Translocation of Drought-Responsive Proteins from the Chloroplasts

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
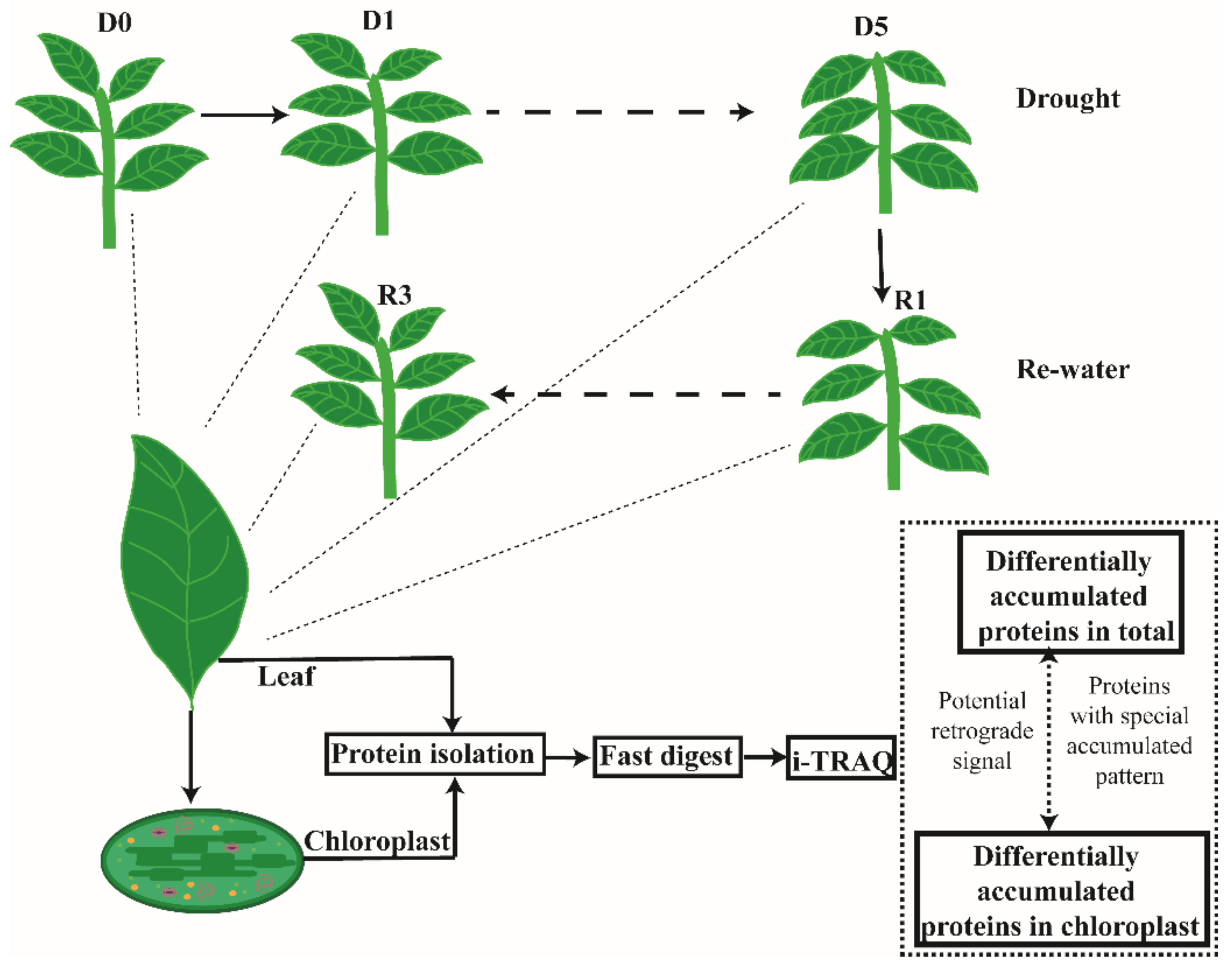
2.1. Plant Growth Conditions and Treatments
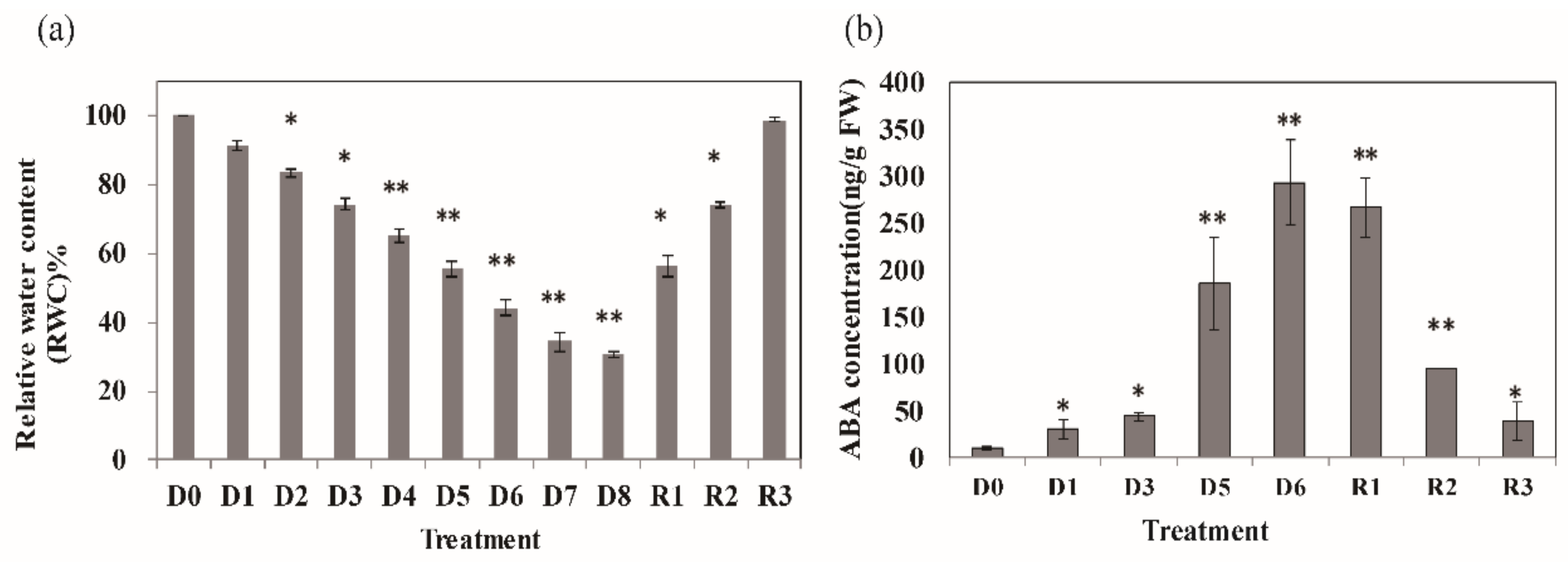
2.2. Quantification of Phytohormones
2.3. Preparation of Protein Samples
2.4. Western Blot Analysis
2.5. ITRAQ Labeling and LC-MS/MS Assay
2.6. Bioinformatic Analysis
2.7. Plasmid Construction and Transformation
3. Results
3.1. Physiological Changes in Nicotiana Benthamiana Plants under Drought Stress
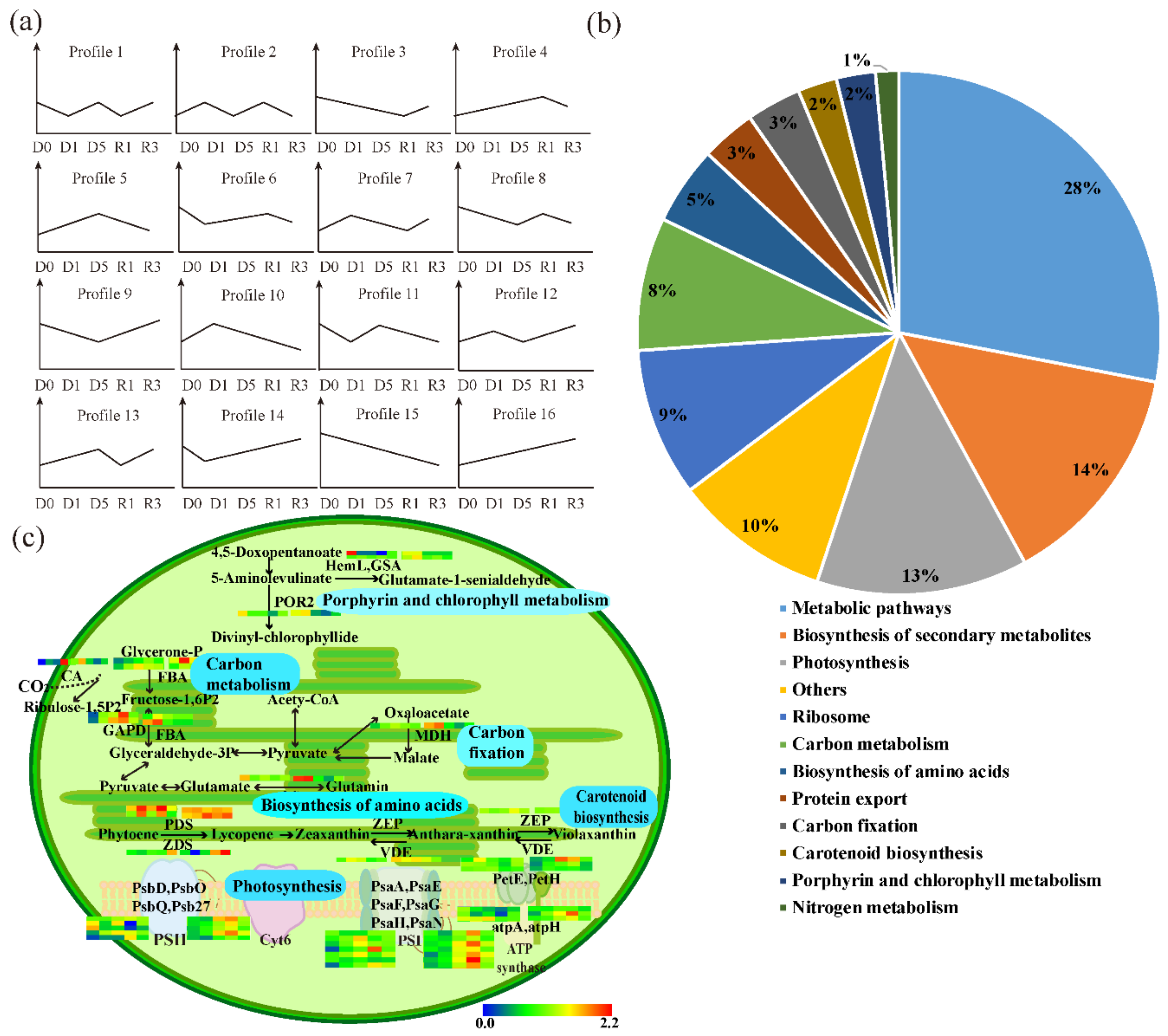
3.2. Differentially Accumulated Proteins (DAPs) in the Leaf and Chloroplasts under Drought Stress
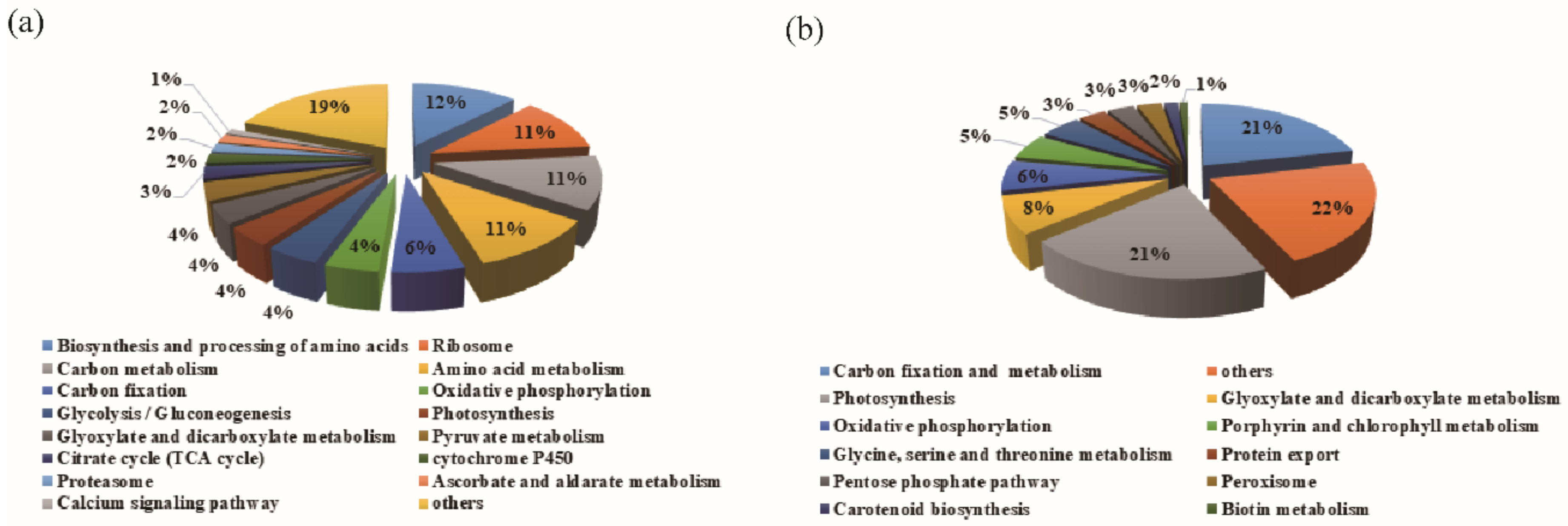
3.3. The Abundant Pathways Induced by Drought Stress in Leaves and Chloroplasts
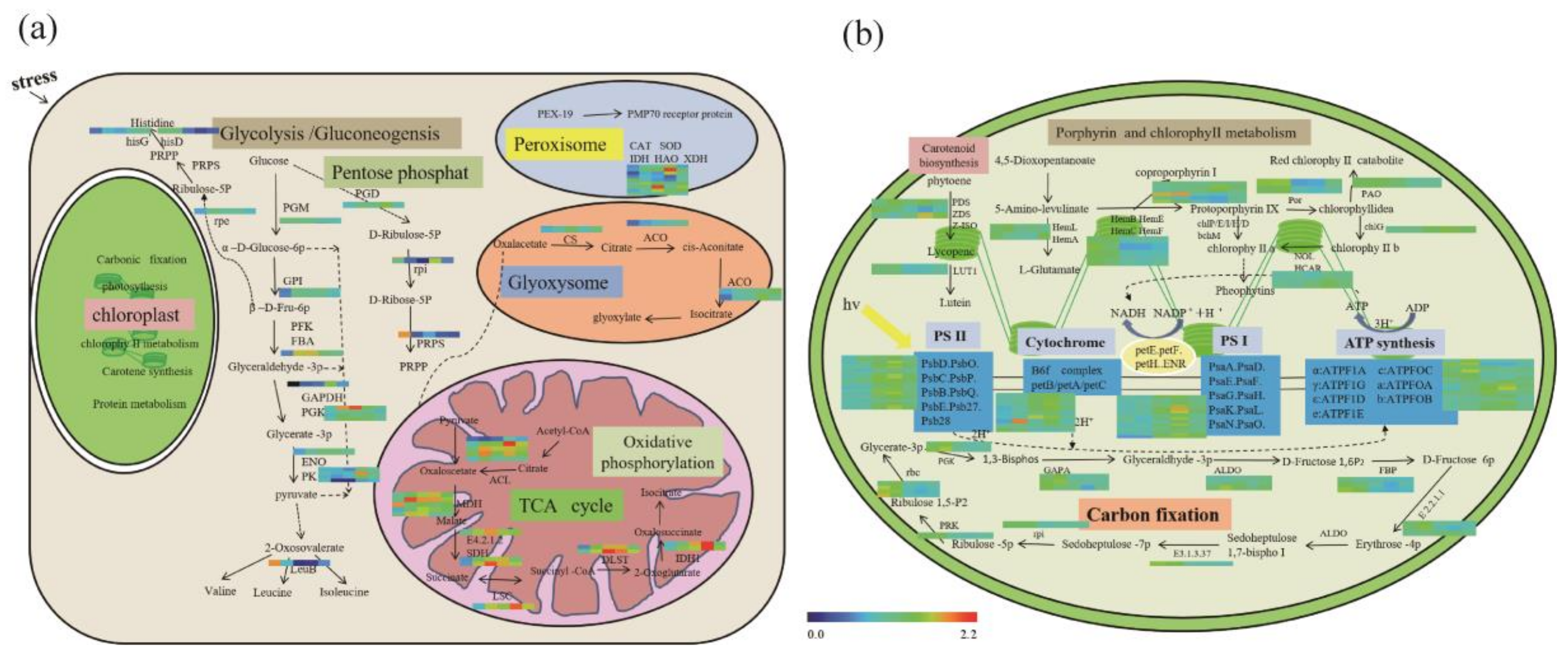
3.4. PSAPs Induced by Drought Stress in TPs and CPs
3.5. PSAPs Relocated under Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lobell, D.B.; Gourdji, S.M. The Influence of Climate Change on Global Crop Productivity1. Plant. Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosa, S.C.; Lupo, Y.; Moshelion, M. Quantitative and comparative analysis of whole-plant performance for functional physiological traits phenotyping: New tools to support pre-breeding and plant stress physiology studies. Plant. Sci. 2019, 282, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Song, T.; Wallrad, L.; Kudla, J.; Wang, X.; Zhang, W. Tissue-specific accumulation of pH-sensing phosphatidic acid determines plant stress tolerance. Nat. Plants 2019, 5, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrillo, E.; Herz, M.A.G.; Fuchs, A.; Reifer, D.; Fuller, J.; Yanovsky, M.J.; Simpson, C.; Brown, J.W.S.; Barta, A.; Kalyna, M.; et al. A chloroplast retrograde signal regulates nuclear alternative splicing. Sci. 2014, 344, 427–430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; De-Zhu, L.I. Advances in Phylogenomics Based on Complete Chloroplast Genomes. Plant. Div. Resour. 2011, 33, 365–375. [Google Scholar]
