
Toward the Analysis of Volatile Organic Compounds from Tomato Plants (Solanum lycopersicum L.) Treated with Trichoderma virens or/and Botrytis cinerea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Plant Material Cultivation, Inoculation with B. cinerea, and the Disease Development Determination
- (i)
- Plants grown in the soil without T. virens TRS 106 spores (control, C);
- (ii)
- Plants grown in the soil with T. virens TRS 106 spores (TRS 106);
- (iii)
- Plants grown in the soil without T. virens TRS 106 spores, and inoculated with B. cinerea (Bc);
- (iv)
- Plants grown in the soil with T. virens TRS 106 spores, and inoculated with B. cinerea (TRS 106 + Bc).
2.3. Assay of VOC Emission
2.4. Statistical Analyses
3. Results
3.1. T. virens TRS 106 Reduces Gray Mold Disease Index in Tomato Plants
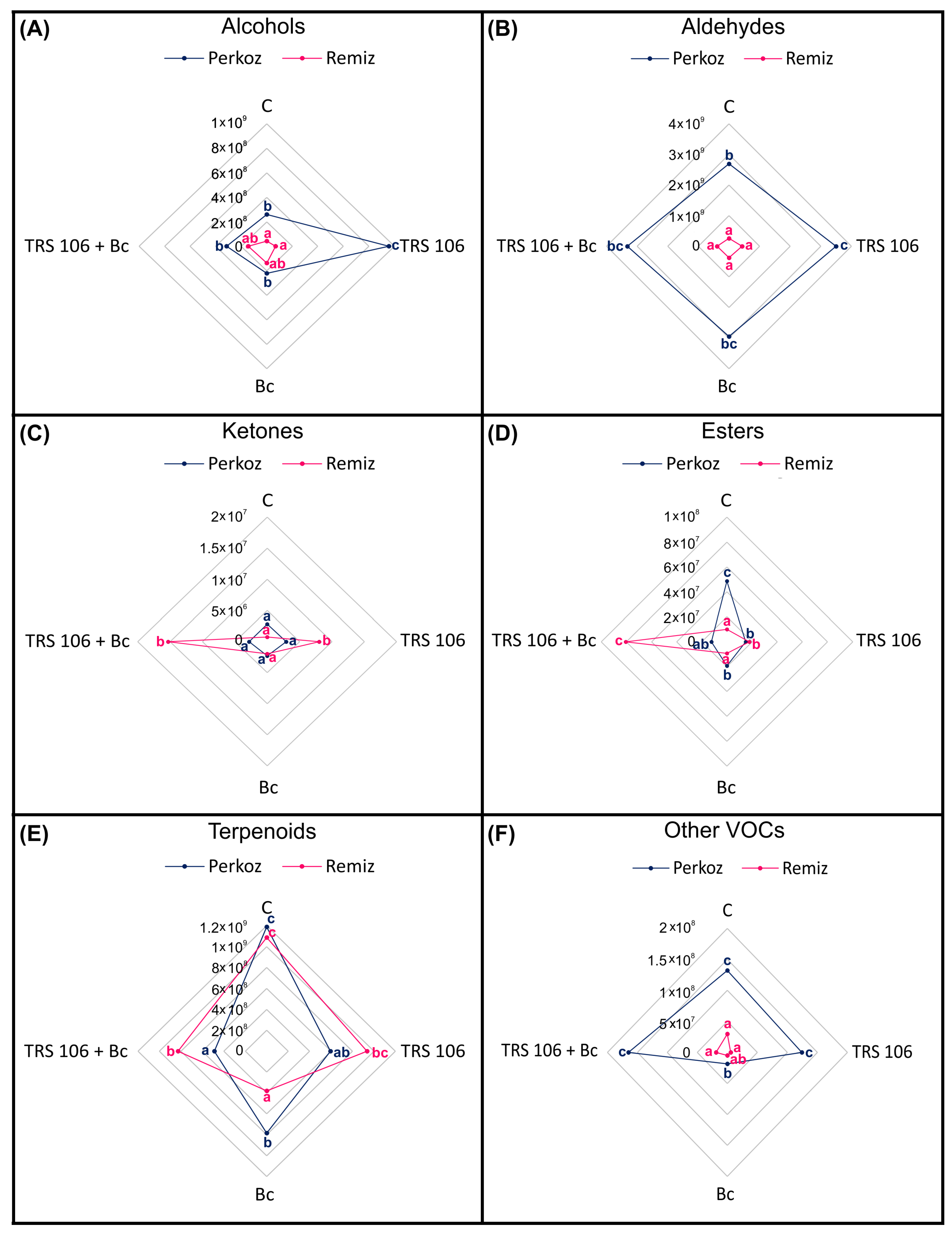
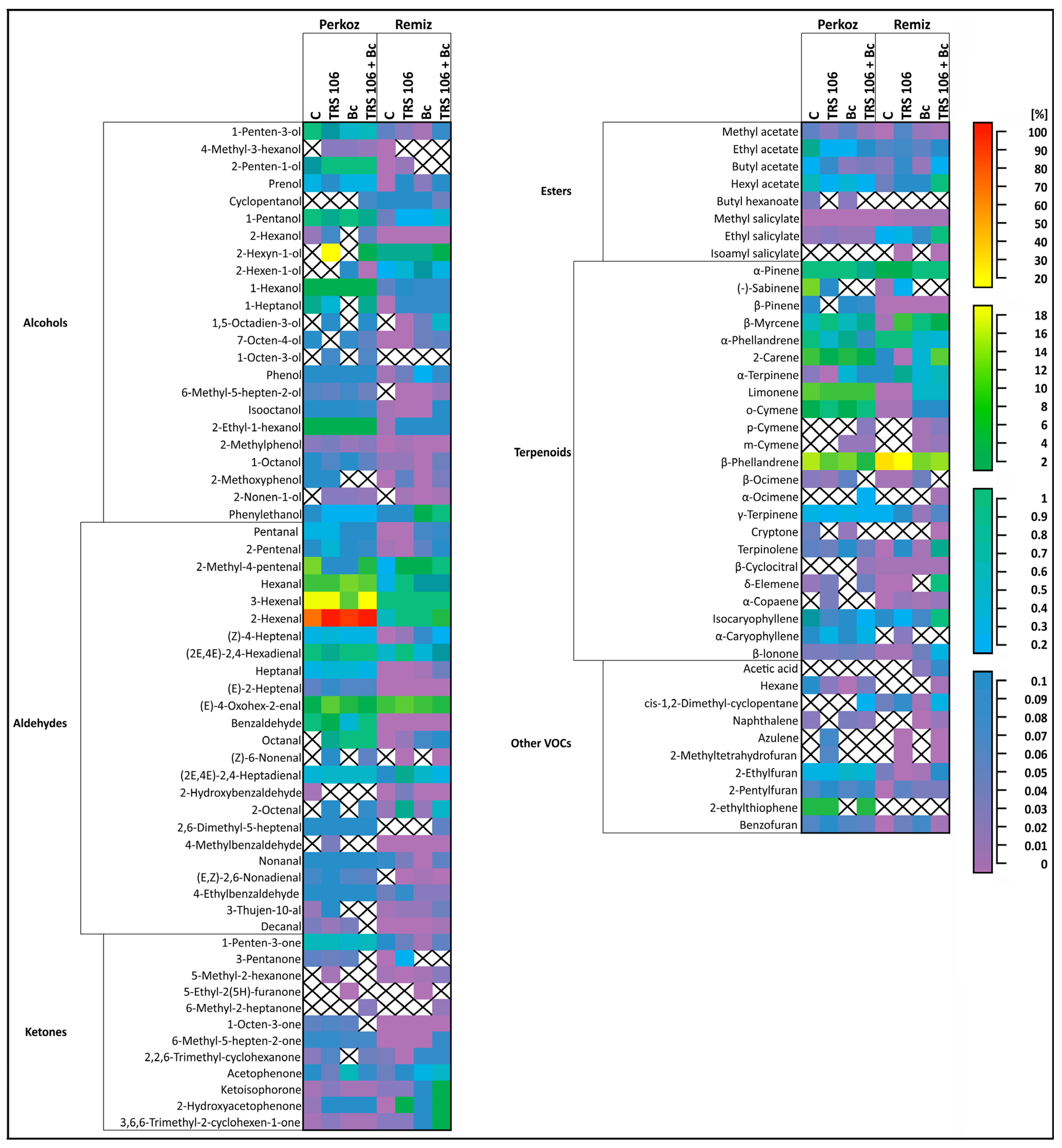
3.2. Tomato Plants Belonging to Perkoz and Remiz Demonstrate Different Emission of VOCs
3.3. VOCs in Tomato Plants, Belonging to the Perkoz and Remiz Varieties, and Infected with B. cinerea
3.4. VOCs in Tomato Plants Treated with T. virens TRS 106 and Uninoculated with B. cinerea
3.5. VOCs in Tomato Plants Treated with T. virens TRS 106 and Inoculated with B. cinerea
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maffei, M.E. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef] [PubMed]
- Viswanath, K.K.; Varakumar, P.; Pamuru, R.R.; Basha, S.J.; Mehta, S.; Rao, A.D. Plant lipoxygenases and their role in plant physiology. J. Plant Biol. 2020, 63, 83–95. [Google Scholar] [CrossRef]
- Chatzivasileiou, A.O.; Ward, V.; Edgar, S.M.; Stephanopoulos, G. Two-step pathway for isoprenoid synthesis. Proc. Natl. Acad. Sci. USA 2019, 116, 506–511. [Google Scholar] [CrossRef]
- Dhandapani, S.; Tjhang, J.G.; Jang, I.C. Production of multiple terpenes of different chain lengths by subcellular targeting of multi-substrate terpene synthase in plants. Metab. Eng. 2020, 61, 397–405. [Google Scholar] [CrossRef]
- Erb, M.; Veyrat, N.; Robert, C.A.M.; Xu, H.; Frey, M.; Ton, J.; Turlings, T.C.J. Indole is an essential herbivore-induced volatile priming signal in maize. Nat. Commun. 2015, 6, 6273. [Google Scholar] [CrossRef] [PubMed]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant. Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernan-dezMontiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef]
- Lazazzara, V.; Avesani, S.; Robatscher, P.; Oberhuber, M.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Biogenic volatile organic compounds in the grapevine response to pathogens, beneficial microorganisms, resistance inducers, and abiotic factors. J. Exp. Bot. 2021, 73, 529–554. [Google Scholar] [CrossRef]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial volatile organic compounds in intra-kingdom and inter-kingdom interactions. Nat. Rev. Microbiol. 2021, 19, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Beneficial effects of microbial volatile organic compounds (MVOCs) in plants. Appl. Soil. Ecol. 2021, 168, 104118. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Schmidt, R.; Etalo, D.W.; De Jager, V.; Gerards, S.; Zweers, H.; De Boer, W.; Garbeva, P. Microbial small talk: Volatiles in fungal–bacterial interactions. Front. Microbiol. 2016, 6, 1495. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tong, Z.; Zhang, X.; Wang, Y.; Fang, W.; Li, L.; Luo, Z. Unveiling the mechanisms for the plant volatile organic compound linalool to control gray mold on strawberry fruits. J. Agric. Food Chem. 2019, 67, 9265–9276. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Rodriguez, E.; Rivera-Macias, L.E.; Adame-Alvarez, R.M.; Torres, J.M.; Heil, M. Shared weapons in fungus-fungus and fungus-plant interactions? Volatile organic compounds of plant or fungal origin exert direct antifungal activity in vitro. Fungal Ecol. 2018, 33, 115–121. [Google Scholar] [CrossRef]
- Pertot, I.; Giovannini, O.; Benanchi, M.; Caffi, T.; Rossi, V.; Mugnai, L. Combining biocontrol agents with different mechanisms of action in a strategy to control Botrytis cinerea on grapevine. Crop. Prot. 2017, 97, 85–93. [Google Scholar] [CrossRef]
- Vlot, A.C.; Sales, J.H.; Lenk, H.M.; Bauer, K.; Brambilla, A.; Sommer, A.; Chen, Y.; Wenig, M.; Nayem, S. Systemic propagation of immunity in plants. New. Phytol. 2021, 229, 1234–1250. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant. Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Pinedo, C.; Moraga, J.; Collado, I.G.; Aleu, J. Botrytis species as biocatalysts. Phytochem. Rev. 2020, 19, 529–558. [Google Scholar] [CrossRef]
- You, J.; Zhang, J.; Wua, M.; Yang, L.; Chen, W.; Li, G. Multiple criteria-based screening of Trichoderma isolates for biological control of Botrytis cinerea on tomato. Biol. Control 2016, 101, 31–38. [Google Scholar] [CrossRef]
- Risoli, S.; Cotrozzi, L.; Sarrocco, S.; Nuzzaci, M.; Pellegrini, E.; Vitti, A. Trichoderma-induced resistance to Botrytis cinerea in Solanum species: A Meta-Analysis. Plants 2022, 11, 180. [Google Scholar] [CrossRef] [PubMed]
