Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. FHL2 Depletion with siRNA-Pools
2.3. Animals and Bile Duct Ligation
2.4. Quantitative Real-Time-PCR Analysis
2.5. Protein Analysis
2.6. (Immono)Histological Analysis
2.7. Statistical Analysis
3. Results
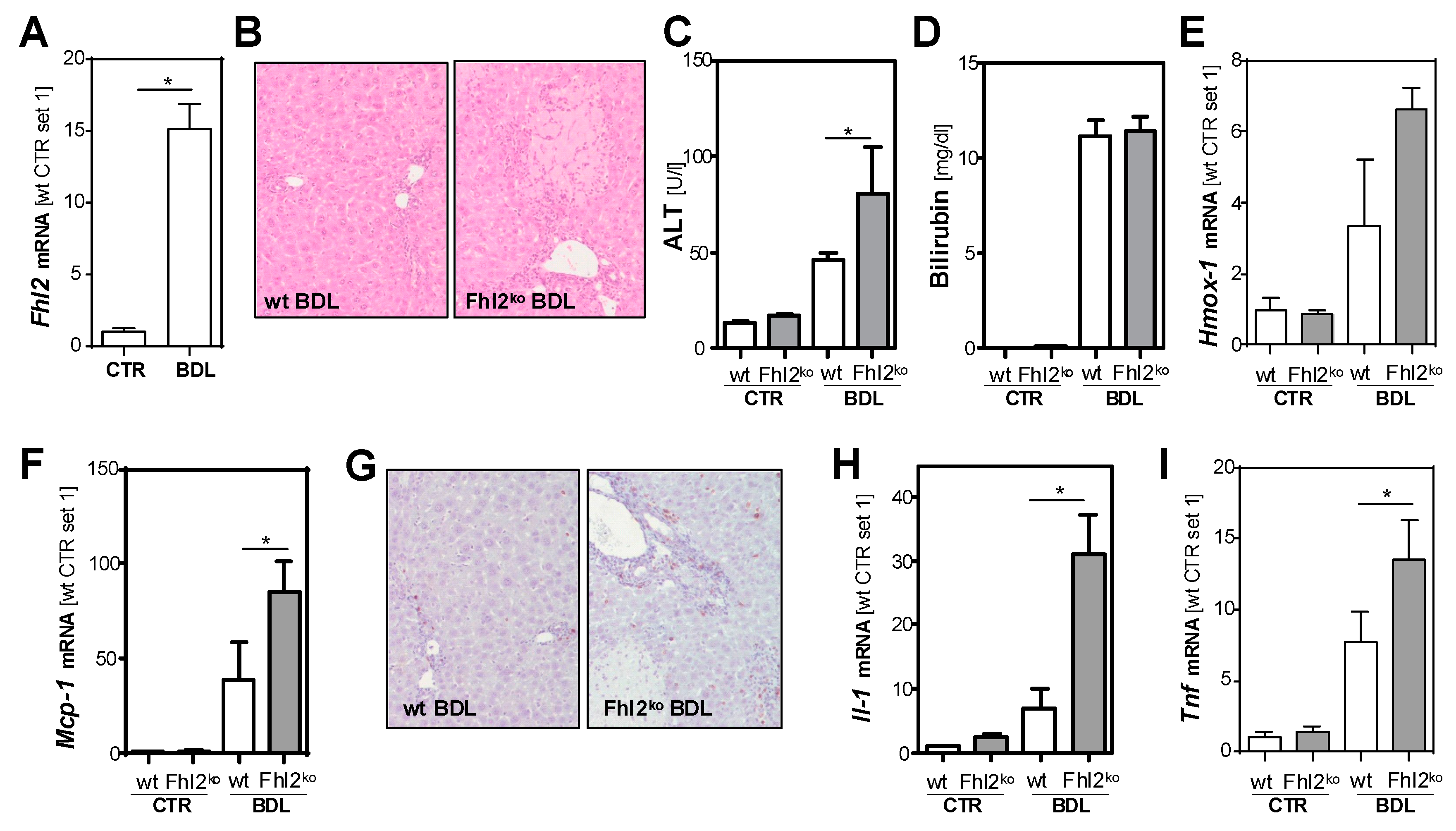
3.1. Fhl2 Deficiency Aggravates Hepatocellular Injury and Inflammation in Cholestatic Liver Injury
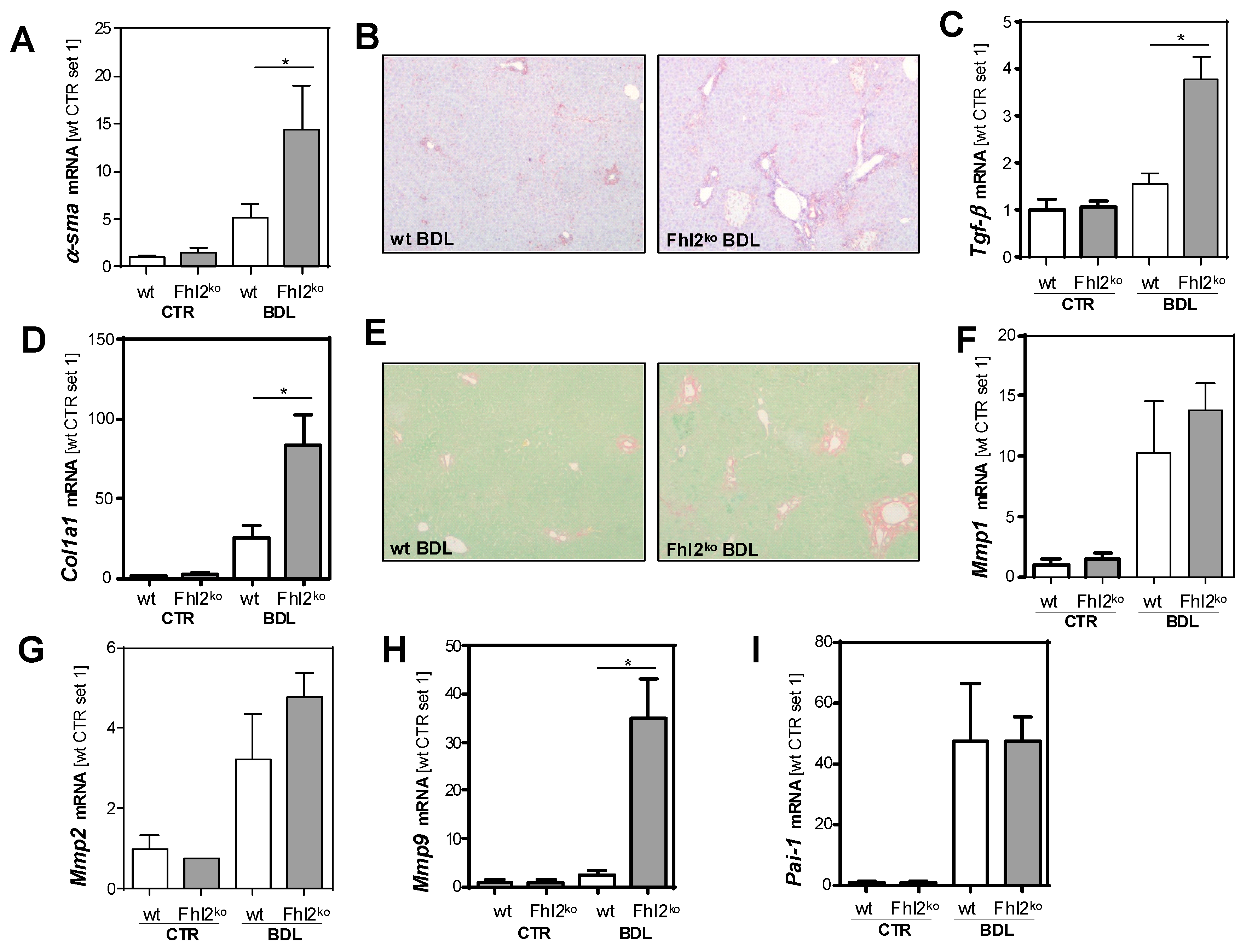
3.2. Fhl2 Deficiency Aggravates Hepatic Fibrosis in the Mouse Model of Bile Duct Ligation
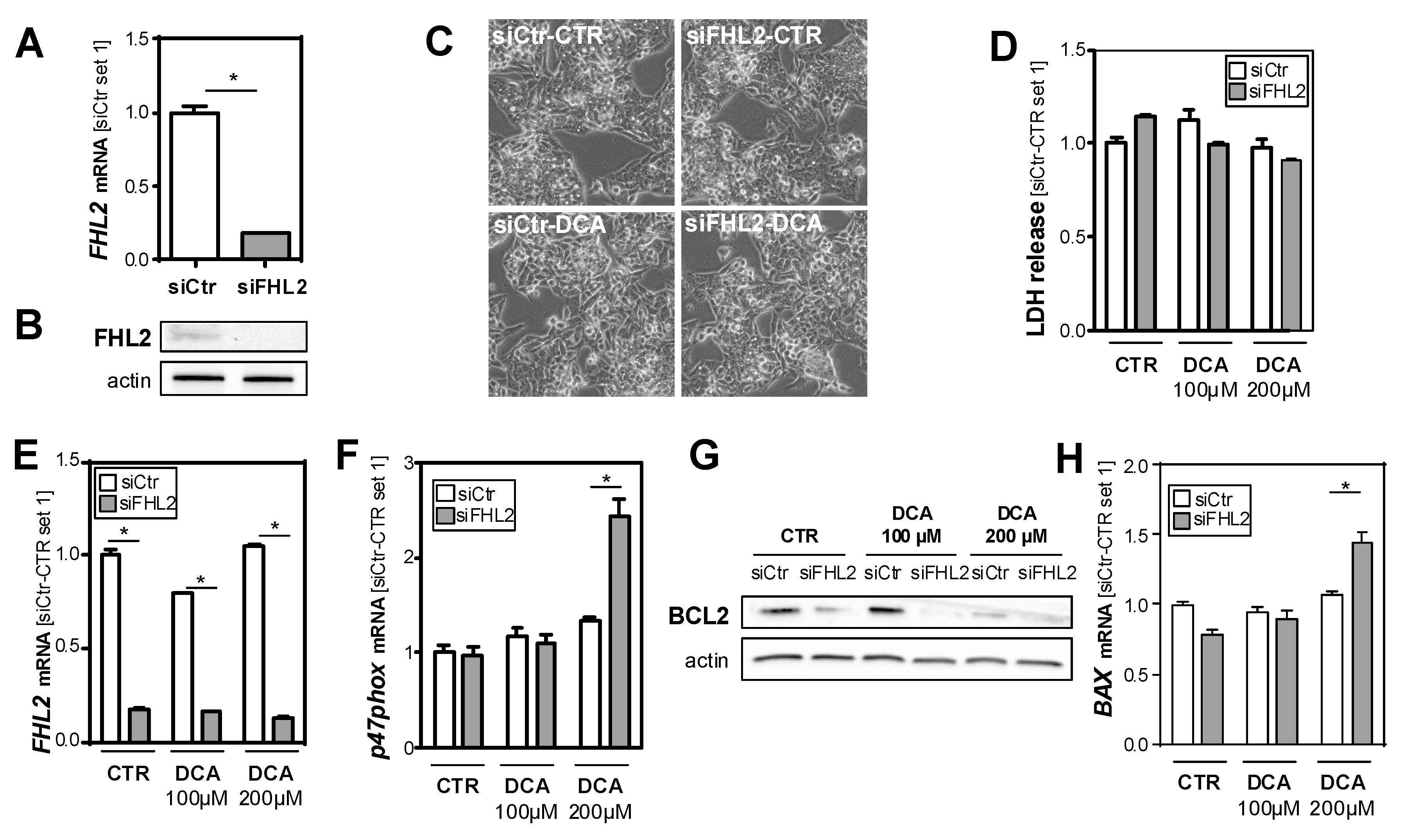
3.3. FHL2 Depletion Promotes Bile Acid-Induced Hepatocellular Injury In Vitro
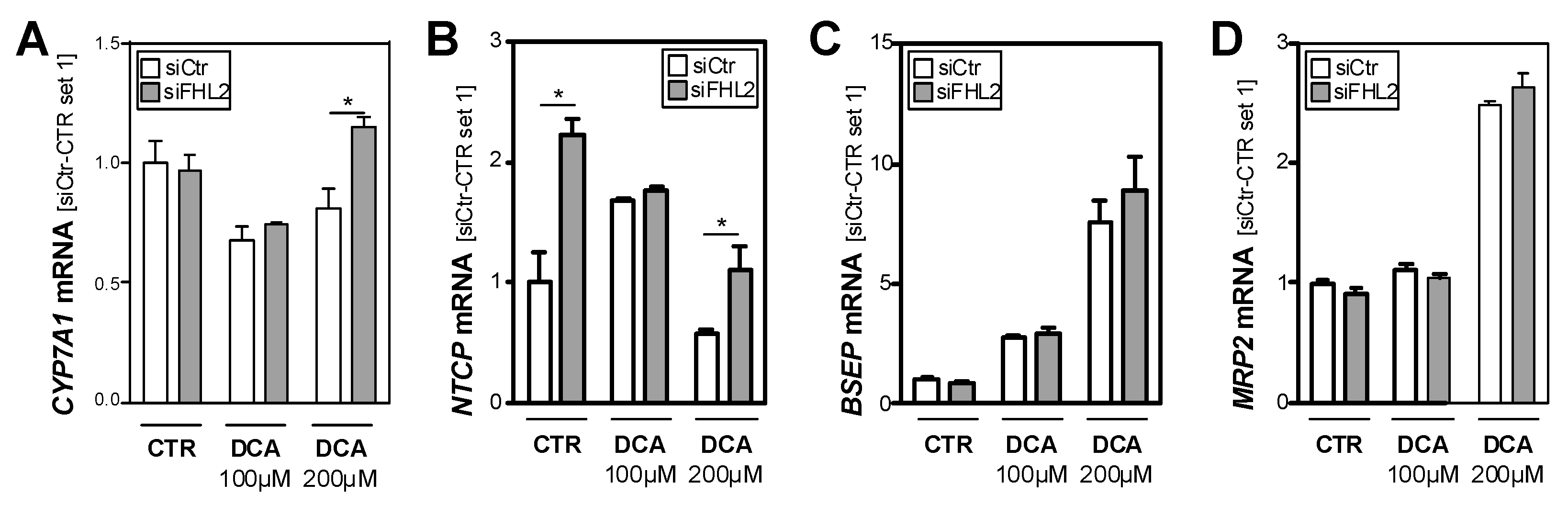
3.4. FHL2 Depletion Affects Expression of Key Enzymes of Bile Acid Metabolism
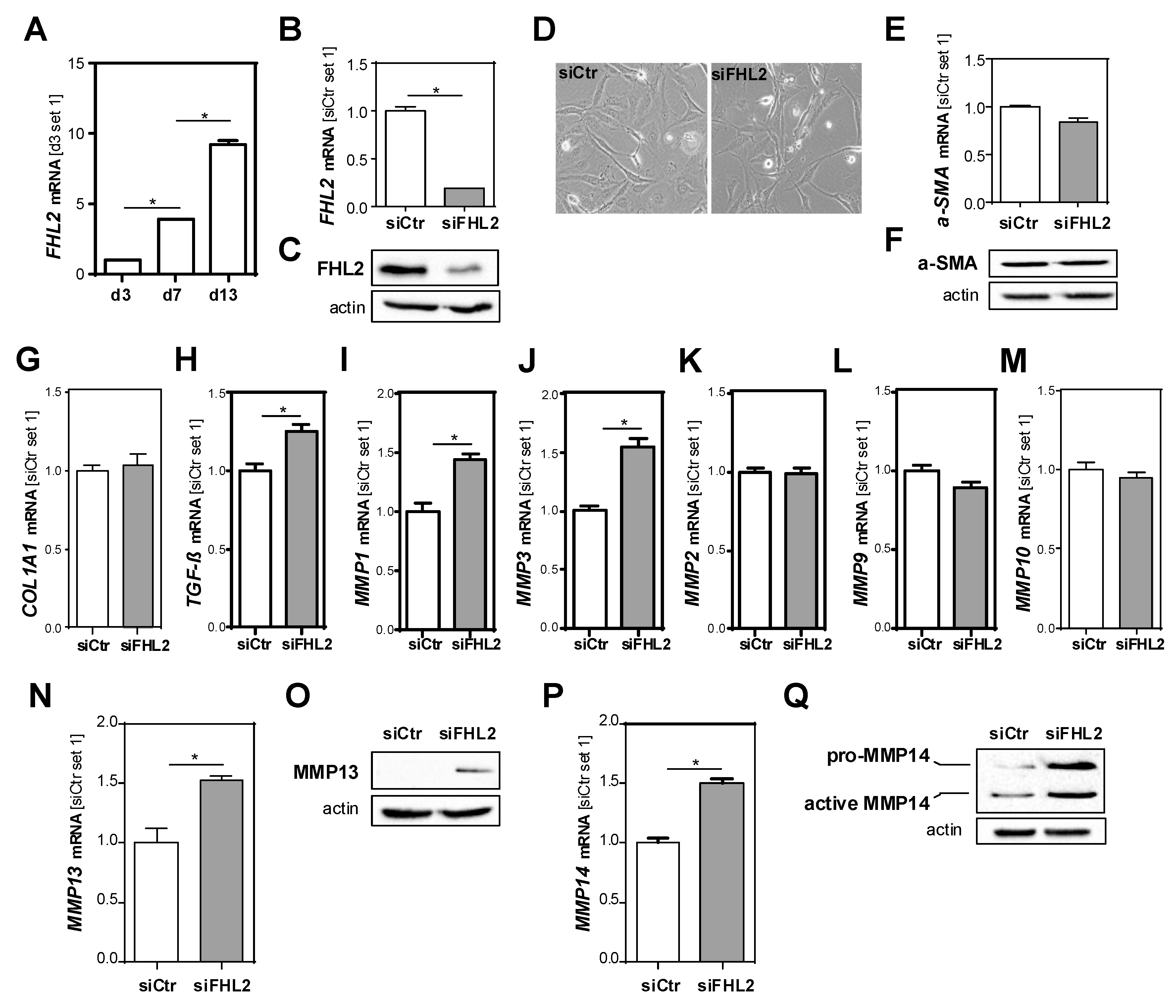
3.5. FHL2 Depletion Promotes Pro-Fibrogenic Gene Expression in Hepatic Stellate Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pellicoro, A.; Ramachandran, P.; Iredale, J.P.; Fallowfield, J.A. Liver fibrosis and repair: Immune regulation of wound healing in a solid organ. Nat. Rev. Immunol. 2014, 14, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.A.; Wallace, M.C.; Friedman, S.L. Pathobiology of liver fibrosis: A translational success story. Gut 2015, 64, 830–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ekstedt, M.; Hagstrom, H.; Nasr, P.; Fredrikson, M.; Stal, P.; Kechagias, S.; Hultcrantz, R. Fibrosis stage is the strongest predictor for disease-specific mortality in NAFLD after up to 33 years of follow-up. Hepatology 2015, 61, 1547–1554. [Google Scholar] [CrossRef] [Green Version]