- Zoschke, R.; Bock, R. Chloroplast Translation: Structural and Functional Organization, Operational Control, and Regulation. Plant. Cell 2018, 30, 745–770. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Wang, J.-Z.; Dehesh, K. Review of stress specific organelles-to-nucleus metabolic signal molecules in plants. Encycl. Appl. Plant. Sci. 2013, 212, 102–107. [Google Scholar] [CrossRef]
- Sun, A.-Z.; Guo, F.-Q. Chloroplast Retrograde Regulation of Heat Stress Responses in Plants. Front. Plant. Sci. 2016, 7, 282. [Google Scholar] [CrossRef] [Green Version]
- Klein, M. The multi-protein family of Arabidopsis sulphotransferases and their relatives in other plant species. J. Exp. Bot. 2004, 55, 1809–1820. [Google Scholar] [CrossRef]
- Estavillo, G.M.; Crisp, P.A.; Pornsiriwong, W.; Wirtz, M.; Collinge, D.; Carrie, C.; Giraud, E.; Whelan, J.; David, P.; Javot, H.; et al. Evidence for a SAL1-PAP chloroplast retrograde pathway that functions in drought and high light signaling in Arabidopsis. Plant. Cell 2011, 23, 3992–4012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, W.; Feng, P.; Ma, J.; Zhang, L. Metabolites and chloroplast retrograde signaling. Curr. Opin. Plant. Boil. 2015, 25, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, S.; Parihar, P.; Singh, V.P.; Prasad, S.M. Retrograde signaling between plastid and nucleus: A review. J. Plant. Physiol. 2015, 181, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhao, L.; Zhou, J.; Zhong, X.; Gu, F.; Liu, Q.; Li, H.; Guo, R. iTRAQ protein profile analysis of young and old leaves of cotton (Gossypium hirsutum L.) differing in response to alkali stress. Plant. Physiol. Biochem. 2019, 141, 370–379. [Google Scholar] [CrossRef]
- Lee, K.H.; Park, J.; Williams, D.S.; Xiong, Y.; Hwang, I.; Kang, B.H. Defective chloroplast development inhibits maintenance of normal levels of abscisic acid in a mutant of the Arabidopsis RH3 DEAD-box protein during early post-germination growth. Plant. J. 2013, 73, 720–732. [Google Scholar] [CrossRef]
