventricles. Finally, it was proposed that microglia cells also contribute to the modulation of energy balance [
1]. Albeit, Cx43 seems to be scarcely expressed by hypothalamic and brainstem microglia, we cannot totally rule out the possibility that microglial Cx43 HCs inhibition underlines the effect of TAT-GAP19 on food intake. The deletion of the Cx43 gene in specific cell type could partly answer this question.
In summary, this study provides the first evidence indicating that inhibition of Cx43 HCs activity interferes with the control of food intake and suggests a possible tonic glial delivery of orexigenic molecules via Cx43 HCs. By reducing food intake, the inhibition of glial Cx43 could constitute a new therapeutic avenue against overweight, obesity and their comorbidities. More broadly, this work helps to support the hypothesis of the involvement of the glial compartment in regulating the energy balance.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}