- Vos, C.M.F.; De Cremer, K.; Cammue, B.P.A.; De Coninck, B. The toolbox of Trichoderma spp. in the biocontrol of Botrytis cinerea disease. Mol. Plant. Pathol. 2015, 16, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Barquero, M.; González-Andrés, F. Insight into the microbiological control strategies against Botrytis cinerea using systemic plant resistance activation. Agronomy 2020, 10, 1822. [Google Scholar] [CrossRef]
- Gomes, E.V.; Ulhoa, C.J.; Cardoza, R.E.; Silva, R.N.; Gutiérrez, S. Involvement of Trichoderma harzianum Epl-1 protein in the regulation of Botrytis virulence- and tomato defense-related genes. Front. Plant Sci. 2017, 8, 880. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Siwulski, M.; Sobieralski, K.; Lisiecka, J.; Jędryczka, M. Trichoderma spp.–application and prospects for use in organic farming and industry. J. Plant Prot. Res. 2014, 54, 309–317. [Google Scholar] [CrossRef]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma as biological control agent: Scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U. Diversity in plant systemic resistance induced by Trichoderma. Biol. Control 2013, 67, 149–156. [Google Scholar] [CrossRef]
- Ghazanfar, M.U.; Raza, M.; Raza, W.; Qamar, M.I. Trichoderma as potential biocontrol agent, its exploitation in agriculture: A review. Plant Prot. 2018, 2, 109–135. [Google Scholar]
- Gupta, R.; Bar, M. Plant immunity, priming, and systemic resistance as mechanisms for Trichoderma spp. biocontrol. In Trichoderma; Sharma, A., Sharma, P., Eds.; Springer: Singapore, 2020; pp. 81–110. [Google Scholar]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Leroux, P. Chemical control of Botrytis and its resistance to chemical fungicides. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 195–222. [Google Scholar]
- Tapwal, A.; Singh, U.; Teixeira, J.A.; Singh, G.; Garg, S.; Kumar, R. In vitro antagonism of Trichoderma viride against five phytopathogens. Pest Technol. 2011, 5, 59–62. [Google Scholar]
- Marín-Chacón, M.A.; Rivera-Coto, G.; Villalobos-Moya, K.; Orozco-Rodríguez, R.; Orozco-Cayasso, S. Evaluation of Botrytis cinerea Pers., antagonist fungi in blackberry plantations of Costa Rica. Agron. Costarric. 2017, 41, 7–18. [Google Scholar] [CrossRef]
- Roca-Couso, R.; Flores-Félix, J.D.; Rivas, R. Mechanisms of action of microbial biocontrol agents against Botrytis cinerea. J. Fungi 2021, 7, 1045. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Herrera-Estrella, A.; Lόpez-Bucio, J. The 4-phosphopantetheinyl transferase of Trichoderma virens plays a role in plant protection against Botrytis cinerea through volatile organic compound emission. Plant Soil 2014, 379, 261–274. [Google Scholar] [CrossRef]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Uresti-Rivera, E.E.; Castro-Longoria, E.; Herrera-Estrella, A.; Casas-Flores, S. Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acid/ethylene and salicylic acid pathways. Eur. J. Plant Pathol. 2011, 131, 15–26. [Google Scholar] [CrossRef]
- Salas-Marina, M.A.; Isordia-Jasso, M.I.; Islas-Osuna, M.A.; Delgado-Sã¡nchez, P.; Bremont, J.F.J.; Rodríguez-Kessler, M.; Rosales-Saavedra, M.T.; Eherrera-Estrella, A.; Casas-Flores, S. The Epl1 and Sm1 proteins from Trichoderma atroviride and Trichoderma virens differentially modulate systemic disease resistance against different life style pathogens in Solanum lycopersicum. Front. Plant Sci. 2015, 6, 77. [Google Scholar] [CrossRef]
- Xie, Y.; Tian, L.; Han, X.; Yang, Y. Research advances in allelopathy of volatile organic compounds (VOCs) of plants. Horticulturae 2021, 7, 278. [Google Scholar] [CrossRef]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mold (Botrytis cinerea)–prospects and challenges. Biocontrol Sci. Technol. 2019, 29, 207–228. [Google Scholar] [CrossRef]
- Małolepsza, U.; Nawrocka, J.; Szczech, M. Trichoderma virens 106 inoculation stimulates defence enzyme activities and enhances phenolic levels in tomato plants leading to lowered Rhizoctonia solani infection. Biocontrol Sci. Technol. 2017, 27, 180–199. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szczech, M. Involvement of metabolic components, volatile compounds, PR proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef]
- Oskiera, M.; Szczech, M.; Bartoszewski, G. Molecular identification of Trichoderma strains collected to develop plant growth-promoting and biocontrol agents. J. Hort. Res. 2015, 23, 75–86. [Google Scholar] [CrossRef]
- Nawrocka, J.; Szymczak, K.; Maćkowiak, A.; Skwarek-Fadecka, M.; Małolepsza, U. Determination of Reactive Oxygen or Nitrogen Species and novel Volatile Organic Compounds in the defense responses of tomato plants against Botrytis cinerea induced by Trichoderma virens TRS 106. Cells 2022, 11, 3051. [Google Scholar] [CrossRef]
- Taheri, P.; Tarighi, S. Riboflavin induces resistance in rice against Rhizoctonia solani via jasmonate-mediated priming of phenylpropanoid pathway. J. Plant Physiol. 2010, 167, 201–208. [Google Scholar] [CrossRef]
