N-Cadherin Is Critical for the Survival of Germ Cells, the Formation of Steroidogenic Cells, and the Architecture of Developing Mouse Gonads

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Genotyping
2.3. RNA Isolation from the Gonads and Real-Time Quantitative PCR
2.4. Gonadal Cell Isolation and Sorting
2.5. Histology and Immunohistochemistry
2.6. Electron Microscopy
2.7. Measurements
3. Results
3.1. The Effectiveness of Genetic Knockout
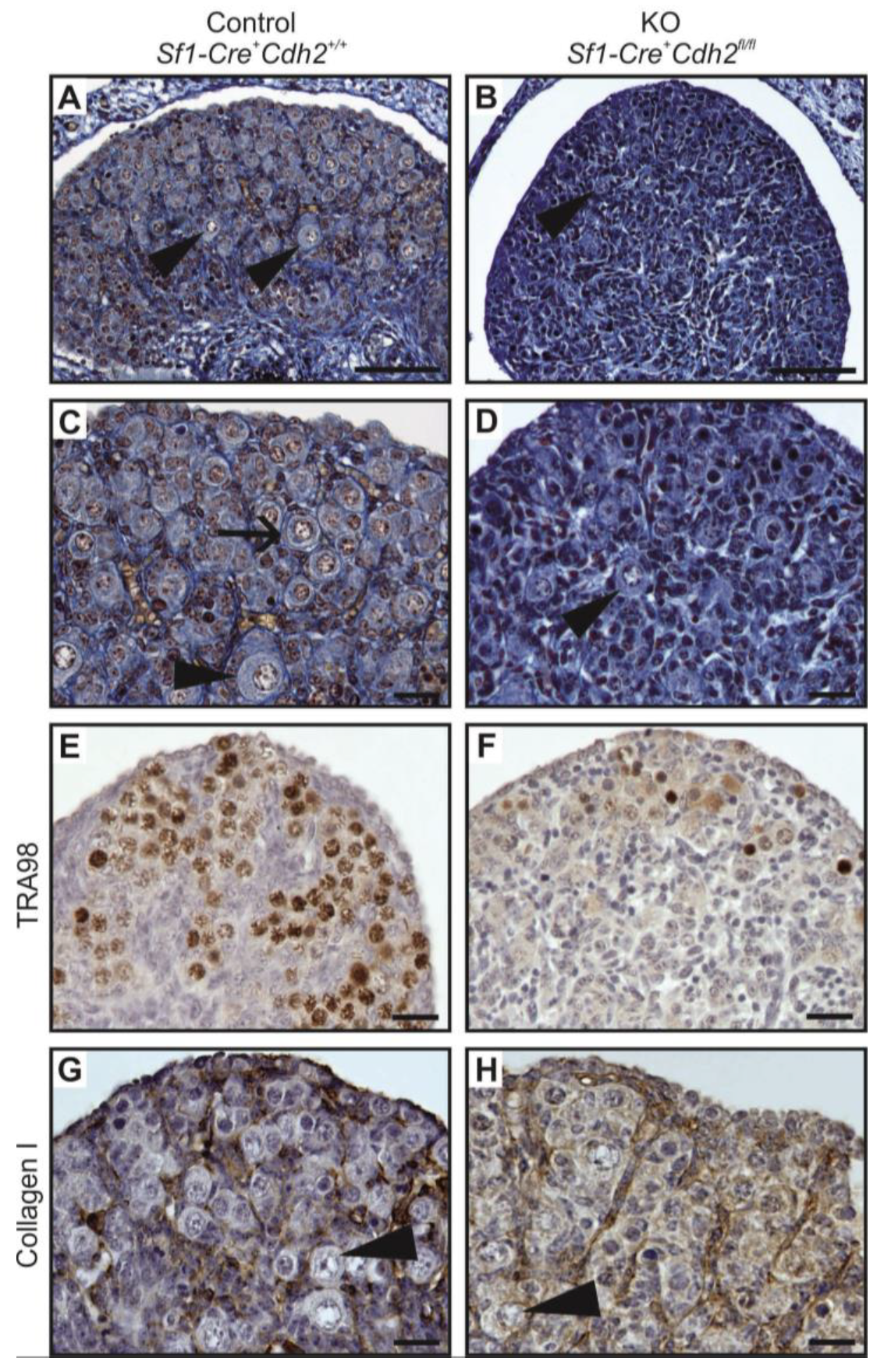
3.2. Knockout of N-Cadherin (Cdh2) in SF1+ Somatic Cells of Developing Testis
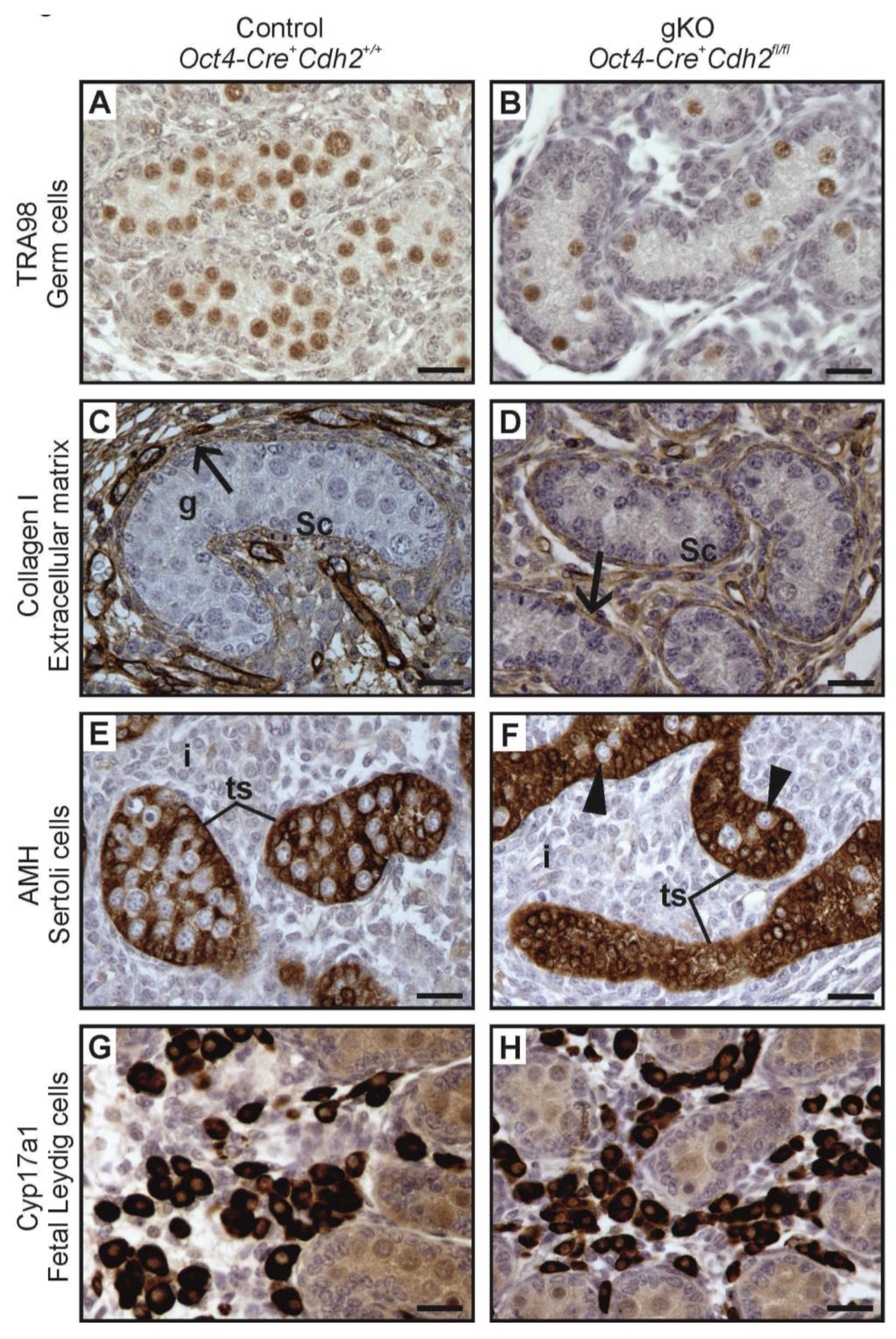
3.3. N-Cadherin Knockout in SF1+ Somatic Cells Affects Steroidogenic Fetal Leydig Cells (FLCs)
3.4. The Effect of N-Cadherin Knockout in Somatic Cells on Sex Determination
3.5. Knockout of N-Cadherin (Cdh2) in SF1+ Somatic Cells of the Developing Ovary
3.6. Knockout of N-Cadherin (Cdh2) in OCT4+ Germ Cells in Developing Testes
3.7. Knockout of N-Cadherin (Cdh2) in OCT4+ Germ Cells in Developing Ovaries
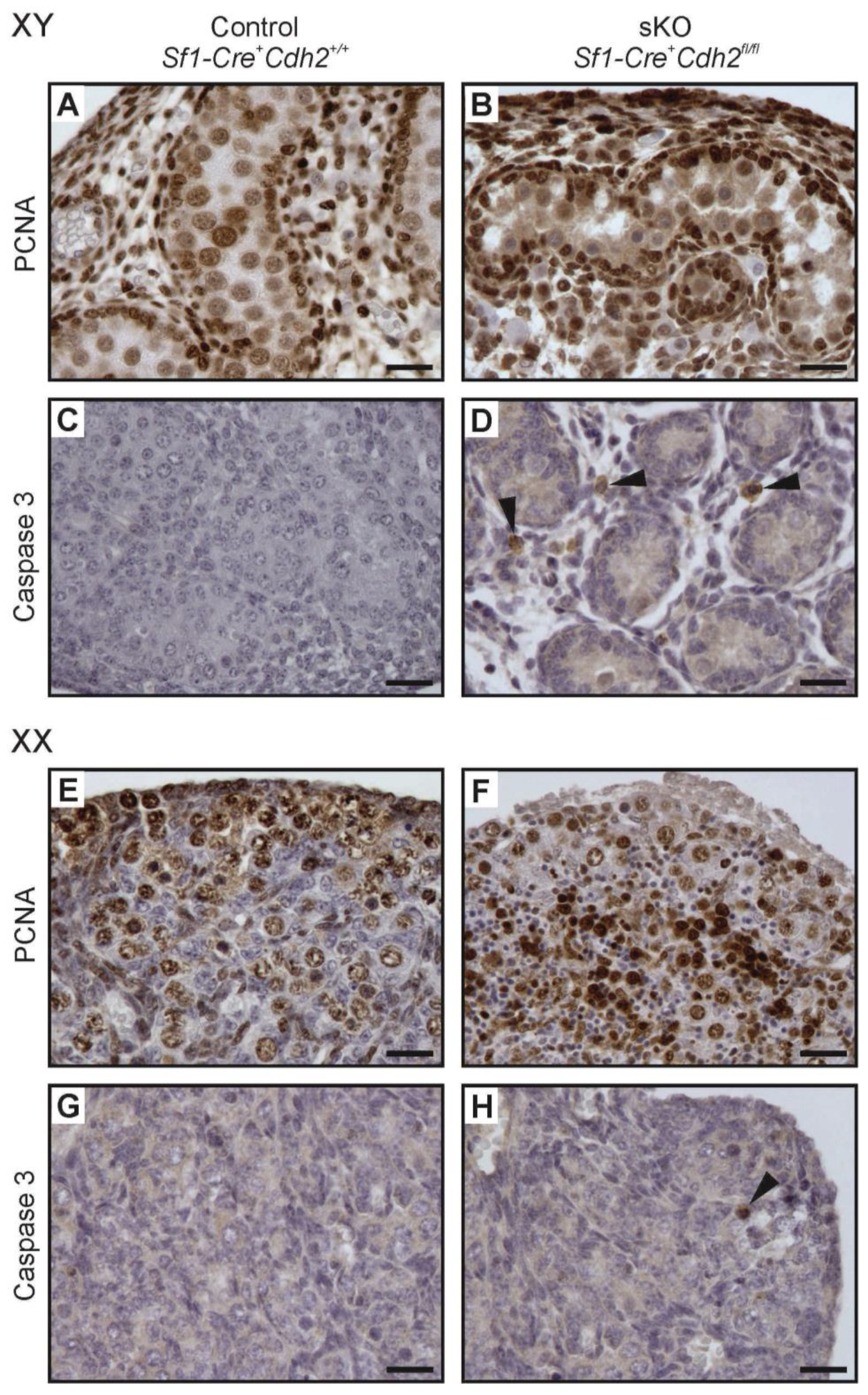
3.8. Proliferation and Apoptosis in N-Cadherin Knockouts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Piprek, R.P.; Kloc, M.; Kubiak, J.Z. Early Development of the gonads: Origin and differentiation of the somatic cells of the genital ridges. Results Probl. Cell Differ. 2016, 58, 1–22. [Google Scholar] [PubMed]
- Albrecht, K.H.; Eicher, E.M. Evidence that Sry is expressed in pre-Sertoli cells and Sertoli and granulosa cells have a common precursor. Dev. Biol. 2001, 240, 92–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFalco, T.; Takahashi, S.; Capel, B. Two distinct origins for Leydig cell progenitors in the fetal testis. Dev. Biol. 2011, 352, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. Cell adhesion molecules expression pattern indicates that somatic cells arbitrate gonadal sex of differentiating bipotential fetal mouse gonad. Mech. Dev. 2017, 147, 17–27. [Google Scholar] [CrossRef]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. Tissue-specific knockout of E-cadherin (Cdh1) in developing mouse gonads causes germ cells loss. Reproduction 2019, 158, 147–157. [Google Scholar] [CrossRef]
