
Generation of Highly Functional Hepatocyte-like Organoids from Human Adipose-Derived Mesenchymal Stem Cells Cultured with Endothelial Cells
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. ADSC Isolation
2.2. HLC and H-Organoid Differentiation
2.3. Morphological Analysis
2.4. Quantitative Reverse Transcription–PCR (qRT-PCR) Analysis
2.5. Fluorescence-Activated Cell Sorting (FACS) Analysis
2.6. Live/Dead Cell Staining
2.7. Immunofluorescence Staining
2.8. CYP3A4 Activity Assay
2.9. Ammonium Metabolism Assay
2.10. Statistical Analysis
3. Results
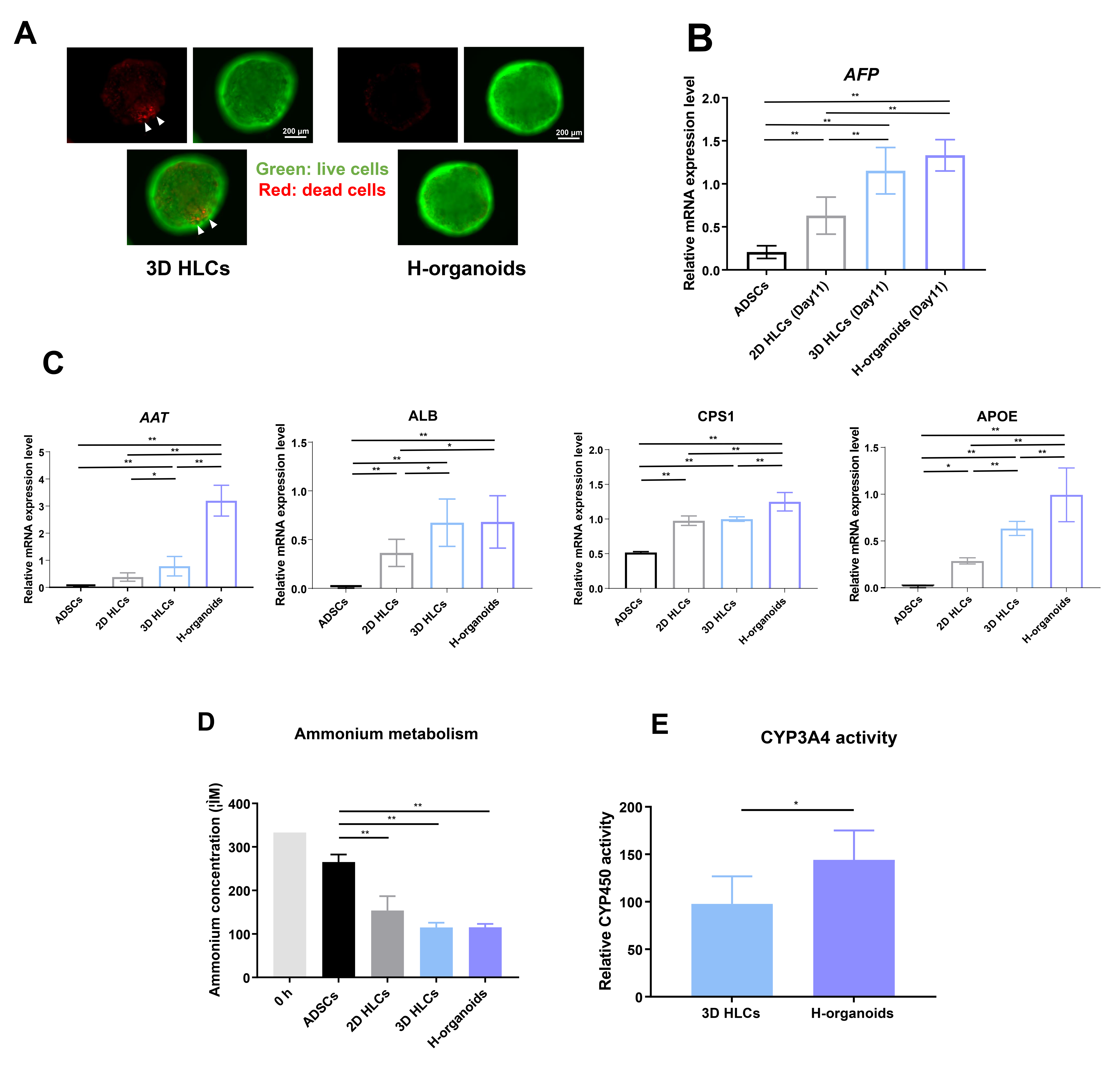
3.2. H-Organoids Present with Better Liver-like Functions In Vitro
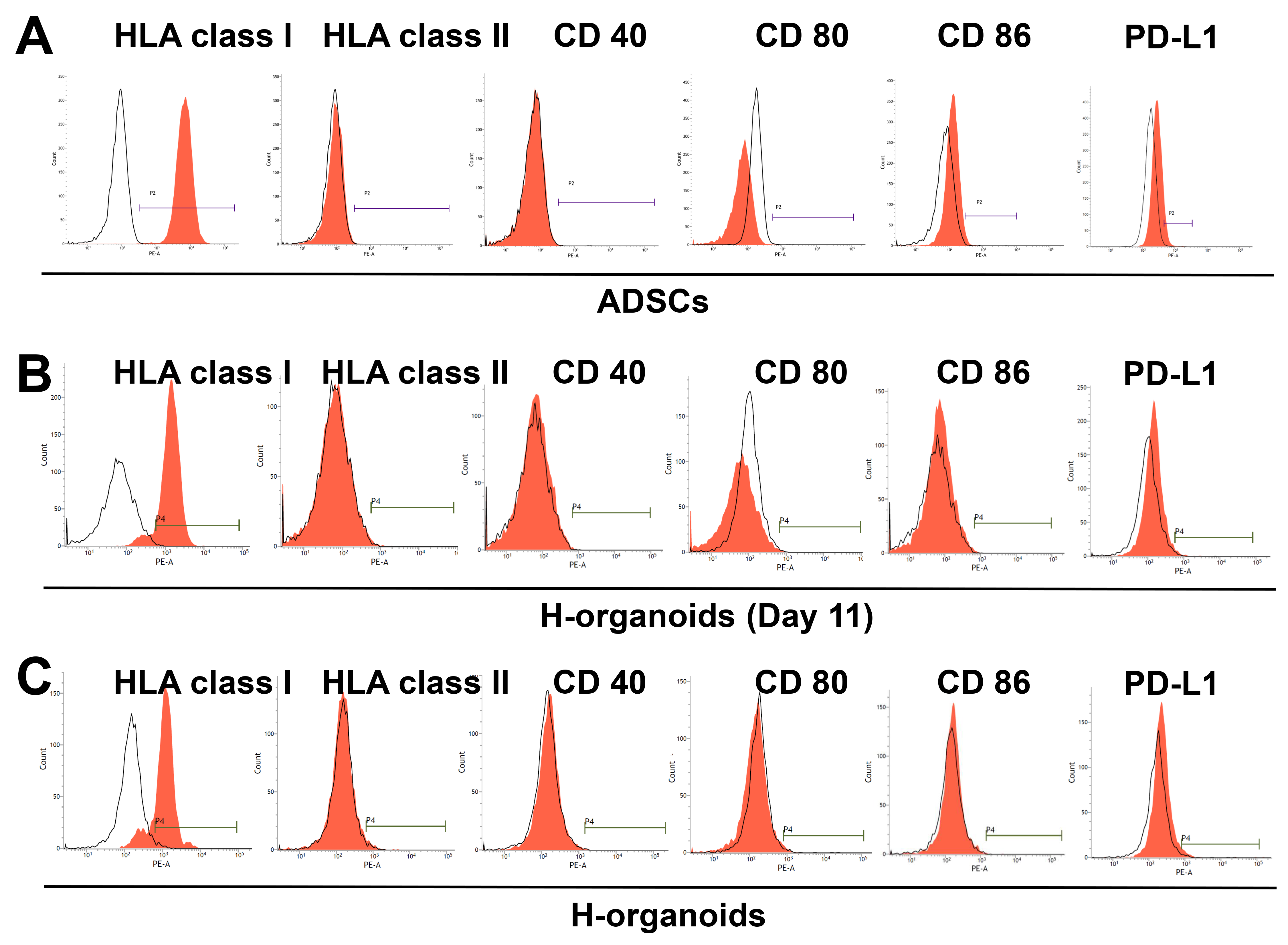
3.3. Immunogenicity of H-Organoids during Differentiation from ADSCs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| alpha-1 antitrypsin | AAT |
| adipose-derived mesenchymal stem cells | ADSCs |
| α-fetoprotein | AFP |
| apolipoprotein E | APOE |
| bovine serum albumin | BSA |
| carbamoyl-phosphate synthase 1 | CPS1 |
| cytochrome P | CYP |
| Hanks’ Balanced Salt Solution | HBSS |
| hematoxylin–eosin | HE |
| hepatocyte nuclear factor 4 alpha | HNF4α |
| hepatocyte-like cells | HLCs |
| hepatocyte-like organoids | H-organoids |
| human umbilical vein endothelial cells | HUVECs |
| human leukocyte antigen | HLA |
| human umbilical vein endothelial cells | HUVECs |
| immunofluorescence | IF |
| induced pluripotent stem cells | iPSCs |
| phosphate-buffered saline | PBS |
| reverse transcription–PCR | qRT-PCR |
| three-dimensional | 3D |
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Lee, W.M.; Squires, R.H., Jr.; Nyberg, S.L.; Doo, E.; Hoofnagle, J.H. Acute liver failure: Summary of a workshop. Hepatology 2008, 47, 1401–1415. [Google Scholar] [CrossRef]
- Saito, Y.; Ikemoto, T.; Yuji Morine, Y.; Shimada, M. Current status of hepatocyte-like cell therapy from stem cells. Surg. Today 2021, 51, 340–349. [Google Scholar] [CrossRef]
