

Granular Insights: Neutrophil Predominance and Elastase Release in Severe Asthma Exacerbations in a Pediatric Cohort
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Data Collection
2.3. Blood Collection and Human Neutrophil and Eosinophil Purification
2.4. Cell Surface Expression of Neutrophil and Eosinophil Markers of Activation
2.5. Ex Vivo Cytokine Production and Elastase Release
2.6. Statistical Analysis
3. Results
3.1. Enrollment
3.2. Neutrophilia in Children with Severe Asthma Exacerbations
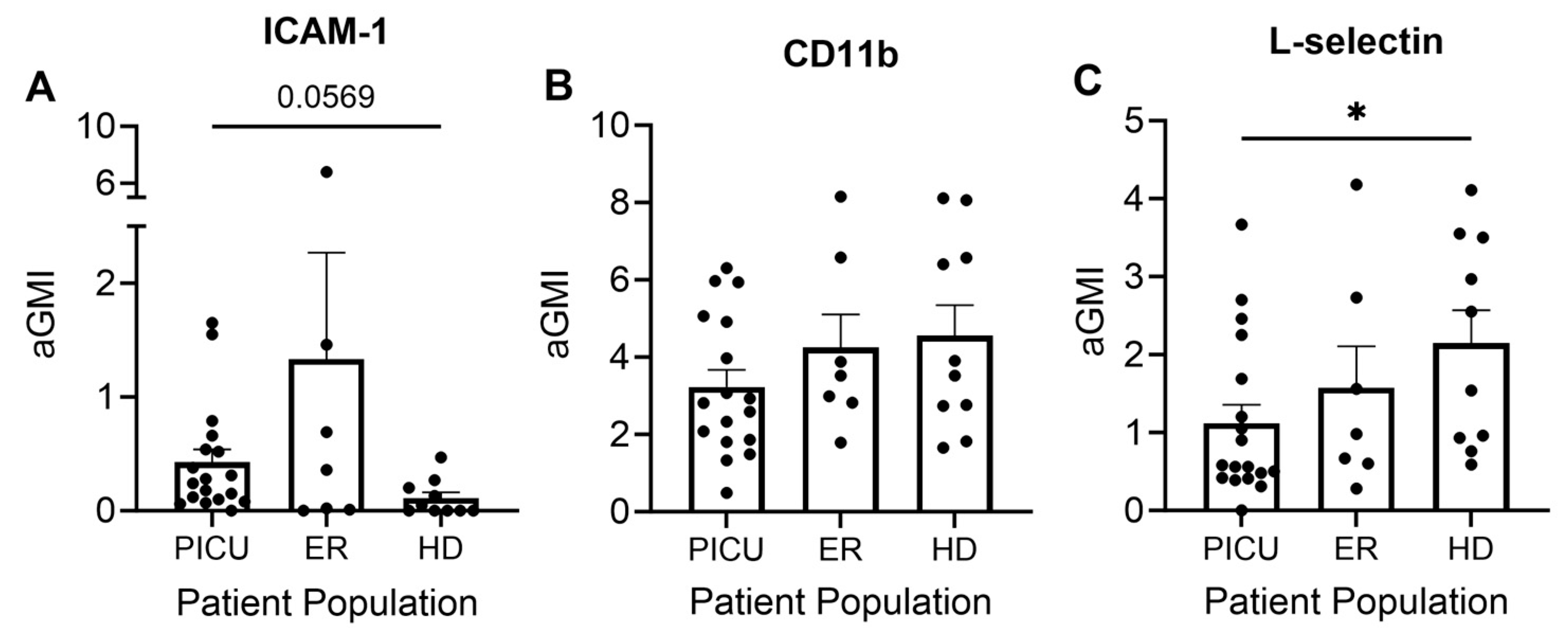
3.3. Neutrophil and Eosinophil Inflammatory Cell Surface Markers
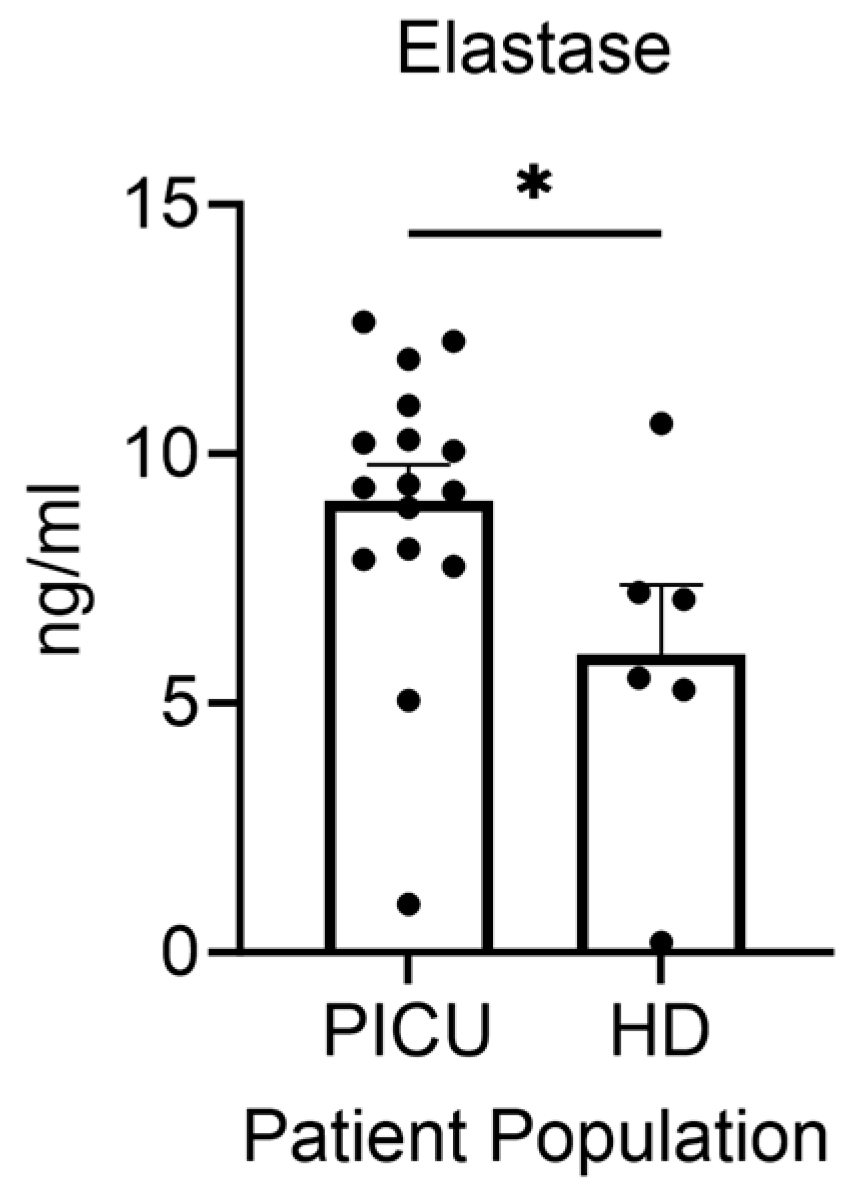
3.4. Elastase Secretion by Neutrophils during Asthma Exacerbation
3.5. Cytokine Secretion by Neutrophils
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bacharier, L.B.; Boner, A.; Carlsen, K.H.; Eigenmann, P.A.; Frischer, T.; Gotz, M.; Helms, P.J.; Hunt, J.; Liu, A.; Papadopoulos, N.; et al. Diagnosis and treatment of asthma in childhood: A PRACTALL consensus report. Allergy 2008, 63, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Sly, R.M. New guidelines for diagnosis and management of asthma. Ann. Allergy Asthma Immunol. 1997, 78, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef] [PubMed]
- National Asthma, E.; Prevention, P. Expert Panel Report 3 (EPR-3): Guidelines for the Diagnosis and Management of Asthma-Summary Report 2007. J. Allergy Clin. Immunol. 2007, 120, S94–S138. [Google Scholar] [CrossRef]
- Dolan, C.M.; Fraher, K.E.; Bleecker, E.R.; Borish, L.; Chipps, B.; Hayden, M.L.; Weiss, S.; Zheng, B.; Johnson, C.; Wenzel, S.; et al. Design and baseline characteristics of the epidemiology and natural history of asthma: Outcomes and Treatment Regimens (TENOR) study: A large cohort of patients with severe or difficult-to-treat asthma. Ann. Allergy Asthma Immunol. 2004, 92, 32–39. [Google Scholar] [CrossRef]
- Nievas, I.F.; Anand, K.J. Severe acute asthma exacerbation in children: A stepwise approach for escalating therapy in a pediatric intensive care unit. J. Pediatr. Pharmacol. Ther. 2013, 18, 88–104. [Google Scholar] [CrossRef]
- Kochanek, K.D.; Murphy, S.L.; Xu, J.; Tejada-Vera, B. Deaths: Final Data for 2014. Natl. Vital. Stat. Rep. 2016, 65, 1–122. [Google Scholar]