- Carlin, S.; Vrhovsek, U.; Franceschi, P.; Lotti, C.; Bontempo, L.; Camin, F.; Toubiana, D.; Zottele, F.; Toller, G.; Fait, A.; et al. Regional features of northern Italian sparkling wines, identified using solidphase micro extraction and comprehensive two-dimensional gas chromatography coupled with time-of-flight mass spectrometry. Food Chem. 2016, 208, 68–80. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Lisón, P.; Campos, L.; Rodrigo, I.; Rambla, J.L.; Granell, A.; Conejero, V.; Bellés, J.M. A non-targeted metabolomics approach unravels the VOCs associated with the tomato immune response against Pseudomonas syringae. Front. Plant Sci. 2017, 8, 1188. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Souza, M.A.; Peres, L.E.; Freschi, J.R.; Purgatto, E.; Lajolo, F.M.; Hassimotto, N.M.A. Changes in flavonoid and carotenoid profiles alter volatile organic compounds in purple and orange cherry tomatoes obtained by allele introgression. J. Sci. Food Agric. 2020, 100, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Kreissl, J.; Schieberle, P. Characterization of aroma-active compounds in Italian tomatoes with emphasis on new odorants. J. Sci. Food Agric. 2017, 65, 5198–5208. [Google Scholar] [CrossRef]
- Lim, T.K. Edible Medicinal and Non-Medicinal Plants; Springer: Dordrecht, The Netherlands, 2012; Volume 6, pp. 389–394. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Chang, P.; Shen, Y.; Wu, L.; El-Sappah, A.H.; Zhang, F.; Liang, Y. Comparing the flavor characteristics of 71 tomato (Solanum lycopersicum) accessions in Central Shaanxi. Front. Plant Sci. 2020, 11, 586834. [Google Scholar] [CrossRef]
- Clancy, M.V.; Haberer, G.; Jud, W.; Niederbacher, B.; Niederbacher, S.; Senft, M.; Zytynska, S.E.; Weisser, W.W.; Schnitzler, J.P. Under fire-simultaneous volatilome and transcriptome analysis unravels fine-scale responses of tansy chemotypes to dual herbivore attack. BMC Plant Biol. 2020, 20, 551. [Google Scholar] [CrossRef]
- Ricciardi, V.; Marcianò, D.; Sargolzaei, M.; Maddalena, G.; Maghradze, D.; Tirelli, A.; Casati, P.; Bianco, P.A.; Failla, O.; Fracassetti, D.; et al. From plant resistance response to the discovery of antimicrobial compounds: The role of volatile organic compounds (VOCs) in grapevine downy mildew infection. Plant Physiol. Biochem. 2021, 160, 294–305. [Google Scholar] [CrossRef]
- Harman, G.E.; Herrera-Estrella, A.H.; Horwitz, B.A.; Lorito, M. Trichoderma–from basic biology to biotechnology. Microbiology 2012, 158, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Yang, C.A.; Peng, K.C. Antagonism of Trichoderma harzianum ETS 323 on Botrytis cinerea mycelium in culture conditions. Phytopathology 2012, 102, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Mathys, J.; De Cremer, K.; Timmermans, P.; Van Kerckhove, S.; Lievens, B.; Vanhaecke, M.; Cammue, B.P.; De Coninck, B. Genome-Wide Characterization of ISR Induced in Arabidopsis thaliana by Trichoderma hamatum T382 Against Botrytis cinerea Infection. Front. Plant Sci. 2012, 3, 108. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Mulla, S.I.; Lee, K.-J.; Chae, J.-C.; Shukla, P. VOCs-mediated hormonal signaling and crosstalk with plant growth promoting microbes. Crit. Rev. Biotechnol. 2018, 38, 1277–1296. [Google Scholar] [CrossRef]
- Battaglia, D.; Bossi, S.; Cascone, P.; Digilio, M.C.; Prieto, J.D.; Fanti, P.; Guerrieri, E.; Iodice, L.; Lingua, G.; Lorito, M.; et al. Tomato below ground–above ground interactions: Trichoderma longibrachiatum affects the performance of Macrosiphum euphorbiae and its natural antagonists. Mol. Plant-Microbe 2013, 26, 1249–1256. [Google Scholar] [CrossRef]
- Dini, I.; Marra, R.; Cavallo, P.; Pironti, A.; Sepe, I.; Troisi, J.; Scala, G.; Lombari, P.; Vinale, F. Trichoderma strains and metabolites selectively increase the production of volatile organic compounds (VOCs) in olive trees. Metabolites 2021, 11, 213. [Google Scholar] [CrossRef]
- Martina, M.; Tikunov, Y.; Portis, E.; Bovy, A.G. The genetic basis of tomato aroma. Genes 2021, 12, 226. [Google Scholar] [CrossRef]
- Neri, F.; Cappellin, L.; Spadoni, A.; Cameldi, I.; Algarra Alarcon, A.; Aprea, E.; Romano, A.; Gasperi, F.; Biasioli, F. Role of strawberry volatile organic compounds in the development of Botrytis cinerea infection. Plant Pathol. 2015, 64, 709–717. [Google Scholar] [CrossRef]
- Xu, Y.; Tong, Z.; Zhang, X.; Zhang, X.; Luo, Z.; Shao, W.; Li, L.; Ma, Q.; Zheng, X.; Fang, W. Plant volatile organic compound (E)-2-hexenal facilitates Botrytis cinerea infection of fruits by inducing sulfate assimilation. New. Phytol. 2021, 231, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Koeduka, T. Green leaf volatiles in plant signaling and response. Subcell. Biochem. 2016, 86, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Chen, T.; Xu, Y.; Tian, S. Antifungal effects of hinokitiol on development of Botrytis cinerea in vitro and in vivo. Postharvest Biol. Technol. 2020, 159, 111038. [Google Scholar] [CrossRef]
- Weber, H. Fatty acid-derived signals in plants. Trends Plant Sci. 2002, 7, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef]