- Saito, Y.; Ikemoto, T.; Tokuda, K.; Miyazaki, K.; Yamada, S.; Imura, S.; Miyake, M.; Morine, Y.; Oyadomari, S.; Shimada, M. Effective three-dimensional culture of hepatocyte-like cells generated from human adipose-derived mesenchymal stem cells. J. Hepatobiliary Pancreat. Sci. 2021, 28, 705–715. [Google Scholar] [CrossRef]
- Afonso, M.B.; Marques, V.; Mil, S.W.C.; Rodrigues, C.M.P. Human liver organoids: From generation to applications. Hepatology 2023. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zahmatkesh, E.; Ghanian, M.H.; Zarkesh, I.; Farzaneh, Z.; Halvaei, M.; Heydari, Z.; Moeinvaziri, F.; Othman, A.; Ruoß, M.; Piryaei, A.; et al. Tissue-specific microparticles improve organoid microenvironment for efficient maturation of pluripotent stem-cell-derived hepatocytes. Cells 2021, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Pettinato, G.; Lehoux, S.; Ramanathan, R.; Salem, M.M.; He, L.X.; Muse, O.; Flaumenhaft, R.; Thompson, M.T.; Rouse, E.A.; Cummings, R.D.; et al. Generation of fully functional hepatocyte-like organoids from human induced pluripotent stem cells mixed with Endothelial Cells. Sci. Rep. 2019, 9, 8920. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Ikemoto, T.; Morine, Y.; Imura, S.; Saito, Y.; Yamada, S.; Shimada, M. The differences in the characteristics of insulin-producing cells using human adipose-tissue derived mesenchymal stem cells from subcutaneous and visceral tissues. Sci. Rep. 2019, 9, 13204. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, K.; Ikemoto, T.; Yamashita, S.; Miyazaki, K.; Okikawa, S.; Yamada, S.; Saito, Y.; Morine, Y.; Shimada, M. Syngeneically transplanted insulin producing cells differentiated from adipose derived stem cells undergo delayed damage by autoimmune responses in NOD mice. Sci. Rep. 2022, 12, 5852. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.R.; Ueno, Y.; Zheng, Y.W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Yoshitomi, H.; Rossant, J.; Zaret, K.S. Liver organogenesis promoted by endothelial cells prior to vascular function. Science 2001, 294, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Li, W.H.; Ma, M.H.; Lu, X.; Zhang, H.Y. Endothelial cell-derived matrix promotes the metabolic functional maturation of hepatocyte via integrin-Src signaling. J. Cell Mol. Med. 2017, 21, 2809–2822. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.M.; Zhou, X.; Chen, C.Y.; Lv, D.D.; Huang, W.J.; Peng, Y.; Wu, H.P.; Chen, Y.; Tang, D.; Guo, L.N.; et al. Establishment of functional liver spheroids from human hepatocyte-derived liver progenitor-like cells for cell therapy. Front. Bioeng. Biotechnol. 2021, 9, 738081. [Google Scholar] [CrossRef] [PubMed]
- Raggi, C.; M’Callum, M.A.; Pham, Q.T.; Gaub, P.; Selleri, S.; Baratang, N.V.; Mangahas, C.L.; Cagnone, G.; Reversade, B.; Joyal, J.S.; et al. Leveraging interacting signaling pathways to robustly improve the quality and yield of human pluripotent stem cell-derived hepatoblasts and hepatocytes. Stem Cell Rep. 2022, 17, 584–598. [Google Scholar] [CrossRef]
- Li, J.; Xing, F.; Chen, F.; He, L.; So, K.F.; Liu, Y.; Xiao, J. Functional 3D human liver bud assembled from MSC-derived multiple liver cell lineages. Cell Transplant. 2019, 28, 510–521. [Google Scholar] [CrossRef]
- Weng, Y.; Han, S.; Sekyi, M.T.; Su, T.; Mattis, A.N.; Chang, T.T. Self-assembled Matrigel-free iPSC-derived liver organoids demonstrate wide-ranging highly differentiated liver functions. Stem Cells 2023, 41, 126–139. [Google Scholar] [CrossRef]
- Nakazato, T.; Kawamura, T.; Uemura, T.; Liu, L.; Li, J.; Sasai, M.; Harada, A.; Ito, E.; Iseoka, H.; Toda, K.; et al. Engineered three-dimensional cardiac tissues maturing in a rotating wall vessel bioreactor remodel diseased hearts in rats with myocardial infarction. Stem Cell Rep. 2022, 17, 1170–1182. [Google Scholar] [CrossRef]