- Angulo, P.; Kleiner, D.E.; Dam-Larsen, S.; Adams, L.A.; Bjornsson, E.S.; Charatcharoenwitthaya, P.; Mills, P.R.; Keach, J.C.; Lafferty, H.D.; Stahler, A.; et al. Liver Fibrosis, but No Other Histologic Features, Is Associated With Long-term Outcomes of Patients With Nonalcoholic Fatty Liver Disease. Gastroenterology 2015, 149, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.K.; Kurakula, K.; Koenis, D.S.; de Vries, C.J. Protein-protein interactions of the LIM-only protein FHL2 and functional implication of the interactions relevant in cardiovascular disease. Biochim. Biophys. Acta 2016, 1863, 219–228. [Google Scholar] [CrossRef]
- Johannessen, M.; Moller, S.; Hansen, T.; Moens, U.; Van Ghelue, M. The multifunctional roles of the four-and-a-half-LIM only protein FHL2. Cell Mol. Life Sci. 2006, 63, 268–284. [Google Scholar] [CrossRef]
- Verset, L.; Feys, L.; Trepant, A.L.; De Wever, O.; Demetter, P. FHL2: A scaffold protein of carcinogenesis, tumour-stroma interactions and treatment response. Histol. Histopathol. 2016, 31, 469–478. [Google Scholar] [CrossRef]
- Wixler, V. The role of FHL2 in wound healing and inflammation. FASEB J. 2019, 33, 7799–7809. [Google Scholar] [CrossRef]
- Huang, P.H.; Chen, C.Y.; Lin, C.P.; Wang, C.H.; Tsai, H.Y.; Lo, W.Y.; Leu, H.B.; Chen, J.W.; Lin, S.J.; Chu, P.H. Deletion of FHL2 gene impaired ischemia-induced blood flow recovery by modulating circulating proangiogenic cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Kirfel, J.; Pantelis, D.; Kabba, M.; Kahl, P.; Roper, A.; Kalff, J.C.; Buettner, R. Impaired intestinal wound healing in Fhl2-deficient mice is due to disturbed collagen metabolism. Exp. Cell Res. 2008, 314, 3684–3691. [Google Scholar] [CrossRef] [PubMed]