- Fleming, A.; Ghahramani, N.; Zhu, M.X.; Délot, E.C.; Vilain, E. Membrane β-Catenin and adherens junctions in early gonadal patterning. Dev. Dyn. 2012, 241, 1782–1798. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.H.; DePhilip, R.M. Sex-dependent expression of placental (P)-cadherin during mouse gonadogenesis. Anat. Rec. 1996, 246, 535–544. [Google Scholar] [CrossRef]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef] [Green Version]
- Newton, S.C.; Blaschuk, O.W.; Millette, C.F. N-cadherin mediates Sertoli cell-spermatogenic cell adhesion. Dev. Dyn. 1993, 197, 1–13. [Google Scholar] [CrossRef]
- Yan, H.H.N.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells. FASEB J. 2008, 22, 1945–1959. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.B.; Walker, W.H. The regulation of spermatogenesis by androgens. Semin Cell Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Ma, T.; Zhang, Y.; Zhang, H.; Yin, S.; Zheng, W.; Wang, L.; Wang, Z.; Khan, M.; Sheikh, S.W. Specific deletion of Cdh2 in Sertoli cells leads to altered meiotic progression and subfertility of mice. Biol. Reprod. 2015, 92, 79. [Google Scholar] [CrossRef] [PubMed]
- Piprek, R.P.; Kloc, M.; Tassan, J.P.; Kubiak, J.Z. Development of Xenopus laevis bipotential gonads into testis or ovary is driven by sex-specific cell-cell interactions, proliferation rate, cell migration and deposition of extracellular matrix. Dev. Biol. 432, 298–310. [CrossRef]
- Radice, G.L.; Rayburn, H.; Matsunami, H.; Knudsen, K.A.; Takeichi, M.; Hynes, R.O. Developmental defects in mouse embryos lacking N-cadherin. Dev. Biol. 1997, 181, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, H.; Zigman, J.M.; Ye, C.; Lee, C.E.; McGovern, R.A.; Tang, V.; Kenny, C.D.; Christiansen, L.M.; White, R.D.; Edelstein, E.A. Leptin directly activates SF1 neurons in the VMH, and this action by leptin is required for normal body-weight homeostasis. Neuron 2006, 49, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greder, L.V.; Gupta, S.; Li, S.; Abedin, M.J.; Sajini, A.; Segal, Y.; Slack, J.M.W.; Dutton, J.R. Analysis of endogenous Oct4 activation during induced pluripotent stem cell reprogramming using an inducible Oct4 lineage label. Stem Cells 2012, 30, 2596–2601. [Google Scholar] [CrossRef] [Green Version]
- Kostetskii, I.; Li, J.; Xiong, Y.; Zhou, R.; Ferrari, V.A.; Patel, V.V.; Molkentin, J.D.; Radice, G.L. Induced deletion of the N-cadherin gene in the heart leads to dissolution of the intercalated disc structure. Circ. Res. 2005, 96, 346–354. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, L.; Truong, V.; Palmer, J.S.; Wilhelm, D. Novel PCR assay for determining the genetic sex of mice. Sex. Dev. 2013, 7, 207–211. [Google Scholar] [CrossRef]