- Garbowicz, K.; Liu, Z.; Alseekh, S.; Tieman, D.; Taylor, M.; Kuhalskaya, A.; Ofner, I.; Zamir, D.; Klee, H.J.; Fernie, A.R.; et al. Quantitative trait loci analysis identifies a prominent gene involved in the production of fatty acid-derived flavor volatiles in tomato. Mol. Plant. 2018, 11, 1147–1165. [Google Scholar] [CrossRef]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Fungicidal activity of plant volatile compounds for controlling Monilinia laxa in stone fruit. Plant Dis. 2007, 91, 30–35. [Google Scholar] [CrossRef]
- He, P.Q.; Tian, L.; Chen, K.S.; Hao, L.H.; Li, G.Y. Induction of volatile organic compounds of Lycopersicon esculentum Mill. and its resistance to Botrytis cinerea Pers. by burdock oligosaccharide. J. Integr. Plant Biol. 2006, 48, 550–557. [Google Scholar] [CrossRef]
- Guenther, A.; Karl, T.; Harley, P.; Wiedinmyer, C.; Palmer, P.I.; Geron, C. Estimates of global terrestrial isoprene emissions using MEGAN (Model of emissions of gases and aerosols from nature). Atmos. Chem. Phys. 2006, 6, 3181–3210. [Google Scholar] [CrossRef]
- Lipko, A.; Swiezewska, E. Isoprenoid generating systems in plants—A handy toolbox how to assess contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthetic process. Prog. Lipid Res. 2016, 63, 70–92. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biot. 2015, 148, 63–106. [Google Scholar] [CrossRef]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef]
- Hall, D.E.; Robert, J.A.; Keeling, C.I.; Domanski, D.; Quesada, A.L.; Jancsik, S.; Kuzyk, M.A.; Hamberger, B.; Borchers, C.H.; Bohlmann, J. An integrated genomic, proteomic and biochemical analysis of (+)-3-carene biosynthesis in Sitka spruce (Picea sitchensis) genotypes that are resistant or susceptible to white pine weevil. Plant J. 2011, 65, 936–948. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Venables, B.; Petros, R.A.; Nalam, V.; Li, M.Y.; Wang, X.M.; Takemoto, L.J.; Shah, J. An abietane diterpenoid is a potent activator of systemic acquired resistance. Plant J. 2012, 71, 161–172. [Google Scholar] [CrossRef]
- Simas, D.L.; de Amorim, S.H.; Goulart, F.R.; Alviano, C.S.; Alviano, D.S.; da Silva, A.J.R. Citrus species essential oils and their components can inhibit or stimulate fungal growth in fruit. Ind. Crop. Prod. 2017, 98, 108–115. [Google Scholar] [CrossRef]
- Sekine, T.; Sugano, M.; Majid, A.; Fujii, Y. Antifungal effects of volatile compounds from black zira (Bunium persicum) and other spices and herbs. J. Chem. Ecol. 2007, 33, 2123–2132. [Google Scholar] [CrossRef]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef]
- Faize, L.; Faize, M. Functional analogues of salicylic acid and their use in crop protection. Agronomy 2018, 8, 5. [Google Scholar] [CrossRef]
- Paul, D.; Park, K.S. Identification of volatiles produced by Cladosporium cladosporioides CL-1, a fungal biocontrol agent that promotes plant growth. Sensors 2013, 13, 13969–13977. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tang, L.; Hu, J.; Wang, J.; Adelakun, T.A.; Yang, D.; Di, Y.; Zhang, Y.; Hao, X. Munronin O, a potential activator for plant resistance. Pestic. Biochem. Phys. 2018, 146, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; He, M.; Xiang, X.W.; Adnan, M.; Cui, Z.N. Novel S-thiazol-2-yl-furan-2-carbothioate derivatives as potential T3SS inhibitors against Xanthomonas oryzae on rice. J. Agric. Food Chem. 2019, 67, 11867–11876. [Google Scholar] [CrossRef] [PubMed]
- Narsibhai, B.D.K.; Mishra, D.; Vyavahare, L.V.; Singh, A. Synthesis, characterization and antimicrobial activity of novel heterocyclic compounds. J. Curr. Chem. Pharm. Sci. 2012, 2, 233–239. [Google Scholar]
- Rahman, M.; Mostofa, M.G.; Islam, M.; Keya, S.S.; Das, A.K.; Miah, M.G.; Kawser, A.Q.M.R.; Ahsan, S.M.; Hashem, A.; Tabassum, B.; et al. Acetic acid: A cost-effective agent for mitigation of seawater-induced salt toxicity in mung bean. Sci. Rep. 2019, 9, 15186. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.; Augusto, C.; Reis, P.; Rego, C.; Figueiredo, A.C.; Fortes, A.M. Volatile metabolism of wine grape Trincadeira: Impact of infection with Botrytis cinerea. Plants 2022, 11, 141. [Google Scholar] [CrossRef]
- Emanuel, R.V.; César Arturo, P.U.; Lourdes Iveth, M.R.; de la Cruz Homero, R.; Mauricio Nahuam, C.A. In vitro growth of Colletotrichum gloeosporioides is affected by butyl acetate, a compound produced during the co-culture of Trichoderma sp. and Bacillus subtilis. 3 Biotech 2020, 10, 329. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| VOC | Perkoz | Remiz | ||||||
|---|---|---|---|---|---|---|---|---|
| C | TRS 106 | Bc | TRS 106 + Bc | C | TRS 106 | Bc | TRS 106 + Bc | |
| Alcohols | ||||||||
| 1-Penten-3-ol | 4.3 × 107 ± 4.1 × 106 (c) | 1.8 × 107 ± 1.5 × 106 (b) | 1.3 × 107 ± 1.8 × 106 (b) | 1.6 × 107 ± 3.5 × 106 (b) | 1.5 × 106 ± 7.6 × 104 (a) | 8.9 × 105 ± 2.9 × 104 (a) | 3.4 × 105 ± 1.9 × 104 (a) | 2.4 × 106 ± 3.5 × 105 (a) |
| 4-Methyl-3-hexanol | nd | 8.5 × 105 ± 3.7 × 104 (a) | 8.5 × 105 ± 3.3 × 104 (a) | 6.5 × 105 ± 1.9 × 104 (a) | nd | nd | nd | nd |