- Goltz, D.; Hittetiya, K.; Gevensleben, H.; Kirfel, J.; Diehl, L.; Meyer, R.; Buttner, R. Loss of the LIM-only protein Fhl2 impairs inflammatory reaction and scar formation after cardiac ischemia leading to better hemodynamic performance. Life Sci. 2016, 151, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Wixler, V.; Hirner, S.; Muller, J.M.; Gullotti, L.; Will, C.; Kirfel, J.; Gunther, T.; Schneider, H.; Bosserhoff, A.; Schorle, H.; et al. Deficiency in the LIM-only protein Fhl2 impairs skin wound healing. J. Cell Biol. 2007, 177, 163–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amann, T.; Maegdefrau, U.; Hartmann, A.; Agaimy, A.; Marienhagen, J.; Weiss, T.S.; Stoeltzing, O.; Warnecke, C.; Scholmerich, J.; Oefner, P.J.; et al. GLUT1 expression is increased in hepatocellular carcinoma and promotes tumorigenesis. Am. J. Pathol. 2009, 174, 1544–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhlbauer, M.; Bosserhoff, A.K.; Hartmann, A.; Thasler, W.E.; Weiss, T.S.; Herfarth, H.; Lock, G.; Scholmerich, J.; Hellerbrand, C. A novel MCP-1 gene polymorphism is associated with hepatic MCP-1 expression and severity of HCV-related liver disease. Gastroenterology 2003, 125, 1085–1093. [Google Scholar] [CrossRef]