- Nawaz, G.; Kang, H. Chloroplast- or Mitochondria-Targeted DEAD-Box RNA Helicases Play Essential Roles in Organellar RNA Metabolism and Abiotic Stress Responses. Front. Plant. Sci. 2017, 8, 871. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, F.; Hong, Y.; Huang, J.; Shi, H.; Zhu, J.-K. Two Chloroplast Proteins Suppress Drought Resistance by Affecting ROS Production in Guard Cells1. Plant. Physiol. 2016, 172, 2491–2503. [Google Scholar] [CrossRef]
- Hu, H.H.; Aurélien, B.-D.; Maria, I.-N.; Maik, B.; Xue, S.W.; Amber, R.; Jan, G.; Josef, M.K.; Julian, I.S. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat. Cell Biol. 2010, 1212, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Liao, C.; Zhao, S.; Wang, C.; Guo, Y. The Glycosyltransferase QUA1 Regulates Chloroplast-Associated Calcium Signaling During Salt and Drought Stress in Arabidopsis. Plant. Cell Physiol. 2017, 58, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Blumwald, E. Stress-Induced Chloroplast Degradation in Arabidopsis Is Regulated via a Process Independent of Autophagy and Senescence-Associated Vacuoles[W]. Plant. Cell 2014, 26, 4875–4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Wang, S.; Tian, L.; Wu, L.; Li, M.; Zhang, J.; Li, P.; Zhang, W.; Chen, Y. Comparative proteomic analysis of the maize responses to early leaf senescence induced by preventing pollination. J. Proteom. 2018, 177, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Zhang, J.; Liu, D.; Yin, C.; Wang, F.; Chen, P.; Chen, H.; Ma, J.; Zhang, B.; Xu, J.; et al. iTRAQ-based analysis of developmental dynamics in the soybean leaf proteome reveals pathways associated with leaf photosynthetic rate. Mol. Genet. Genom. 2016, 291, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Xu, F.; Sun, J.; Peng, Z.; He, S.; Pan, Z.; Du, X. iTRAQ-Based Comparative Proteomic Analysis of Seedling Leaves of Two Upland Cotton Genotypes Differing in Salt Tolerance. Front. Plant. Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Y.; Cao, Y.; Lei, Y.; Jiang, H. Quantitative Proteomic Analysis Reveals Populus cathayana Females Are More Sensitive and Respond More Sophisticatedly to Iron Deficiency than Males. J. Proteome Res. 2016, 15, 840–850. [Google Scholar] [CrossRef]