| 2-Penten-1-ol | 1.8 × 107± 2.4 × 106 (b) | 1.5 × 107 ± 7.0 × 106 (b) | 2.8 × 107 ± 8.5 × 106 (bc) | 3.4 × 107 ± 4.5 × 106 (c) | 1.3 × 105 ± 4.1 × 104 (a) | 6.3 × 105 ± 1.2 × 105 (a) | nd | nd |
| Prenol | 7.7 × 106 ± 6.8 × 105 (bc) | 4.4 × 106 ± 5.8 × 105 (b) | 7.8 × 106 ± 1.0 × 106 (bc) | 9.0 × 106 ± 4.2 × 105 (c) | 1.4 × 105 ± 3.2 × 104 (a) | 4.6 × 106 ± 1.7 × 105 (b) | 8.0 × 105 ± 4.7 × 104 (a) | 4.6 × 106 ± 2.5 × 105 (b) |
| Cyclopentanol | nd | nd | nd | 2.1 × 106 ± 1.8 × 105 (ab) | 3.3 × 106 ± 4.6 × 105 (ab) | 3.4 × 106 ± 5.3 × 105 (ab) | 4.6 × 106 ± 3.4 × 105 (b) | 1.4 × 106 ± 4.0 × 105 (a) |
| 1-Pentanol | 3.5 × 107 ± 3.9 × 106 (c) | 2.4 × 107 ± 1.8 × 106 (bc) | 2.8 × 107 ± 5.0 × 106 (bc) | 2.1 × 107 ± 4.9 × 106 (bc) | 1.4 × 106 ± 8.4 × 105 (a) | 6.2 × 106 ± 2.1 × 105 (ab) | 5.7 × 106 ± 4.4 × 105 (ab) | 1.1 × 107 ± 8.4 × 104 (b) |
| 2-Hexanol | 6.3 × 105 ± 2.3 × 104 (b) | 2.1 × 106 ± 8.6 × 104 (c) | nd | 1.6 × 106 ± 3.9 × 105 (c) | 1.1 × 105 ± 8.0 × 103 (a) | 2.3 × 105 ± 1.8 × 104 (a) | 1.3 × 105 ± 2.9 × 104 (a) | 1.7 × 105 ± 1.6 × 104 (a) |
| 2-Hexyn-1-ol | nd | 7.2 × 108 ± 1.7 × 107 (b) | nd | 6.6 × 107 ± 3.8 × 106 (a) | 2.3 × 107 ± 1.2 × 106 (a) | 2.4 × 107 ± 3.0 × 106 (a) | 2.3 × 107 ± 2.4 × 106 (a) | 5.2 × 107 ± 8.4 × 105 (a) |
| 2-Hexen-1-ol | nd | nd | 2.7 × 106 ± 2.6 × 105 (b) | 1.5 × 105 ± 1.0 × 104 (a) | 6.0 × 106 ± 4.7 × 105 (b) | 1.1 × 107 ± 9.4 × 104 (abc) | 1.8 × 107 ± 3.9 × 106 (c) | 7.2 × 106 ± 1.3 × 105 (b) |
| 1-Hexanol | 5.9 × 107 ± 3.5 × 106 (b) | 5.4 × 107 ± 5.4 × 106 (b) | 5.5 × 107 ± 5.0 × 106 (b) | 6.1 × 107 ± 9.5 × 105 (b) | 1.6 × 106 ± 4.6 × 104 (a) | 2.6 × 106 ± 3.1 × 105 (a) | 2.3 × 106 ± 2.4 × 105 (a) | 2.4 × 106 ± 3.0 × 105 (a) |
| 1-Heptanol | 2.4 × 107 ± 2.7 × 106 (c) | 1.1 × 107 ± 1.3 × 106 (b) | nd | 2.3 × 107 ± 4.1 × 106 (c) | 2.3 × 107 ± 3.6 × 106 (a) | 2.2 × 106 ± 3.1 × 105 (ab) | 2.3 × 106 ± 1.1 × 105 (ab) | 2.4 × 106 ± 4.9 × 105 (ab) |
| 1,5-Octadien-3-ol | nd | 2.8 × 106 ± 5.0 × 105 (b) | nd | 3.4 × 106 ± 5.4 × 105 (b) | nd | 1.5 × 105 ± 3.8 × 104 (a) | 1.2 × 106 ± 2.6 × 105 (b) | 1.2 × 107 ± 2.1 × 106 (c) |
| 7-Octen-4-ol | 3.6 × 106 ± 6.7 × 105 (b) | nd | 2.2 × 106 ± 3.8 × 105 (b) | 1.6 × 106 ± 4.2 × 105 (b) | 2.3 × 105 ± 4.7 × 104 (a) | 2.3 × 105 ± 5.2 × 104 (a) | 1.2 × 106 ± 1.4 × 105 (ab) | 1.6 × 106 ± 3.6 × 105 (b) |
| 1-Octen-3-ol | nd | 2.1 × 106 ± 2.9 × 105 (a) | nd | 1.7 × 106 ± 2.1 × 105 (a) | nd | nd | nd | nd |
| Phenol | 2.5 × 106 ± 1.1 × 105 (b) | 3.1 × 106 ± 2.1 × 105 (b) | 3.2 × 106 ± 1.8 × 105 (b) | 2.5 × 106 ± 1.3 × 105 (b) | 2.5 × 105 ± 6.1 × 104 (a) | 1.3 × 106 ± 2.4 × 105 (ab) | 6.7 × 106 ± 5.4 × 105 (c) | 2.2 × 106 ± 3.5 × 105 (b) |
| 6-Methyl-5-hepten-2-ol | 1.9 × 106 ± 2.1 × 105 (bc) | 1.6 × 106 ± 5.5 × 105 (bc) | 2.1 × 106 ± 3.3 × 105 (c) | 1.3 × 106 ± 3.4 × 105 (bc) | nd | 2.3 × 105 ± 3.4 × 104 (a) | 1.2 × 105 ± 1.9 × 104 (a) | 5.6 × 105 ± 3.9 × 104 (ab) |
| Isooctanol | 3.1 × 106 ± 2.2 × 105 (c) | 2.7 × 106 ± 3.1 × 105 (c) | 3.8 × 106 ± 3.7 × 105 (c) | 2.2 × 106 ± 1.9 × 105 (c) | 3.2 × 105 ± 3.3 × 104 (b) | 2.3 × 105 ± 3.5 × 104 (ab) | 1.2 × 105 ± 3.7 × 104 (a) | 2.7 × 106 ± 4.7 × 105 (c) |
| 2-Ethyl-1-hexanol | 5.1 × 107 ± 9.9 × 105 (b) | 7.9 × 107 ± 1.3 × 106 (c) | 6.4 × 107 ± 3.7 × 106 (bc) | 5.9 × 107 ± 3.6 × 106 (bc) | 3.6 × 105 ± 3.0 × 104 (a) | 4.5 × 106 ± 3.4 × 105 (b) | 3.2 × 106 ± 2.5 × 105 (ab) | 4.3 × 106 ± 2.9 × 105 (ab) |
| 2-Methylphenol | 9.4 × 105 ± 2.4 × 104 (c) | 9.9 × 105 ± 5.2 × 104 (c) | 5.9 × 105 ± 7.5 × 104 (bc) | 7.9 × 105 ± 6.0 × 104 (c) | 2.3 × 104 ± 6.9 × 103 (a) | 3.4 × 105 ± 4.0 × 104 (b) | 1.6 × 105 ± 6.7 × 104 (ab) | 6.9 × 104 ± 6.9 × 103 (ab) |
| 1-Octanol | 2.5 × 106 ± 8.7 × 104 (c) | 1.9 × 106 ± 1.9 × 105 (c) | 2.5 × 106 ± 2.0 × 105 (c) | 1.7 × 106 ± 2.8 × 105 (c) | 5.2 × 105 ± 4.1 × 104 (ab) | 5.3 × 105 ± 3.7 × 104 (ab) | 2.2 × 105 ± 2.3 × 104 (a) | 1.2 × 106 ± 2.0 × 104 (bc) |
| 2-Methoxyphenol | 2.8 × 106 ± 9.4 × 105 (c) | 3.6 × 105 ± 9.7 × 105 (c) | nd | nd | 1.2 × 105 ± 1.0 × 104 (a) | 1.7 × 106 ± 9.7 × 104 (b) | 1.6 × 105 ± 2.2 × 104 (a) | 1.1 × 106 ± 1.6 × 105 (abc) |
| 2-Nonen-1-ol | nd | 9.1 × 105 ± 9.5 × 103 (c) | 8.7 × 105 ± 2.9 × 104 (bc) | 6.2 × 105 ± 2.6 × 104 (b) | nd | 2.6 × 105 ± 4.0 × 104 (ab) | 1.1 × 105 ± 2.2 × 104 (a) | 2.5 × 105 ± 4.5 × 104 (ab) |
| Phenylethanol | 4.3 × 106 ± 3.7 × 105 (a) | 5.8 × 106 ± 8.0 × 105 (a) | 6.8 × 106 ± 7.1 × 105 (a) | 6.6 × 106 ± 6.7 × 105 (a) | 4.3 × 106 ± 3.7 × 105 (a) | 4.4 × 106 ± 4.0 × 105 (a) | 6.7 × 107 5.8 × 106 (c) | 3.4 × 107 ± 5.2 × 106 (b) |
| Aldehydes | ||||||||
| Pentanal | 7.3 × 106 ± 3.6 × 105 (bc) | 7.8 × 106 ± 9.9 × 105 (c) | 3.2 × 106 ± 1.7 × 105 (b) | 3.5 × 106 ± 4.1 × 105 (b) | 2.2 × 104 ± 8.1 × 102 (a) | 2.3 × 105 ± 6.7 × 102 (a) | 2.1 × 106 ± 8.8 × 104 (ab) | 2.3 × 106 ± 9.3 × 105 (ab) |
| 2-Pentenal | 4.2 × 106 ± 1.4 × 105 (b) | 1.0 × 107 ± 9.3 × 105 (c) | 4.5 × 106 ± 1.5 × 105 (bc) | 2.4 × 106 ± 3.5 × 104 (ab) | 2.9 × 105 ± 1.6 × 104 (a) | 1.9 × 105 ± 5.9 × 103 (a) | 1.7 × 106 ± 1.1 × 105 (ab) | 2.9 × 106 ± 8.7 × 104 (ab) |