- Svingen, T.; Spiller, C.M.; Kashimada, K.; Harley, V.R.; Koopman, P. Identification of suitable normalizing genes for quantitative real-time RT-PCR analysis of gene expression in fetal mouse gonads. Sex. Dev. 2009, 3, 194–204. [Google Scholar] [CrossRef]
- Durcova-Hills, G.; Tokunaga, T.; Kurosaka, S.; Yamaguchi, M.; Takahashi, S.; Imai, H. Immunomagnetic isolation of primordial germ cells and the establishment of embryonic germ cell lines in the mouse. Cloning 1999, 1, 217–224. [Google Scholar] [CrossRef]
- Medrano, J.V.; Marqués-Marí, A.I.; Aguilar, C.E.; Riboldi, M.; Garrido, N.; Martínez-Romero, A.; O’Connor, E.; Gil-Salom, M.; Simón, C. Comparative analysis of the germ cell markers c-KIT, SSEA-1 and VASA in testicular biopsies from secretory and obstructive azoospermias. Mol. Hum. Reprod. 2010, 16, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiernan, J.A. Staining with dyes in one or two colours. In Histological and Histochemical Methods: Theory and Practice, 5th ed.; Kiernan, J.A., Ed.; Scion Publishing: Banbury, UK, 2015; pp. 137–169. [Google Scholar]
- Piprek, R.P.; Pecio, A.; Kubiak, J.Z.; Szymura, J.M. Differential effects of busulfan on gonadal development in five divergent anuran species. Reprod. Toxicol. 2012, 34, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, S.; Karnowsky, M.J. Formaldehyde-glutaraldehyde fixatives containing trinitro compounds. J. Cell Biol. 1968, 36, 168. [Google Scholar]
- Piprek, R.P.; Pecio, A.; Szymura, J.M. Differentiation and development of gonads in the yellow-bellied toad, Bombina variegata L., 1758 (Amphibia: Anura: Bombinatoridae). Zoolog. Sci. 2010, 27, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Piprek, R.P. Molecular mechanisms underlying female sex determination – antagonism between female and male pathway. Folia Biol. 2009, 57, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Rios-Rojas, C.; Bowles, J.; Koopman, P. On the role of germ cells in mammalian gonad development: Quiet passengers or back-seat drivers? Reproduction 2015, 149, R181–R191. [Google Scholar] [CrossRef] [Green Version]