- Thasler, W.E.; Weiss, T.S.; Schillhorn, K.; Stoll, P.T.; Irrgang, B.; Jauch, K.W. Charitable State-Controlled Foundation Human Tissue and Cell Research: Ethic and Legal Aspects in the Supply of Surgically Removed Human Tissue For Research in the Academic and Commercial Sector in Germany. Cell Tissue Bank. 2003, 4, 49–56. [Google Scholar] [CrossRef]
- Mahli, A.; Seitz, T.; Beckroge, T.; Freese, K.; Thasler, W.E.; Benkert, M.; Dietrich, P.; Weiskirchen, R.; Bosserhoff, A.; Hellerbrand, C. Bone Morphogenetic Protein-8B Expression is Induced in Steatotic Hepatocytes and Promotes Hepatic Steatosis and Inflammation In Vitro. Cells 2019, 8, 457. [Google Scholar] [CrossRef] [Green Version]
- Hannus, M.; Beitzinger, M.; Engelmann, J.C.; Weickert, M.T.; Spang, R.; Hannus, S.; Meister, G. siPools: Highly complex but accurately defined siRNA pools eliminate off-target effects. Nucleic Acids Res. 2014, 42, 8049–8061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huss, S.; Stellmacher, C.; Goltz, D.; Khlistunova, I.; Adam, A.C.; Trebicka, J.; Kirfel, J.; Buttner, R.; Weiskirchen, R. Deficiency in four and one half LIM domain protein 2 (FHL2) aggravates liver fibrosis in mice. BMC Gastroenterol. 2013, 13, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabele, E.; Muhlbauer, M.; Dorn, C.; Weiss, T.S.; Froh, M.; Schnabl, B.; Wiest, R.; Scholmerich, J.; Obermeier, F.; Hellerbrand, C. Role of TLR9 in hepatic stellate cells and experimental liver fibrosis. Biochem. Biophys. Res. Commun. 2008, 376, 271–276. [Google Scholar] [CrossRef]