- Lei, R.; Du, Z.; Kong, J.; Li, G.; He, Y.; Qiu, Y.; Yan, J.; Zhu, S. Blue Native/SDS-PAGE and iTRAQ-Based Chloroplasts Proteomics Analysis of Nicotiana tabacum Leaves Infected with M Strain of Cucumber Mosaic Virus Reveals Several Proteins Involved in Chlorosis Symptoms. Proteomics 2018, 18. [Google Scholar] [CrossRef]
- Kosmala, A.; Perlikowski, D.; Pawłowicz, I.; Rapacz, M. Changes in the chloroplast proteome following water deficit and subsequent watering in a high- and a low-drought-tolerant genotype of Festuca arundinacea. J. Exp. Bot. 2012, 63, 6161–6172. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple exposures to drought ’train’ transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, X.; Zhou, F.; Zhang, Z. Accessing the agronomic and photosynthesis-related traits of high-yielding winter wheat mutants induced by ultra-high pressure. Field Crop. Res. 2017, 213, 165–173. [Google Scholar] [CrossRef]
- Huang, Q.C.; Wei, Y.h.; Wei, F.l. The Comparative Analysis of Leaf Color and Carotenoids Content of the Common Apocynaceae Plants. J. Anhui Agric. Sci. 2011, 39, 15203–15262. [Google Scholar]
- Cai, B.-D.; Ye, E.-C.; Yuan, B.-F.; Feng, Y.-Q. Sequential solvent induced phase transition extraction for profiling of endogenous phytohormones in plants by liquid chromatography-mass spectrometry. J. Chromatogr. B 2015, 1004, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Radhakrishnan, R.; Kang, S.-M.; Kim, J.-H.; Lee, I.-Y.; Moon, B.-K.; Yoon, B.-W.; Lee, I.-J. Phytotoxic mechanisms of bur cucumber seed extracts on lettuce with special reference to analysis of chloroplast proteins, phytohormones, and nutritional elements. Ecotoxicol. Env. Saf. 2015, 122, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liang, W.; Xing, J.; Tan, F.; Chen, Y.; Huang, L.; Cheng, C.