- Peluso, J.J.; Pappalardo, A.; Trolice, M.P. N-cadherin-mediated cell contact inhibits granulosa cell apoptosis in a progesterone-independent manner. Endocrinology 1996, 137, 1196–1203. [Google Scholar] [CrossRef]
- Nguyen, P.T.; Nguyen, D.; Chea, C.; Miyauchi, M.; Fujii, M.; Takata, T. Interaction between N-cadherin and decoy receptor-2 regulates apoptosis in head and neck cancer. Oncotarget 2018, 9, 31516–31530. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.L.; Adams, D.G.; Vaillancourt, R.R.; Heimark, R.L. Signal transduction from N-cadherin increases Bcl-2. Regulation of the phosphatidylinositol 3-kinase/Akt pathway by homophilic adhesion and actin cytoskeletal organization. J. Biol. Chem. 2002, 277, 32905–32914. [Google Scholar] [CrossRef] [Green Version]
- Hazan, R.B.; Qiao, R.; Keren, R.; Badano, I.; Suyama, K. Cadherin switch in tumor progression. Ann. N. Y. Acad. Sci. 2004, 1014, 155–163. [Google Scholar] [CrossRef]
- Soh, B.S.; Buac, K.; Xu, H.; Li, E.; Ng, S.Y.; Wu, H.; Chmielowiec, J.; Jiang, X.; Bu, L.; Li, R.A.; et al. N-cadherin prevents the premature differentiation of anterior heart field progenitors in the pharyngeal mesodermal microenvironment. Cell Res. 2014, 24, 1420–1432. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; He, W.; Lin, N.; Wang, X.; Fan, Q.X. Downregulation of N-cadherin expression inhibits invasiveness, arrests cell cycle and induces cell apoptosis in esophageal squamous cell carcinoma. Cancer Invest. 2010, 28, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Koutsouki, E.; Beeching, C.A.; Slater, S.C.; Blaschuk, O.W.; Sala-Newby, G.B.; George, S.J. N-cadherin-dependent cell-cell contacts promote human saphenous vein smooth muscle cell survival. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 982–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnaud, G.; Gonelle-Gispert, C.; Morel, P.; Giovannoni, L.; Muller, Y.D.; Meier, R.; Borot, S.; Berney, T.; Bosco, D. Cadherin engagement protects human β-cells from apoptosis. Endocrinology 2011, 152, 4601–4609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haÿ, E.; Nouraud, A.; Marie, P.J. N-cadherin negatively regulates osteoblast proliferation and survival by antagonizing Wnt, ERK and PI3K/Akt signalling. PLoS ONE. 2009, 4, e8284. [Google Scholar] [CrossRef]
- Peluso, J.J.; Pappalardo, A.; Fernandez, G. E-cadherin-mediated cell contact prevents apoptosis of spontaneously immortalized granulosa cells by regulating Akt kinase activity. Biol. Reprod. 2001, 64, 1183–1190. [Google Scholar] [CrossRef] [Green Version]