| 2-Methyl-4-pentenal | 2.5 × 108 ± 4.8 × 107 (b) | 2.5 × 106 ± 7.0 × 105 (a) | 4.0 × 106 ± 9.0 × 105 (a) | 1.3 × 108 ± 9.7 × 106 (b) | 5.3 × 106 ± 2.6 × 105 (a) | 5.6 × 107 ± 1.0 × 106 (ab) | 6.7 × 107 ± 1.2 × 107 (ab) | 4.6 × 107 ± 8.2 × 106 (ab) |
| Hexanal | 1.8 × 108 ± 1.7 × 107 (b) | 1.5 × 108 ± 1.9 × 105 (b) | 2.6 × 108 ± 1.8 × 107 (b) | 2.3 × 108 ± 7.4 × 106 (b) | 7.3 × 106 ± 8.1 × 105 (a) | 2.7 × 107 ± 6.7 × 106 (a) | 1.9 × 107 ± 6.0 × 105 (a) | 1.7 × 107 ± 2.2 × 106 (a) |
| 3-Hexenal | 4.5 × 108 ± 1.4 × 107 (b) | 4.6 × 108 ± 5.0 × 107 (b) | 2.2 × 108 ± 1.3 × 107 (ab) | 5.1 × 108 ± 2.2 × 107 (b) | 3.3 × 107 ± 3.8 × 106 (a) | 3.6 × 107 ± 2.0 × 106 (a) | 2.9 × 107 ± 1.6 × 106 (a) | 3.9 × 107 ± 2.9 × 106 (a) |
| 2-Hexenal | 1.7 × 109 ± 1.0 × 108 (c) | 2.5 × 109 ± 2.5 × 108 (d) | 2.2 × 109 ± 2.4 × 108 (cd) | 2.2 × 109 ± 3.6 × 108 (cd) | 1.5 × 107 ± 1.2 × 106 (a) | 2.4 × 107 ± 6.7 × 106 (a) | 3.3 × 107 ± 4.6 × 106 (a) | 9.8 × 107 ± 2.5 × 106 (b) |
| (Z)-4-Heptenal | 7.6 × 106 ± 5.0 × 104 (b) | 9.8 × 106 ± 1.5 × 105 (c) | 7.7 × 106 ± 2.5 × 105 (b) | 8.0 × 106 ± 2.8 × 105 (bc) | 4.7 × 104 ± 2.1 × 103 (a) | 5.4 × 105 ± 1.1 × 104 (ab) | 2.4 × 106 ± 5.0 × 105 (a) | 6.5 × 106 ± 9.4 × 104 (b) |
| (2E,4E)-2,4-Hexadienal | 2.6 × 107 ± 1.7 × 106 (b) | 1.9 × 107 ± 1.2 × 106 (ab) | 3.4 × 107 ± 6.0 × 106 (c) | 3.3 × 107 ± 6.8 × 106 (c) | 1.4 × 107 ± 4.2 × 106 (a) | 2.7 × 107 ± 7.3 × 106 (bc) | 1.1 × 107 ± 1.2 × 106 (a) | 1.8 × 107 ± 6.0 × 106 (a) |
| Heptanal | 7.6 × 106 ± 2.2 × 105 (bc) | 9.7 × 106 ± 2.5 × 105 (bc) | 1.1 × 107 ± 1.1 × 106 (c) | 8.0 × 106 ± 2.3 × 105 (bc) | 2.3 × 105 ± 3.0 × 104 (a) | 2.3 × 105 ± 3.6 × 104 (a) | 2.6 × 105 ± 1.4 × 104 (a) | 1.2 × 106 ± 1.9 × 105 b) |
| (E)-2-Heptenal | 1.6 × 106 ± 1.8 × 105 (ab) | 2.4 × 106 ± 1.7 × 105 (b) | 1.8 × 106 ± 2.8 × 105 (ab) | 1.8 × 106 ± 1.9 × 105 (ab) | 1.3 × 105 ± 4.8 × 103 (a) | 1.6 × 105 ± 2.2 × 104 (a) | 1.2 × 105 ± 2.0 × 104 (a) | 1.5 × 105 ± 3.6 × 104 (a) |
| (E)-4-Oxohex-2-enal | 5.4 × 107 ± 3.4 × 106 (a) | 2.2 × 108 ± 6.9 × 106 (b) | 1.4 × 108 ± 2.2 × 107 (ab) | 6.9 × 107 ± 4.3 × 106 (a) | 1.8 × 108 ± 1.7 × 107 (ab) | 2.1 × 108 ± 1.4 × 107 (ab) | 1.9 × 108 ± 4.6 × 107 (ab) | 1.3 × 108 ± 6.0 × 106 (ab) |
| Benzaldehyde | 3.4 × 107 ± 5.6 × 106 (bc) | 8.2 × 107 ± 4.0 × 106 (c) | 1.1 × 107 ± 1.8 × 106 (b) | 3.6 × 107 ± 4.1 × 106 (bc) | 2.1 × 104 ± 2.3 × 103 (a) | 2.4 × 104 ± 9.6 × 102 (a) | 2.3 × 103 ± 2.1 × 102 (a) | 2.8 × 104 ± 2.7 × 103 (a) |
| Octanal | nd | 2.3 × 107 ± 3.8 × 106 ab() | 3.6 × 107 ± 2.6 × 106 (b) | 2.7 × 107 ± 6.6 × 106 (b) | 2.4 × 105 ± 3.8 × 104 (a) | 5.5 × 105 ± 3.5 × 104 (a) | 2.1 × 106 ± 2.4 × 105 (a) | 3.3 × 106 ± 2.9 × 105 (a) |
| (Z)-6-Nonenal | nd | 2.5 × 106 ± 3.1 × 105 (c) | nd | 1.7 × 106 ± 8.5 × 104 (c) | nd | 1.6 × 105 ± 1.3 × 104 (a) | nd | 2.4 × 105 ± 2.1 × 104 (bc) |
| (2E,4E)-2,4-Heptadienal | 9.9 × 106 ± 2.2 × 105 (ab) | 1.3 × 107 ± 1.6 × 106 (ab) | 1.3 × 107 ± 3.6 × 106 (ab) | 1.4 × 107 ± 1.5 × 106 (ab) | 3.7 × 106 ± 1.8 × 105 (a) | 2.3 × 107 ± 2.4 × 106 (b) | 1.2 × 107 ± 1.4 × 106 (ab) | 7.2 × 106 ± 1.5 × 105 (ab) |
| 2-Hydroxybenzaldehyde | 3.7 × 105 ± 3.4 × 104 (b) | nd | nd | nd | 2.3 × 105 ± 8.1 × 103 (ab) | 1.1 × 106 ± 1.9 × 105 (b) | 3.3 × 104 ± 5.5 × 102 (a) | 1.1 × 105 ± 9.6 × 103 (ab) |
| 2-Octenal | nd | 1.6 × 106 ± 2.5 × 105 (a) | nd | 2.2 × 106 ± 6.2 × 104 (a) | 6.1 × 105 ± 3.0 × 104 (a) | 2.1 × 107 ± 3.3 × 106 (b) | 6.4 × 105 ± 1.8 × 104 (a) | 1.2 × 107 ± 2.3 × 106 (b) |
| 2,6-Dimethyl-5-heptenal | 4.0 × 106 ± 3.4 × 105 (b) | 3.4 × 106 ± 5.3 × 105 (ab) | 3.5 × 106 ± 5.1 × 105 (ab) | 3.2 × 106 ± 2.8 × 105 (ab) | nd | nd | nd | 1.6 × 106 ± 2.0 × 105 (a) |
| 4-Methylbenzaldehyde | nd | 1.2 × 106 ± 2.5 × 105 (a) | nd | nd | 6.8 × 103 ± 1.2 × 103 (a) | 2.1 × 105 ± 4.1 × 104 (ab) | 1.7 × 105 ± 3.2 × 104 (ab) | 7.8 × 104 ± 4.2 × 103 (ab) |
| Nonanal | 3.1 × 106 ± 3.1 × 105 (ab) | 3.4 × 106 ± 3.2 × 105 (b) | 3.1 × 106 ± 2.0 × 105 (ab) | 3.0 × 10 ± 2.8 × 105 (ab) | 2.2 × 106 ± 2.3 × 105 (ab) | 1.2 × 106 ± 2.1 × 105 (ab) | 2.3 × 105 ± 6.4 × 103 (a) | 1.5 × 106 ± 3.2 × 105 (ab) |
| (E,Z)-2,6-Nonadienal | 2.2 × 106 ± 2.7 × 105 (bc) | 3.1 × 106 ± 2.2 × 105 (c) | 1.9 × 106 ± 1.9 × 105 (bc) | 1.5 × 106 ± 1.7 × 105 (bc) | nd | 2.2 × 105 ± 1.8 × 104 (a) | 4.4 × 105 ± 5.0 × 104 (ab) | 1.6 × 105 ± 2.4 × 104 (a) |
| 4-Ethylbenzaldehyde | 3.4 × 106 ± 1.5 × 105 (b) | 4.7 × 106 ± 9.4 × 105 (b) | 4.3 × 106 ± 1.7 × 105 (b) | 4.6 × 106 ± 1.7 × 105 (b) | 1.3 × 106 ± 2.2 × 105 (a) | 2.3 × 106 ± 2.2 × 105 (ab) | 8.0 × 105 ± 5.6 × 104 (ab) | 8.0 × 105 ± 6.1 × 104 (a) |
| 3-Thujen-10-al | 5.2 × 105 ± 2.6 × 104 (a) | 4.2 × 106 ± 2.0 × 105 (b) | nd | nd | 4.7 × 105 ± 1.3 × 104 (a) | 5.7 × 105 ± 2.3 × 104 (a) | 5.1 × 105 ± 1.7 × 104 (a) | 1.4 × 106 ± 2.0 × 105 (a) |
| Decanal | 1.1 × 106 ± 1.1 × 105 (c) | 6.7 × 105 ± 3.6 × 104 (bc) | 9.7 × 105 ± 7.9 × 103 (c) | nd | 2.3 × 104 ± 3.7 × 103 (a) | 2.2 × 105 ± 5.9 × 104 (b) | 2.3 × 104 ± 2.2 × 103 (a) | 3.2 × 105 ± 3.1 × 104 (b) |
| Ketones | ||||||||