-L.; Chen, W. Dynamics of Chloroplast Proteome in Salt-Stressed MangroveKandelia candel(L.) Druce. J. Proteome Res. 2013, 12, 5124–5136. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Li, B.; Xing, S.; Pan, T. Reconfigurable microfluidic dilution for high-throughput quantitative assays. Lab. A Chip 2015, 15, 2670–2679. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.L. Multiplexed Protein Quantitation in Saccharomyces cerevisiae Using Amine-reactive Isobaric Tagging Reagents. Mol. Cell. Proteom. 2004, 3, 1154–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Yang, D.-H.; Yao, H.; Bai, G.; Zhang, Y.-H.; Xiao, B.-G. iTRAQ-based quantitative proteomic analysis reveals proteomic changes in leaves of cultivated tobacco (Nicotiana tabacum) in response to drought stress. Biochem. Biophys. Res. Commun. 2016, 469, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; López, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Morinaga, K.; Kohno, H.; Maruyama, Y.; Ninomiya, T.; Ishitobi, Y.; Tanaka, Y.; Tsuru, J.; Hanada, H.; Yoshikawa, T.; et al. An uncommon case of random fire-setting behavior associated with Todd paralysis: A case report. BMC Psychiatry 2012, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Yu, A.; Li, P.; Yang, H.; Liu, G.; Liu, L. Sequence analysis of the Hsp70 family in moss and evaluation of their functions in abiotic stress responses. Sci. Rep. 2016, 6, 33650. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Mao, J.; Liu, F.; Zeng, F. Expression of a nematode symbiotic bacterium-derived protease inhibitor protein in tobacco enhanced tolerance against Myzus persicae. Plant. Cell Rep. 2012, 31, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Ghassemian, M.; Anderson, J.C.; Peck, S.C.; Hu, H.; Schroeder, J.I. Carbonic anhydrases, EPF2 and a novel protease mediate CO2 control of stomatal development. Nature 2014, 513, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, T.; Hiwatashi, Y.; Sakakibara, K.; Kato, M.; Hasebe, M. Tagged Mutagenesis and Gene-trap in the Moss, Physcomitrella patens by Shuttle Mutagenesis. Curr. Neuropharmacol. 