- Bogliotti, Y.; Vander Roest, M.; Mattis, A.N.; Gish, R.G.; Peltz, G.; Anwyl, R.; Kivlighn, S.; Schuur, E.R. Clinical application of induced hepatocyte-like cells produced from mesenchymal stromal cells: A literature review. Cells 2022, 11, 1998. [Google Scholar] [CrossRef]
- Aurich, H.; Sgodda, M.; Kaltwaßer, P.; Vetter, M.; Weise, A.; Liehr, T.; Brulport, M.; Hengstler, J.G.; Dollinger, M.M.; Fleig, W.E.; et al. Hepatocyte differentiation of mesenchymal stem cells from human adipose tissue in vitro promotes hepatic integration in vivo. Gut 2009, 58, 570–581. [Google Scholar] [CrossRef]
- Bruno, S.; Herrera Sanchez, M.B.; Pasquino, C.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G. Human liver-derived stem cells improve fibrosis and inflammation associated with nonalcoholic steatohepatitis. Stem Cells Int. 2019, 2019, 6351091. [Google Scholar] [CrossRef] [PubMed]
- Afzali, B.; Lombardi, G.; Lechler, R.I. Pathways of Major Histocompatibility Complex Allorecognition. Curr. Opin. Organ Transplant. 2008, 13, 438–444. [Google Scholar] [CrossRef]
- Romano, M.; Elgueta, R.; McCluskey, D.; Ortega-Prieto, A.M.; Stolarczyk, E.; Dazzi, F.; Lucendo-Villarin, B.; Meseguer-Ripolles, J.; Williams, J.; Fanelli, G.; et al. Pluripotent stem cell-derived hepatocytes inhibit T cell proliferation in vitro through tryptophan starvation. Cells 2021, 11, 24. [Google Scholar] [CrossRef]
- Huang, X.P.; Sun, Z.; Miyagi, Y.; McDonald Kinkaid, H.; Zhang, L.; Weisel, R.D.; Li, R.K. Differentiation of allogeneic mesenchymal stem cells induces immunogenicity and limits their long-term benefits for myocardial repair. Circulation 2010, 122, 2419–2429. [Google Scholar] [CrossRef]