- Sommer, J.; Mahli, A.; Freese, K.; Schiergens, T.S.; Kuecuekoktay, F.S.; Teufel, A.; Thasler, W.E.; Muller, M.; Bosserhoff, A.K.; Hellerbrand, C. Analysis of molecular mechanisms of 5-fluorouracil-induced steatosis and inflammation in vitro and in mice. Oncotarget 2017, 8, 13059–13072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, C.; Engelmann, J.C.; Saugspier, M.; Koch, A.; Hartmann, A.; Muller, M.; Spang, R.; Bosserhoff, A.; Hellerbrand, C. Increased expression of c-Jun in nonalcoholic fatty liver disease. Lab. Investig. 2014, 94, 394–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arndt, S.; Wacker, E.; Dorn, C.; Koch, A.; Saugspier, M.; Thasler, W.E.; Hartmann, A.; Bosserhoff, A.K.; Hellerbrand, C. Enhanced expression of BMP6 inhibits hepatic fibrosis in non-alcoholic fatty liver disease. Gut 2015, 64, 973–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, I.; Solis-Munoz, P.; Fernandez-Moreira, D.; Munoz-Yague, T.; Solis-Herruzo, J.A. In vitro treatment of HepG2 cells with saturated fatty acids reproduces mitochondrial dysfunction found in nonalcoholic steatohepatitis. Dis. Model Mech. 2015, 8, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, R.; Becker, S.; Keitel, V.; Eberle, A.; Grether-Beck, S.; Haussinger, D. Bile salt-induced apoptosis involves NADPH oxidase isoform activation. Gastroenterology 2005, 129, 2009–2031. [Google Scholar] [CrossRef]
- Govoni, K.E.; Baylink, D.J.; Chen, J.; Mohan, S. Disruption of four-and-a-half LIM 2 decreases bone mineral content and bone mineral density in femur and tibia bones of female mice. Calcif. Tissue Int. 2006, 79, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.Y.; Ouyang, X.; Chen, Y.; Soroka, C.J.; Wang, J.; Mennone, A.; Wang, Y.; Mehal, W.Z.; Jain, D.; Boyer, J.L. Bile