2000, 7, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.; Sangireddy, S.; Okekeogbu, I.; Zhou, S.; Yu, C.-L.; Hui, D.; Howe, K.J.; Fish, T.; Thannhauser, T.W. Drought-Induced Leaf Proteome Changes in Switchgrass Seedlings. Int. J. Mol. Sci. 2016, 17, 1251. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Yang, Z.; Wang, J.; Wang, P.; Ma, X.; Zhou, M.; Li, J.; Gang, N.; Feng, G.; Zhao, J.; et al. Comparative proteomic analyses reveal the proteome response to short-term drought in Italian ryegrass (Lolium multiflorum). PLoS ONE 2017, 12, e0184289. [Google Scholar] [CrossRef]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.-S.; Penfold, C.A.; Jenkins, D.; et al. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant. Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Adam, D.L.; Franklin, T.-R.; Samuel, A.H.; Barton, M.K. The Arabidopsis transcription factor ABIG1 relays ABA signaled growth inhibition and drought induced senescence. Elife 2016, 5, e13768. [Google Scholar] [CrossRef]
- Noor, E.; Bar-Even, A.; Flamholz, A.; Reznik, E.; Liebermeister, W.; Milo, R. Pathway Thermodynamics Highlights Kinetic Obstacles in Central Metabolism. PLoS Comput. Boil. 2014, 10, e1003483. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.N.; Ignatova, L.K.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic anhydrases in photosynthetic cells of higher plants. Biochemistry 2015, 80, 674–687. [Google Scholar] [CrossRef]
- Zhao, C.; Haigh, A.M.; Holford, P.; Chen, Z.-H. Roles of Chloroplast Retrograde Signals and Ion Transport in Plant Drought Tolerance. Int. J. Mol. Sci. 2018, 19, 963. [Google Scholar] [CrossRef] [Green Version]