- Technau, A.; Froelich, K.; Hagen, R.; Kleinsasser, N. Adipose tissuederived stem cells show both immunogenic and immunosuppressive properties after chondrogenic differentiation. Cytotherapy 2011, 13, 310–317. [Google Scholar] [CrossRef]
- Liu, H.; Kemeny, D.M.; Heng, B.C.; Ouyang, H.W.; Melendez, A.J.; Cao, T. The immunogenicity and immunomodulatory function of osteogenic cells differentiated from mesenchymal stem cells. J. Immunol. 2006, 176, 2864–2871. [Google Scholar] [CrossRef] [PubMed]
- Tee, J.Y.; Vaghjiani, V.; Liu, Y.H.; Murthi, P.; Chan, J.; Manuelpillai, U. Immunogenicity and immunomodulatory properties of hepatocyte-like cells derived from human amniotic epithelial cells. Curr. Stem Cell Res. Ther. 2013, 8, 91–99. [Google Scholar] [PubMed]
- Zhu, W.; Li, M.; Zou, J.; Zhang, D.; Fang, M.; Sun, Y.; Li, C.; Tang, M.; Wang, Y.; Zhou, Q.; et al. Induction of local immunosuppression in allogeneic cell transplantation by cell-type-specific expression of PD-L1 and CTLA4Ig. Stem Cell Rep. 2023, 18, 2344–2355. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Saito, Y.; Waki, Y.; Ikemoto, T.; Teraoku, H.; Yamada, S.; Morine, Y.; Shimada, M. Generation of Highly Functional Hepatocyte-like Organoids from Human Adipose-Derived Mesenchymal Stem Cells Cultured with Endothelial Cells. Cells 2024, 13, 547. https://doi.org/10.3390/cells13060547
Chen S, Saito Y, Waki Y, Ikemoto T, Teraoku H, Yamada S, Morine Y, Shimada M. Generation of Highly Functional Hepatocyte-like Organoids from Human Adipose-Derived Mesenchymal Stem Cells Cultured with Endothelial Cells. Cells. 2024; 13(6):547. https://doi.org/10.3390/cells13060547
Chicago/Turabian StyleChen, Shuhai, Yu Saito, Yuhei Waki, Tetsuya Ikemoto, Hiroki Teraoku, Shinichiro Yamada, Yuji Morine, and Mitsuo Shimada. 2024. "Generation of Highly Functional Hepatocyte-like Organoids from Human Adipose-Derived Mesenchymal Stem Cells Cultured with Endothelial Cells" Cells 13, no. 6: 547. https://doi.org/10.3390/cells13060547