acids initiate cholestatic liver injury by triggering a hepatocyte-specific inflammatory response. JCI Insight 2017, 2, e90780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hong, J.Y.; Rockwell, C.E.; Copple, B.L.; Jaeschke, H.; Klaassen, C.D. Effect of bile duct ligation on bile acid composition in mouse serum and liver. Liver Int. 2012, 32, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Kurakula, K.; Sommer, D.; Sokolovic, M.; Moerland, P.D.; Scheij, S.; van Loenen, P.B.; Koenis, D.S.; Zelcer, N.; van Tiel, C.M.; de Vries, C.J. LIM-only protein FHL2 is a positive regulator of liver X receptors in smooth muscle cells involved in lipid homeostasis. Mol. Cell Biol. 2015, 35, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Dahan, J.; Levillayer, F.; Xia, T.; Nouet, Y.; Werts, C.; Fanton d’Andon, M.; Adib-Conquy, M.; Cassard-Doulcier, A.M.; Khanna, V.; Chen, J.; et al. LIM-Only Protein FHL2 Is a Negative Regulator of Transforming Growth Factor beta1 Expression. Mol. Cell Biol. 2017, 37, e00636-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naim, A.; Pan, Q.; Baig, M.S. Matrix Metalloproteinases (MMPs) in Liver Diseases. J. Clin. Exp. Hepatol. 2017, 7, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Roderfeld, M. Matrix metalloproteinase functions in hepatic injury and fibrosis. Matrix Biol. 2018, 68, 452–462. [Google Scholar] [CrossRef]