- Chipps, B.E.; Zeiger, R.S.; Borish, L.; Wenzel, S.E.; Yegin, A.; Hayden, M.L.; Miller, D.P.; Bleecker, E.R.; Simons, F.E.; Szefler, S.J.; et al. Key findings and clinical implications from The Epidemiology and Natural History of Asthma: Outcomes and Treatment Regimens (TENOR) study. J. Allergy Clin. Immunol. 2012, 130, 332–342. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Kotsimbos, A.T.; Hamid, Q. IL-5 and IL-5 receptor in asthma. Mem. Inst. Oswaldo Cruz 1997, 92 (Suppl. 2), 75–91. [Google Scholar] [CrossRef]
- Nakagome, K.; Matsushita, S.; Nagata, M. Neutrophilic inflammation in severe asthma. Int. Arch. Allergy Immunol. 2012, 158 (Suppl. 1), 96–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Jarjour, N.N.; Busse, W.W.; Kelly, E.A. Chemokine receptor expression on human eosinophils from peripheral blood and bronchoalveolar lavage fluid after segmental antigen challenge. J. Allergy Clin. Immunol. 2003, 112, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Poulakos, M.N.; Cargill, S.M.; Waineo, M.F.; Wolford, A.L., Jr. Mepolizumab for the treatment of severe eosinophilic asthma. Am. J. Health Syst. Pharm. 2017, 74, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, C.L.; Shaughnessy, T.E.; Matthay, M.A.; Fahy, J.V. Increased neutrophil numbers and IL-8 levels in airway secretions in acute severe asthma: Clinical and biologic significance. Am. J. Respir. Crit. Care Med. 2000, 161, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, F.P.; Wang, G.; Mao, H. Role of Neutrophil Extracellular Traps in Asthma and Chronic Obstructive Pulmonary Disease. Chin. Med. J. 2017, 130, 730–736. [Google Scholar] [CrossRef]
- Pease, J.E. Asthma, allergy and chemokines. Curr. Drug Targets 2006, 7, 3–12. [Google Scholar] [CrossRef]
- Nauseef, W.M. Isolation of human neutrophils from venous blood. Methods Mol. Biol. 2014, 1124, 13–18. [Google Scholar] [CrossRef]
- Luder, E.; Melnik, T.A.; DiMaio, M. Association of being overweight with greater asthma symptoms in inner city black and Hispanic children. J. Pediatr. 1998, 132, 699–703. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, L.; Lizarraga, R.; Chen, Y. Human Airway Epithelial Cells Direct Significant Rhinovirus Replication in Monocytic Cells by Enhancing ICAM1 Expression. Am. J. Respir. Cell Mol. Biol. 2017, 57, 216–225. [Google Scholar] [CrossRef]