| 1-Penten-3-one | 1.7 × 107 ± 1.3 × 106 (b) | 1.7 × 107 ± 7.2 × 105 (b) | 1.3 × 107 ± 9.5 × 105 (b) | 1.7 × 107 ± 2.9 × 106 (b) | 2.8 × 106 ± 3.5 × 105 (a) | 1.3 × 106 ± 2.0 × 105 (ab) | 4.3 × 105 ± 2.8 × 104 (a) | 1.5 × 106 ± 2.4 × 105 (a) |
| 3-Pentanone | 1.5 × 106 ± 1.2 × 105 (ab) | 1.4 × 106 ± 3.8 × 105 (ab) | 1.5 × 106 ± 3.7 × 105 (ab) | nd | 2.2 × 105 ± 3.5 × 104 (a) | 5.9 × 106 ± 5.1 × 105 (b) | nd | nd |
| 5-Methyl-2-hexanone | nd | 2.9 × 105 ± 4.9 × 103 (abc) | nd | nd | 2.9 × 105 ± 1.7 × 104 (ab) | 1.1 × 105 ± 2.3 × 104 (abc) | 2.6 × 105 ± 1.4 × 104 (b) | 8.2 × 105 ± 3.5 × 104 (c) |
| 5-Ethyl-2(5H)-furanone | nd | nd | 7.7 × 103 ± 6.6 × 102 (a) | nd | nd | nd | 6.4 × 103 ± 1.8 × 102 (a) | nd |
| 6-Methyl-2-heptanone | nd | nd | nd | 7.5 × 105 ± 3.4 × 104 (a) | nd | nd | nd | 6.7 × 105 ± 3.6 × 104 (a) |
| 1-Octen-3-one | 1.6 × 106 ± 2.3 × 105 (bc) | 1.8 × 106 ± 5.0 × 105 (bc) | 1.6 × 106 ± 6.0 × 105 (c) | nd | 1.6 × 105 ± 4.9 × 104 (a) | 2.3 × 105 ± 4.9 × 104 (b) | 1.5 × 105 ± 4.2 × 104 (ab) | 2.2 × 105 ± 3.5 × 104 (abc) |
| 6-Methyl-5-hepten-2-one | 2.4 × 106 ± 2.6 × 105 (b) | 2.2 × 106 ± 1.8 × 105 (b) | 2.1 × 106 ± 3.3 × 105 (b) | 2.1 × 105 ± 3.8 × 104 (b) | 2.4 × 105 ± 5.0 × 104 (ab) | 2.2 × 105 ± 2.3 × 103 (a) | 1.4 × 105 ± 4.9 × 103 (a) | 2.3 × 106 ± 3.0 × 105 (b) |
| 2,2,6-Trimethyl-cyclohexanone | 8.7 × 105 ± 5.6 × 104 (ab) | 1.8 × 106 ± 4.8 × 105 (bc) | nd | 1.4 × 106 ± 5.0 × 105 (ab) | 9.7 × 105 ± 1.6 × 104 (ab) | 1.0 × 105 ± 4.5 × 103 (a) | 2.3 × 106 ± 2.3 × 105 (c) | 2.4 × 106 ± 2.7 × 105 (c) |
| Acetophenone | 4.2 × 106 ± 2.2 × 105 (b) | 1.3 × 106 ± 3.5 × 105 (ab) | 1.5 × 106 ± 3.8 × 105 (ab) | 2.4 × 106 ± 2.8 × 105 (ab) | 1.3 × 106 ± 3.7 × 105 (a) | 4.7 × 106 ± 5.7 × 105 (b) | 8.0 × 106 ± 1.0 × 106 (bc) | 1.3 × 107 ± 6.5 × 105 (c) |
| Ketoisophorone | nd | nd | nd | 6.2 × 105 ± 3.6 × 104 (a) | 4.7 × 105 ± 7.0 × 104 (b) | 4.6 × 105 ± 7.1 × 104 (b) | 4.2 × 105 ± 4.2 × 104 (b) | 4.3 × 105 ± 3.7 × 104 (b) |
| 2-Hydroxyacetophenone | 6.0 × 105 ± 3.4 × 104 (a) | 3.3 × 106 ± 1.7 × 105 (a) | 3.1 × 106 ± 1.4 × 105 (ab) | 3.4 × 106 ± 4.5 × 105 (ab) | 1.7 × 105 ± 1.9 × 104 (a) | 6.6 × 107 ± 1.2 × 106 (b) | 3.2 × 106 ± 2.0 × 105 (a) | 6.5 × 107 ± 9.9 × 105 (b) |
| 3,6,6-Trimethyl-2-cyclohexen-1-one | 3.9 × 105 ± 4.0 × 104 (a) | 7.2 × 105 ± 3.8 × 104 (a) | 3.3 × 105 ± 4.4 × 104 (a) | 3.9 × 105 ± 6.2 × 104 (a) | 8.9 × 105 ± 8.0 × 104 (a) | 9.2 × 105 ± 2.3 × 104 (a) | 4.7 × 106 ± 2.3 × 105 (a) | 6.9 × 107 ± 2.5 × 106 (b) |
| Esters | ||||||||
| Methyl acetate | 1.5 × 106 ± 2.0 × 105 (bc) | 8.3 × 105 ± 6.0 × 104 (b) | 1.5 × 106 ± 4.1 × 105 (bc) | 4.7 × 105 ± 3.0 × 104 (ab) | 1.0 × 104 ± 3.0 × 102 (a) | 1.8 × 106 ± 3.5 × 105 (c) | 7.1 × 105 ± 3.9 × 104 (ab) | 4.3 × 105 ± 3.2 × 104 (ab) |
| Ethyl acetate | 2.0 × 107 ± 4.8 × 105 (c) | 5.1 × 106 ± 3.0 × 105 (b) | 5.1 × 106 ± 4.7 × 105 (b) | 4.9 × 106 ± 2.9 × 105 (b) | 1.9 × 106 ± 2.4 × 105 (a) | 2.1 × 106 ± 4.2 × 105 (a) | 1.5 × 106 ± 6.6 × 105 (a) | 3.6 × 106 ± 5.6 × 105 (b) |
| Butyl acetate | 6.5 × 106 ± 6.9 × 105 (c) | 2.2 × 106 ± 2.8 × 105 (b) | 8.3 × 105 ± 7.3 × 104 (a) | 1.1 × 106 ± 2.2 × 105 (ab) | 7.4 × 105 ± 3.9 × 104 (a) | 2.3 × 106 ± 3.3 × 105 (b) | 6.9 × 105 ± 7.0 × 104 (a) | 6.5 × 106 ± 4.1 × 105 (c) |
| Hexyl acetate | 1.5 × 107 ± 2.3 × 106 (bc) | 5.7 × 106 ± 3.8 × 105 (b) | 1.1 × 107 ± 8.2 × 105 (bc) | 5.3 × 106 ± 2.8 × 105 (bc) | 1.3 × 106 ± 2.0 × 105 (a) | 3.6 × 106 ± 3.6 × 105 (ab) | 3.9 × 106 ± 5.5 × 105 (ab) | 2.6 × 107 ± 2.2 × 106 (c) |
| Butyl hexanoate | 5.1 × 106 ± 2.8 × 105 (b) | nd | 8.3 × 105 ± 7.3 × 104 (a) | nd | nd | nd | nd | nd |
| Methyl salicylate | 1.3 × 104 ± 2.1 × 103 (a) | 1.3 × 105 ± 4.9 × 104 (bc) | 5.8 × 104 ± 3.3 × 103 (ab) | 1.3 × 105 ± 2.0 × 104 ()bc | 1.1 × 104 ± 1.8 × 103 (a) | 3.9 × 105 ± 4.8 × 104 (c) | 3.1 × 105 ± 2.3 × 104 (bc) | 4.5 × 105 ± 2.5 × 104 (c) |
| Ethyl salicylate | 5.1 × 105 ± 2.1 × 104 (a) | 8.2 × 105 ± 1.8 × 104 (ab) | 6.1 × 105 ± 7.6 × 104 (a) | 5.1 × 105 ± 1.4 × 104 (a) | 6.0 × 106 ± 1.4 × 105 (b) | 7.6 × 106 ± 6.9 × 105 (bc) | 2.3 × 106 ± 2.5 × 105 (b) | 4.4 × 107 ± 2.5 × 106 (c) |
| Isoamyl salicylate | nd | nd | nd | nd | nd | 1.1 × 104 ± 8.7 × 102 (a) | nd | 2.4 × 105 ± 2.7 × 104 (a) |
| Terpenoids | ||||||||
| α-Pinene | 4.6 × 105 ± 6.4 × 104 (b) | 2.8 × 105 ± 6.4 × 104 (a) | 3.7 × 105 ± 4.3 × 104 (ab) | 2.2 × 105 ± 1.4 × 104 (a) | 6.4 × 106 ± 2.6 × 105 (c) | 6.2 × 106 ± 2.7 × 105 (c) | 3.3 × 106 ± 1.8 × 105 (ab) | 3.5 × 107 ± 3.9 × 106 (abc) |
| (-)-Sabinene | 2.6 × 108 ± 3.2 × 107 (b) | 2.9 × 106 ± 6.1 × 105 (a) | nd | nd | 4.6 × 105 ± 4.5 × 104 (a) | 6.3 × 106 ± 2.6 × 105 (a) | nd | nd |
| β-Pinene | 2.9 × 106 ± 5.3 × 105 (c) | nd | 3.5 × 106 ± 3.0 × 105 (c) | 2.4 × 106 ± 1.3 × 105 (c) | 5.5 × 103 ± 6.9 × 101 (a) | 1.3 × 105 ± 2.6 × 104 (b) | 1.3 × 105 ± 1.1 × 104 (b) | 1.3 × 105 ± 2.1 × 104 (b) |
| β-Myrcene | 1.6 × 107 ± 6.5 × 106 (a) | 2.7 × 107 ± 5.9 × 106 (ab) | 1.6 × 107 ± 6.2 × 106 (a) | 2.0 × 107 4.7 × 106 (ab) | 4.7 × 105 ± 6.3 × 104 (a) | 1.7 × 108 ± 4.9 × 107 (c) | 3.2 × 107 ± 4.2 × 106 (ab) | 7.3 × 107 ± 4.5 × 106 (bc) |