- Kosters, A.; Karpen, S.J. The role of inflammation in cholestasis: Clinical and basic aspects. Semin. Liver Dis. 2010, 30, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Dahan, J.; Nouet, Y.; Jouvion, G.; Levillayer, F.; Adib-Conquy, M.; Cassard-Doulcier, A.M.; Tebbi, A.; Blanc, F.; Remy, L.; Chen, J.; et al. LIM-only protein FHL2 activates NF-kappaB signaling in the control of liver regeneration and hepatocarcinogenesis. Mol. Cell Biol. 2013, 33, 3299–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouet, Y.; Dahan, J.; Labalette, C.; Levillayer, F.; Julien, B.; Jouvion, G.; Cairo, S.; Vives, F.L.; Ribeiro, A.; Huerre, M.; et al. The four and a half LIM-only protein 2 regulates liver homeostasis and contributes to carcinogenesis. J. Hepatol. 2012, 57, 1029–1036. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| 18S | TCTGTGATGCCCTTAGATGTCC | CCATCCAATCGGTAGTAGCG |
| Human | ||
| α-SMA | CGTGGCTATTCCTTCGTTAC | TGCCAGCAGACTCCATCC |
| BAX | GGCCCACCAGCTCTGAGCAGA | GCCACGTGGGCGTCCCAAAGT |
| BCL2 | GCGGATTTGAATCTCTTTCTC | CACTAAACTGACTCCAGCTG |
| BSEP | TGCCCAGAATGGCCCTACA | CCAGCATTGCCCTGAAACCA |
| COL1A1 | CGGCTCCTGCTCCTCTT | GGGGCAGTTCTTGGTCTC |
| CYP7A1 | CCATAAGGTGTTGTGCCACGG | TCCGTGAGGGAATTCAAGGCA |
| FHL2 | GAAACTCACTGGTGGACAAGC | GTGGCAGATGAAGCAGGTCT |
| MMP1 | TCACCAAGGTCTCTGAGGGTCAAGC | GGATGCCATCAATGTCATCCTGAGC |
| MMP2 | GCTGGGAGCATGGCGATGGATACC | GGACAGAAGCCGTACTTGCCATCC |
| MMP3 | TGCTGTTTTTGAAGAATTTGGGTT | CAATTCACAGAGACTTAGGTGAAGA |
| MMP9 | TGCCTTTGGACACGCACG | CCTGGTTCAACTCACTCCGGG |
| MMP10 | GGGGGAAGACAGATATGGGT | CTGTTCAGTGCAATTCAAAAGC |
| MMP13 | TACCAGACTTCACGATGGCATTGCTG | AAAGTGGCTTTTGCCGGTGTAGGTG |
| MMP14 | GGAACCCTGTAGCTTTGTGTCTGTC | TCTCTACCCTCAACAAGATTAGATTCC |