- Bui, T.M.; Wiesolek, H.L.; Sumagin, R. ICAM-1: A master regulator of cellular responses in inflammation, injury resolution, and tumorigenesis. J. Leukoc. Biol. 2020, 108, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Noguera, A.; Batle, S.; Miralles, C.; Iglesias, J.; Busquets, X.; MacNee, W.; Agusti, A.G. Enhanced neutrophil response in chronic obstructive pulmonary disease. Thorax 2001, 56, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling. Front. Immunol. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Hassani, M.; van Staveren, S.; van Grinsven, E.; Bartels, M.; Tesselaar, K.; Leijte, G.; Kox, M.; Pickkers, P.; Vrisekoop, N.; Koenderman, L. Characterization of the phenotype of human eosinophils and their progenitors in the bone marrow of healthy individuals. Haematologica 2020, 105, e52–e56. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Alkhouri, H.; Tang, F.; Che, W.; Ge, Q.; Ammit, A.J. Sphingosine 1-phosphate induces neutrophil chemoattractant IL-8: Repression by steroids. PLoS ONE 2014, 9, e92466. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Ying, S.; Dai, Y. Pathological Roles of Neutrophil-Mediated Inflammation in Asthma and Its Potential for Therapy as a Target. J. Immunol. Res. 2017, 2017, 3743048. [Google Scholar] [CrossRef]
- Possa, S.S.; Leick, E.A.; Prado, C.M.; Martins, M.A.; Tiberio, I.F. Eosinophilic inflammation in allergic asthma. Front. Pharmacol. 2013, 4, 46. [Google Scholar] [CrossRef]
- Mannino, D.M.; Homa, D.M.; Akinbami, L.J.; Moorman, J.E.; Gwynn, C.; Redd, S.C. Surveillance for asthma—United States, 1980–1999. MMWR Surveill. Summ. 2002, 51, 1–13. [Google Scholar]
- Polverino, E.; Rosales-Mayor, E.; Dale, G.E.; Dembowsky, K.; Torres, A. The Role of Neutrophil Elastase Inhibitors in Lung Diseases. Chest 2017, 152, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Doring, G. The role of neutrophil elastase in chronic inflammation. Am. J. Respir. Crit. Care Med. 1994, 150, S114–S117. [Google Scholar] [CrossRef]
- Poto, R.; Shamji, M.; Marone, G.; Durham, S.R.; Scadding, G.W.; Varricchi, G. Neutrophil Extracellular Traps in Asthma: Friends or Foes? Cells 2022, 11, 3521. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, M.; Jackson, D.J.; Swieboda, D.; Guedan, A.; Tsourouktsoglou, T.D.; Ching, Y.M.; Radermecker, C.; Makrinioti, H.; Aniscenko, J.; Bartlett, N.W.; et al. Host DNA released by NETosis promotes rhinovirus-induced type-2 allergic asthma exacerbation. Nat. Med. 2017, 23, 681–691. [Google Scholar] [CrossRef]