- Uberegui, E.; Hall, M.; Lorenzo, Ó.; Schröder, W.P.; Balsera, M. An Arabidopsis soluble chloroplast proteomic analysis reveals the participation of the Executer pathway in response to increased light conditions. J. Exp. Bot. 2015, 66, 2067–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwang-Chul, K.; Verma, D.; Jin, S.; Singh, N.D.; Daniell, H. Release of Proteins from Intact Chloroplasts Induced by Reactive Oxygen Species during Biotic and Abiotic Stress. PLoS ONE 2013, 8, e67106. [Google Scholar]
- McKenna, R.; Frost, S.C. Overview of the carbonic anhydrase family. Subcell Biochem 2014, 75, 3–5. [Google Scholar] [PubMed]
- Hu, H.; Rappel, W.-J.; Occhipinti, R.; Ries, A.; Böhmer, M.; You, L.; Xiao, C.; Engineer, C.B.; Boron, W.F.; Schroeder, J.I. Distinct Cellular Locations of Carbonic Anhydrases Mediate Carbon Dioxide Control of Stomatal Movements. Plant. Physiol. 2015, 169, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, Z.; Biener, G.; Xiong, E.; Malik, S.; Eaton, N.; Zhao, C.Z.; Raicu, V.; Kong, H.; Zhao, D. Carbonic Anhydrases Function in Anther Cell Differentiation Downstream of the Receptor-Like Kinase EMS1. Plant. Cell 2017, 29, 1335–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Wu, H.; Wu, J.; Fan, X.; Li, X.; Lin, Y. Absence of OsbetaCA1 causes a CO2 deficit and affects leaf photosynthesis and the stomatal response to CO2 in rice. Plant. J. 2017, 90, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Studer, A.J.; Gandin, A.; Kolbe, A.R.; Wang, L.; Cousins, A.B.; Brutnell, T.P. A Limited Role for Carbonic Anhydrase in C4 Photosynthesis as Revealed by a ca1ca2 Double Mutant in Maize. Plant. Physiol. 2014, 165, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Slaymaker, D.H.; Navarre, D.A.; Clark, D.; Del Pozo, O.; Martin, G.B.; Klessig, D.F. The tobacco salicylic acid-binding protein 3 (SABP3) is the chloroplast carbonic anhydrase, which exhibits antioxidant activity and plays a role in the hypersensitive defense response. Proc. Natl. Acad. Sci. 2002, 99, 11640–11645. [Google Scholar] [CrossRef] [Green Version]
- Medina-Puche, L.; Castelló, M.J.; Canet, J.V.; Lamilla, L.; Colombo, M.L.; Tornero, P. β-carbonic anhydrases play a role in salicylic acid perception in Arabidopsis. PLoS ONE 2017, 12, e0181820. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