| MRP2 | TCACATGTCCATCCACTGTTTCA | TGCTCAAAACAAGTGGCAGG |
| Mouse | ||
| α-sma | CCAGCCATCTTTCATTGGGAT | CCCCTGACAGGACGTTGTTA |
| Col1a1 | CTGTTCCAGGCAATCCACGA | ATCAGCTGGAGTTTCCGTGC |
| Fhl2 | ACTGCCTGACCTGCTTCTGT | TTGCCTGGTTATGAAAGAAAA |
| Hmox-1 | CACGCATATACCCGCTACCT | CCAGAGTGTTCATTCGAGCA |
| Il-1 | TGCCACCTTTTGACAGTGATG | AAGGTCCACGGGAAAGACAC |
| Mcp-1 | TGCAGGTCCCTGTCATGCTTC | TGGACCCATTCCTTCTTGGGG |
| Mmp1 | CTTGGCCACTCCCTAGGTCT | AGGGCTGGGTCACACTTCTC |
| Mmp2 | ATGGACAGCCCTGCAAGTTC | CAGTGGACATAGCGGTCTCG |
| Mmp9 | GTCCAGACCAAGGGTACAGC | CTGTCGGCTGTGGTTCAGTT |
| Pai-1 | ATGGGGCCGTGGAACAAGAA | AGGCGTGTCAGCTCGTCTAC |
| Tgf-β | CATTGCTGTCCCGTGCAGAG | CAGGCGTATCAGTGGGGGTC |
| Tnf | CCCTCACACTCAGATCATCTTCT | GCTACGACGTGGGCTACAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommer, J.; Dorn, C.; Gäbele, E.; Bataille, F.; Freese, K.; Seitz, T.; Thasler, W.E.; Büttner, R.; Weiskirchen, R.; Bosserhoff, A.; et al. Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury. Cells 2020, 9, 248. https://doi.org/10.3390/cells9010248
Sommer J, Dorn C, Gäbele E, Bataille F, Freese K, Seitz T, Thasler WE, Büttner R, Weiskirchen R, Bosserhoff A, et al. Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury. Cells. 2020; 9(1):248. https://doi.org/10.3390/cells9010248
Chicago/Turabian StyleSommer, Judith, Christoph Dorn, Erwin Gäbele, Frauke Bataille, Kim Freese, Tatjana Seitz, Wolfgang E. Thasler, Reinhard Büttner, Ralf Weiskirchen, Anja Bosserhoff, and et al. 2020. "Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury" Cells 9, no. 1: 248. https://doi.org/10.3390/cells9010248