- Tcherniuk, S.; Cenac, N.; Comte, M.; Frouard, J.; Errazuriz-Cerda, E.; Galabov, A.; Morange, P.E.; Vergnolle, N.; Si-Tahar, M.; Alessi, M.C.; et al. Formyl Peptide Receptor 2 Plays a Deleterious Role During Influenza A Virus Infections. J. Infect. Dis. 2016, 214, 237–247. [Google Scholar] [CrossRef]
- Dworski, R.; Simon, H.U.; Hoskins, A.; Yousefi, S. Eosinophil and neutrophil extracellular DNA traps in human allergic asthmatic airways. J. Allergy Clin. Immunol. 2011, 127, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Gál, Z.; Gézsi, A.; Pállinger, É.; Visnovitz, T.; Nagy, A.; Kiss, A.; Sultész, M.; Csoma, Z.; Tamási, L.; Gálffy, G.; et al. Plasma neutrophil extracellular trap level is modified by disease severity and inhaled corticosteroids in chronic inflammatory lung diseases. Sci. Rep. 2020, 10, 4320. [Google Scholar] [CrossRef]
- Grunwell, J.R.; Stephenson, S.T.; Tirouvanziam, R.; Brown, L.A.S.; Brown, M.R.; Fitzpatrick, A.M. Children with Neutrophil-Predominant Severe Asthma Have Proinflammatory Neutrophils With Enhanced Survival and Impaired Clearance. J. Allergy Clin. Immunol. Pract. 2019, 7, 516–525. [Google Scholar] [CrossRef]
- Xu, S.; Chen, Z.; Ge, L.; Ma, C.; He, Q.; Liu, W.; Zhang, L.; Zhou, L. Identification of potential biomarkers and pathogenesis in neutrophil-predominant severe asthma: A comprehensive bioinformatics analysis. Medicine 2022, 101, e30661. [Google Scholar] [CrossRef]
- Hafezi-Moghadam, A.; Thomas, K.L.; Prorock, A.J.; Huo, Y.; Ley, K. L-selectin shedding regulates leukocyte recruitment. J. Exp. Med. 2001, 193, 863–872. [Google Scholar] [CrossRef]
- Shukla, S.D.; Shastri, M.D.; Vanka, S.K.; Jha, N.K.; Dureja, H.; Gupta, G.; Chellappan, D.K.; Oliver, B.G.; Dua, K.; Walters, E.H. Targeting intercellular adhesion molecule-1 (ICAM-1) to reduce rhinovirus-induced acute exacerbations in chronic respiratory diseases. Inflammopharmacology 2022, 30, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.; Llanos, J.P.; Lafeuille, M.H.; Duh, M.S.; Germain, G.; Lejeune, D.; Sama, S.; Bell, C.; Hahn, B. Effects of systemic corticosteroids on blood eosinophil counts in asthma: Real-world data. J. Asthma 2019, 56, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.J.; Bacharier, L.B.; Calatroni, A.; Gill, M.A.; Hu, J.; Liu, A.H.; Wheatley, L.M.; Gern, J.E.; Gruchalla, R.S.; Khurana Hershey, G.K.; et al. Serum IL-6: A biomarker in childhood asthma? J. Allergy Clin. Immunol. 2020, 145, 1701–1704. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.A. The role of dendritic cells in asthma. J. Allergy Clin. Immunol. 2012, 129, 889–901. [Google Scholar] [CrossRef]