- Galaz, S.; Espada, J.; Stockert, J.C.; Pacheco, M.; Sanz-Rodríguez, F.; Arranz, R.; Rello, S.; Cañete, M.; Villanueva, A.; Esteller, M.; et al. Loss of E-cadherin mediated cell-cell adhesion as an early trigger of apoptosis induced by photodynamic treatment. J. Cell Physiol. 2005, 205, 86–96. [Google Scholar] [CrossRef]
- Fouquet, S.; Lugo-Martínez, V.H.; Faussat, A.M.; Renaud, F.; Cardot, P.; Chambaz, J.; Pinçon-Raymond, M.; Thenet, S. Early loss of E-cadherin from cell-cell contacts is involved in the onset of anoikis in enterocytes. J. Biol. Chem. 2004, 279, 43061–43069. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.C.; Suriano, G.; Mendes, N.; Gomes, B.; Wen, X.; Carneiro, F.; Seruca, R.; Machado, J.C. E-cadherin impairment increases cell survival through Notch-dependent upregulation of Bcl-2. Hum. Mol. Genet. 2012, 21, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Marsters, S.; Ye, X.; Luis, E.; Gonzalez, L.; Ashkenazi, A. E-cadherin couples death receptors to the cytoskeleton to regulate apoptosis. Mol. Cell. 2014, 54, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.J.; Telegan, B.; Lavker, R.M.; Wheelock, M.J. E-cadherin and P-cadherin have partially redundant roles in human epidermal stratification. Cell Tissue Res. 1997, 288, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Hummel, T.; Zipursky, S.L. Afferent induction of olfactory glomeruli requires N-cadherin. Neuron 2004, 42, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Herman, T.; Clandinin, T.R.; Lee, R.; Zipursky, S.L. N-cadherin regulates target specificity in the Drosophila visual system. Neuron 2001, 30, 437–450. [Google Scholar] [CrossRef] [Green Version]
- Nern, A.; Zhu, Y.; Zipursky, S.L. Local N-cadherin interactions mediate distinct steps in the targeting of lamina neurons. Neuron 2008, 58, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, S.; Caldwell, J.C.; Eberl, D.F.; Clandinin, T.R. Drosophila N-cadherin mediates an attractive interaction between photoreceptor axons and their targets. Nat. Neurosci. 2005, 8, 443–450. [Google Scholar] [CrossRef]