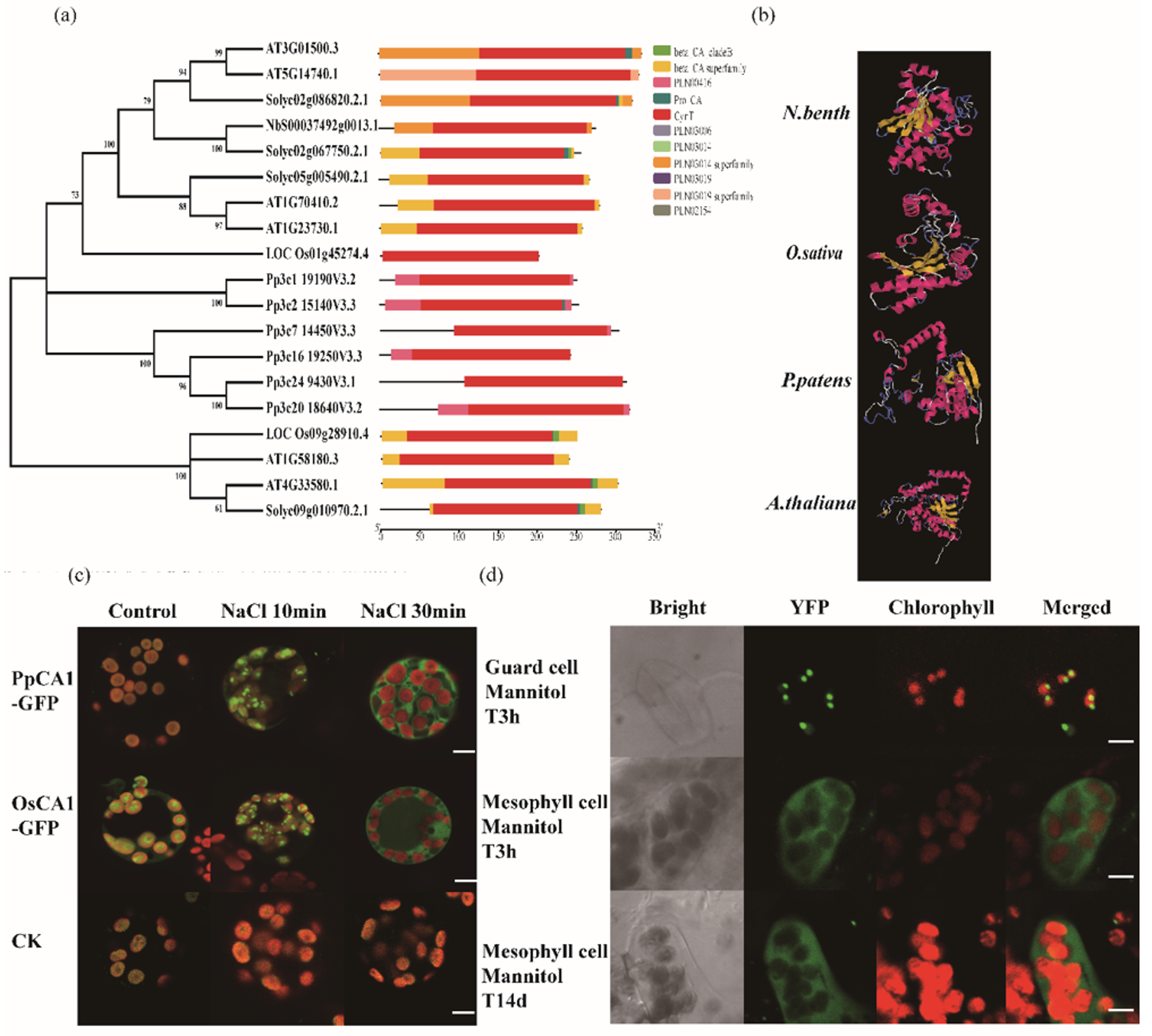
| Protein ID | Species | E-Value | Identity |
|---|---|---|---|
| Pp3c1_19190 | Physcomitrella patens | 1 × 1080 | 0.49 |
| Pp3c16_19250 | Physcomitrella patens | 2 × 1072 | 0.5 |
| Pp3c2_15140 | Physcomitrella patens | 1 × 1070 | 0.41 |
| Pp3c24_9430 | Physcomitrella patens | 13× 1064 | 0.46 |
| Pp3c20_18640 | Physcomitrella patens | 5× 1064 | 0.45 |
| Pp3c7_14450 | Physcomitrella patens | 3 × 1061 | 0.42 |
| LOC_Os01g45274.4 | Oryza sativa | 3 × 1081 | 0.57 |
| LOC_Os09g28910.4 | Oryza sativa | 7× 1062 | 0.45 |
| AT3G01500.3 | Arabidopsis thaliana | 1 × 10115 | 0.64 |
| AT5G14740.1 | Arabidopsis thaliana | 1.7 × 10113 | 0.637 |
| AT1G70410.2 | Arabidopsis thaliana | 13.5× 10101 | 0.563 |
| AT1G23730.1 | Arabidopsis thaliana | 6.3 × 1097 | 0.554 |
| AT1G58180.3 | Arabidopsis thaliana | 5.3 × 10 43 | 0.367 |
| AT4G33580.1 | Arabidopsis thaliana | 4.8 × 1054 | 0.403 |
| Solyc02g086820.2.1 | Solanum lycopersicum | 2.8 × 10130 | 0.708 |
| Solyc02g067750.2.1 | Solanum lycopersicum | 3.2× 10105 | 0.81 |
| Solyc05g005490.2.1 | Solanum lycopersicum | 1.1 × 10112 | 0.625 |
| Solyc09g010970.2.1 | Solanum lycopersicum | 1.9 × 1058 | 0.497 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Liu, H.; Yang, H.; Pu, X.; Li, C.; Huo, H.; Chu, Z.; Chang, Y.; Lin, Y.; Liu, L. Translocation of Drought-Responsive Proteins from the Chloroplasts. Cells 2020, 9, 259. https://doi.org/10.3390/cells9010259
Li P, Liu H, Yang H, Pu X, Li C, Huo H, Chu Z, Chang Y, Lin Y, Liu L. Translocation of Drought-Responsive Proteins from the Chloroplasts. Cells. 2020; 9(1):259. https://doi.org/10.3390/cells9010259
Chicago/Turabian StyleLi, Ping, Haoju Liu, Hong Yang, Xiaojun Pu, Chuanhong Li, Heqiang Huo, Zhaohui Chu, Yuxiao Chang, Yongjun Lin, and Li Liu. 2020. "Translocation of Drought-Responsive Proteins from the Chloroplasts" Cells 9, no. 1: 259. https://doi.org/10.3390/cells9010259