- Wei, Q.; Liao, J.; Jiang, M.; Liu, J.; Liang, X.; Nong, G. Relationship between Th17-mediated immunity and airway inflammation in childhood neutrophilic asthma. Allergy Asthma Clin. Immunol. 2021, 17, 4. [Google Scholar] [CrossRef]
- Nichols, B.E.; Hook, J.S.; Weng, K.; Ahn, C.; Moreland, J.G. Novel neutrophil phenotypic signature in pediatric patients with type 1 diabetes and diabetic ketoacidosis. J. Leukoc. Biol. 2022, 111, 849–856. [Google Scholar] [CrossRef]
- Forno, E.; Ortega, V.E.; Celedon, J.C. Asthma and Chronic Obstructive Pulmonary Disease. Clin. Chest Med. 2023, 44, 519–530. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | PICU Median (IQR) N = 23 | ER Median (IQR) N = 9 | Healthy Donors Median (IQR) N = 10 |
|---|---|---|---|
| Age (yrs) | 9 (6–13) | 11 (6–13) | 31.5 (15–39.75) |
| Sex (% male) | 56.5% | 78% | 40% |
| Ethnic minority (%) | 82.5% | 100% | 40% |
| Black, non-Hispanic | 65% | 78% | 10% |
| White, non-Hispanic | 17.5% | 0% | 60% |
| Hispanic | 17.5% | 22% | 10% |
| Asian | - | - | 20% |
| Severity of Asthma | |||
| Mild persistent | 13% | 33% | |
| Moderate persistent | 52% | 22% | |
| Severe persistent | 35% | 44% | |
| Rhino/Entero + | 67% | - | |
| Comorbidities: | |||
| Allergic Rhinitis | 87% | 78% | |
| Eczema | 26% | 44% | |
| BMI (percentile) | 85th (60–95) n = 19 | 42nd (21–67) n = 4 | |
| IgE (IU/mL) | 948 (264–2080) n = 11 | 954 (434–2136) n = 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henley, K.; Tresselt, E.; Hook, J.S.; Patel, P.A.; Gill, M.A.; Moreland, J.G. Granular Insights: Neutrophil Predominance and Elastase Release in Severe Asthma Exacerbations in a Pediatric Cohort. Cells 2024, 13, 533. https://doi.org/10.3390/cells13060533
Henley K, Tresselt E, Hook JS, Patel PA, Gill MA, Moreland JG. Granular Insights: Neutrophil Predominance and Elastase Release in Severe Asthma Exacerbations in a Pediatric Cohort. Cells. 2024; 13(6):533. https://doi.org/10.3390/cells13060533
Chicago/Turabian StyleHenley, Kirstin, Erin Tresselt, Jessica S. Hook, Parth A. Patel, Michelle A. Gill, and Jessica G. Moreland. 2024. "Granular Insights: Neutrophil Predominance and Elastase Release in Severe Asthma Exacerbations in a Pediatric Cohort" Cells 13, no. 6: 533. https://doi.org/10.3390/cells13060533