- Zhu, H.; Luo, L. Diverse functions of N-cadherin in dendritic and axonal terminal arborization of olfactory projection neurons. Neuron 2004, 42, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, T.; Neuert, H.; Clandinin, T.R. A network of cadherin-mediated interactions polarizes growth cones to determine targeting specificity. Cell 2013, 154, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Borum, K. Oogenesis in the mouse. A study of the meiotic prophase. Exp. Cell Res. 1961, 24, 495–507. [Google Scholar] [CrossRef]
- 50 Pepling, M.E.; Spradling, A.C. Mouse ovarian germ cell cysts undergo programmed breakdown to form primordial follicles. Dev. Biol. 2001, 234, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Pepling, M.E.; Sundman, E.A.; Patterson, N.L.; Gephardt, G.W.; Medico, L.; Wilson, K.I. Differences in oocyte development and estradiol sensitivity among mouse strains. Reproduction 2010, 139, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Wang, Y.; Zhang, T.; He, M.; Sun, G.; Wen, J.; Yan, H.; Cai, H.; Yong, C.; Xi, G. JAK signaling regulates germline cyst breakdown and primordial follicle formation in mice. Biol. Open 2018, 7, bio029470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, R.; Tabata, S.; Nakanishi, Y.; Ohsako, S.; Kurohmaru, M.; Hayashi, Y. Reproliferation and relocation of mouse male germ cells (gonocytes) during prespermatogenesis. Anat. Rec. 2000, 258, 210–220. [Google Scholar] [CrossRef]
- Kumar, T.R.; Palapattu, G.; Wang, P.; Woodruff, T.K.; Boime, I.; Byrne, M.C.; Matzuk, M.M. Transgenic models to study gonadotropin function: The role of follicle-stimulating hormone in gonadal growth and tumorigenesis. Mol. Endocrinol. 1999, 13, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Themmen, A.P.N.; Huhtaniemi, I.T. Mutations of gonadotropins and gonadotropin receptors: Elucidating the physiology and pathophysiology of pituitary-gonadal function. Endocr. Rev. 2000, 21, 551–583. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Mild Phenotype | Severe Phenotype | |
|---|---|---|---|
| Testis size | Normal | Smaller | Smaller |
| Testis cord structure |  |  |  |
| Testis cords | Normal, thick | Smaller, irregular shape | Disrupted |
| Germ cells (g) | Normal number of germ cells | Singular germ cells or no germ cells | Small number or no germ cells |
| Sertoli cells (Sc) | Continuous monolayer of Sertoli cells | Continuous monolayer of Sertoli cells | Disrupted monolayer of Sertoli cells |
| Basement membrane (bm) | Continuous | Continuous | Discontinuous |
| Fetal Leydig cells (fLc) | Normal number | Lower number | Lower number |
| Proliferation | Normal (high level) | Normal (high level) | Normal (high level) |
| Apoptosis | Normal (low level) | Increased | Increased |
| Control | Knockout Phenotype | |
|---|---|---|
| Ovary size | Normal | Smaller |
| Ovigerous cord structure |  |  |
| Ovigerous cords | Normal, irregular | Smaller, straight |
| Germ cells (g) | Normal number of germ cells | Lower number of germ cells |
| Follicular cells (fc) | Present | Present |
| Basement membrane (bm) | Continuous | Continuous |
| Proliferation | Normal (high level) | Normal (high level) |
| Apoptosis | Normal (low level) | Increased |
| Control | Mutant Phenotype | |
|---|---|---|
| Testis size | Normal | Smaller |
| Testis cords | Normal | Smaller |
| Germ cells | Normal number of germ cells | Lower number of germ cells |
| Sertoli cells | Continuous monolayer of Sertoli cells | Continuous monolayer of Sertoli cells |
| Basement membrane | Continuous | Continuous |
| Fetal Leydig cells | Normal number | Normal number |
| Proliferation | Normal (high level) | Normal (high level) |
| Apoptosis | Normal (low level) | Increased |
| Control | Mutant Phenotype | |
|---|---|---|
| Ovary size | Normal | Smaller |
| Ovigerous cords | Normal | Smaller |
| Germ cells | Normal number of germ cells | Lower number of germ cells |
| Follicular cells | Present | Present |
| Basement membrane | Continuous | Continuous |
| Proliferation | Normal (high level) | Normal (high level) |
| Apoptosis | Normal (low level) | Increased |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. N-Cadherin Is Critical for the Survival of Germ Cells, the Formation of Steroidogenic Cells, and the Architecture of Developing Mouse Gonads. Cells 2019, 8, 1610. https://doi.org/10.3390/cells8121610
Piprek RP, Kolasa M, Podkowa D, Kloc M, Kubiak JZ. N-Cadherin Is Critical for the Survival of Germ Cells, the Formation of Steroidogenic Cells, and the Architecture of Developing Mouse Gonads. Cells. 2019; 8(12):1610. https://doi.org/10.3390/cells8121610
Chicago/Turabian StylePiprek, Rafal P., Michal Kolasa, Dagmara Podkowa, Malgorzata Kloc, and Jacek Z. Kubiak. 2019. "N-Cadherin Is Critical for the Survival of Germ Cells, the Formation of Steroidogenic Cells, and the Architecture of Developing Mouse Gonads" Cells 8, no. 12: 1610. https://doi.org/10.3390/cells8121610