| α-Phellandrene | 3.2 × 107 ± 1.6 × 106 (c) | 1.3 × 107 ± 1.4 × 106 (bc) | 1.9 × 107 ± 4.0 × 106 (c) | 2.4 × 106 ± 1.6 × 105 (a) | 3.4 × 107 ± 8.9 × 105 (c) | 2.6 × 107 ± 2.1 × 106 (b) | 1.3 × 107 ± 1.7 × 106 (abc) | 1.0 × 107 ± 8.1 × 105 (ab) |
| 2-Carene | 1.7 × 108 ± 9.2 × 106 (b) | 9.5 × 107 ± 6.7 × 105 (b) | 1.4 × 108 ± 3.4 × 107 (b) | 9.3 × 107 ± 8.9 × 106 (ab) | 4.7 × 106 ± 7.8 × 104 (a) | 4.7 × 104 ± 1.4 × 103 (a) | 1.2 × 107 ± 6.2 × 105 (ab) | 2.3 × 108 ± 4.1 × 107 (b) |
| α-Terpinene | 7.3 × 105 ± 1.9 × 104 (ab) | 1.3 × 105 ± 1.1 × 104 (a) | 1.1 × 107 ± 9.2 × 105 (bc) | 3.1 × 106 ± 1.9 × 105 (b) | 3.6 × 106 ± 3.0 × 105 (b) | 2.2 × 107 ± 1.8 × 106 (c) | 1.1 × 107 ± 1.7 × 106 (b) | 1.7 × 107 ± 2.5 × 106 (bc) |
| Limonene | 2.1 × 108 ± 2.3 × 107 (b) | 1.5 × 108 ± 7.9 × 106 (ab) | 1.9 × 108 ± 1.9 × 107 (b) | 1.6 × 108 ± 3.2 × 107 (ab) | 1.2 × 105 ± 2.2 × 104 (a) | 2.3 × 104 ± 3.1 × 103 (a) | 1.4 × 107 ± 2.8 × 106 (a) | 1.2 × 107 ± 6.3 × 105 (a) |
| o-Cymene | 5.1 × 107 ± 2.4 × 106 (c) | 2.6 × 107 ± 6.3 × 106 (bc) | 5.1 × 107 ± 4.9 × 106 (c) | 3.5 × 107 ± 5.1 × 106 (bc) | 4.6 × 105 ± 2.0 × 104 (a) | 4.6 × 105 ± 4.9 × 104 (a) | 3.9 × 106 ± 6.2 × 105 (b) | 2.7 × 106 ± 6.7 × 105 (ab) |
| p-Cymene | nd | nd | nd | 7.5 × 105 ± 3.4 × 104 (a) | nd | nd | 4.6 × 105 ± 1.7 × 104 (a) | 7.9 × 105 ± 1.7 × 104 (a) |
| m-Cymene | nd | nd | 7.1 × 105 ± 1.7 × 104 (a) | 4.9 × 105 ± 4.0 × 104 (a) | nd | nd | 4.6 × 105 ± 5.0 × 104 (a) | 5.7 × 105 ± 5.6 × 104 (a) |
| β-Phellandrene | 3.7 × 108 ± 5.9 × 107 (ab) | 2.3 × 108 ± 1.3 × 107 (ab) | 2.9 × 108 ± 2.1 × 107 (a) | 1.2 × 108 ± 1.5 × 107 (ab) | 9.7 × 108 ± 1.9 × 107 (c) | 6.3 × 108 ± 3.4 × 107 (bc) | 2.5 × 108 ± 2.4 × 107 (ab) | 3.3 × 108 ± 2.4 × 107 (abc) |
| β-Ocimene | 8.1 × 105 ± 3.9 × 104 (b) | 7.1 × 105 ± 3.3 × 104 (b) | 1.7 × 106 ± 1.9 × 105 (c) | nd | 2.5 × 105 ± 2.0 × 104 (a) | 4.6 × 105 ± 2.1 × 104 (ab) | 1.3 × 106 ± 2.3 × 105 (bc) | nd |
| α-Ocimene | nd | nd | nd | 4.9 × 106 ± 4.9 × 105 (b) | nd | nd | nd | 4.6 × 105 ± 1.8 × 104 (a) |
| γ-Terpinene | 9.4 × 106 ± 3.7 × 105 (c) | 5.7 × 106 ± 2.2 × 105 (bc) | 9.4 × 106 ± 5.2 × 105 (c) | 6.0 × 106 ± 4.2 × 105 (b) | 5.4 × 106 ± 2.7 × 105 (bc) | 3.6 × 106 ± 1.4 × 105 (b) | 4.9 × 105 ± 2.0 × 104 (a) | 1.8 × 106 ± 1.6 × 105 (ab) |
| Cryptone | 1.2 × 106 ± 3.5 × 105 (a) | nd | 6.1 × 105 ± 1.3 × 104 (a) | nd | nd | nd | nd | 2.4 × 104 ± 2.1 × 103 (a) |
| Terpinolene | 1.6 × 106 ± 4.3 × 105 (a) | 1.3 × 106 ± 2.4 × 105 (a) | 2.9 × 106 ± 3.2 × 105 (a) | 1.3 × 106 ± 1.9 × 105 (a) | 4.6 × 104 ± 3.2 × 103 (a) | 4.6 × 106 ± 6.1 × 105 (a) | 3.2 × 105 ± 2.4 × 104 (a) | 2.3 × 107 ± 5.8 × 106 (b) |
| β-Cyclocitral | 7.8 × 105 ± 8.8 × 104 (b) | 9.9 × 105 ± 5.9 × 104 (bc) | 1.0 × 106 ± 9.6 × 104 (bc) | 8.2 × 105 ± 1.3 × 105 (b) | 8.9 × 105 ± 2.9 × 105 (b) | 1.7 × 106 ± 8.3 × 105 (c) | 1.2 × 105 ± 2.0 × 105 (a) | 1.4 × 106 ± 7.0 × 105 (bc) |
| δ-Elemene | 6.9 × 105 ± 3.3 × 104 (a) | 1.2 × 106 ± 2.9 × 105 (a) | nd | 1.4 × 106 ± 1.3 × 105 (a) | 1.3 × 104 ± 7.3 × 102 (a) | 1.7 × 105 ± 4.3 × 104 (a) | nd | 3.5 × 107 ± 4.7 × 106 (b) |
| α-Copaene | nd | 1.2 × 106 ± 2.6 × 105 (c) | nd | nd | 2.2 × 105 ± 1.7 × 104 (a) | 6.6 × 105 ± 1.0 × 104 (bc) | 4.7 × 105 ± 6.4 × 104 (b) | 5.6 × 105 ± 5.7 × 104 (bc) |
| Isocaryophyllene | 1.7 × 107 ± 1.7 × 106 (b) | 2.1 × 106 ± 1.6 × 105 (a) | 4.4 × 106 ± 2.8 × 105 (a) | 5.8 × 106 ± 3.9 × 105 (a) | 3.2 × 106 ± 3.4 × 105 (a) | 5.4 × 106 ± 1.4 × 105 (a) | 2.1 × 106 ± 1.8 × 105 (a) | 4.0 × 107 ± 2.6 × 106 (b) |
| α-Caryophyllene | 3.9 × 106 ± 6.7 × 105 (ab) | 8.8 × 106 ± 4.9 × 105 (b) | 4.5 × 106 ± 3.7 × 105 (ab) | 8.4 × 106 ± 1.7 × 105 (b) | nd | 9.4 × 105 ± 9.2 × 104 (a) | nd | nd |
| β-lonone | 1.0 × 106 ± 1.7 × 105 (a) | 1.1 × 106 ± 9.9 × 104 (a) | 1.3 × 106 ± 2.4 × 105 (a) | 1.2 × 106 ± 7.2 × 104 (a) | 4.7 × 105 ± 4.6 × 104 (a) | 4.7 × 105 ± 4.4 × 104 (a) | 1.3 × 106 ± 4.9 × 105 (a) | 7.9 × 106 ± 3.3 × 105 (b) |
| Other VOCs | ||||||||
| Acetic acid | nd | nd | nd | nd | nd | nd | 7.3 × 105 ± 3.9 × 104 (a) | 2.3 × 106 ± 3.9 × 105 (a) |
| Hexane | 3.5 × 106 ± 1.7 × 105 (c) | 8.4 × 105 ± 3.8 × 104 (b) | 2.0 × 105 ± 2.9 × 104 (a) | 1.2 × 106 ± 1.4 × 105 (b) | nd | nd | nd | 3.0 × 105 ± 2.1 × 104 (a) |
| cis-1,2-Dimethyl-cyclopentane | nd | nd | nd | 6.0 × 106 ± 7.3 × 105 (bc) | 1.3 × 106 ± 3.4 × 105 (ab) | 2.3 × 106 ± 3.8 × 105 (b) | 3.3 × 105 ± 4.7 × 104 (a) | 8.4 × 106 ± 5.2 × 105 (c) |
| Naphthalene | 8.9 × 105 ± 5.1 × 104 (b) | nd | 9.7 × 105 ± 4.5 × 104 (b) | 8.9 × 105 ± 4.2 × 104 (b) | nd | nd | 2.2 × 105 ± 2.4 × 104 (a) | 6.4 × 105 ± 5.2 × 104 (ab) |
| Azulene | Nd | 2.0 × 106 ± 5.4 × 105 (b) | nd | nd | nd | 1.6 × 105 ± 6.4 × 104 (a) | nd | 1.4 × 105 ± 4.8 × 104 (a) |
| 2-Methyltetrahydrofuran | nd | 1.0 × 106 ± 5.4 × 105 (b) | nd | nd | nd | 1.2 × 105 ± 2.5 × 104 (a) | nd | 1.3 × 105 ± 4.2 × 104 (a) |
| 2-Ethylfuran | 7.2 × 106 ± 3.7 × 105 (b) | 8.9 × 106 ± 8.9 × 105 (bc) | 1.4 × 107 ± 4.1 × 106 (c) | 9.9 × 106 ± 4.4 × 105 (bc) | 1.2 × 106 ± 3.0 × 105 (a) | 7.8 × 104 ± 1.2 × 104 (a) | 3.3 × 105 ± 1.8 × 104 (a) | 3.5 × 106 ± 6.6 × 105 (b) |
| 2-Pentylfuran | 1.8 × 106 ± 4.3 × 105 (bc) | 2.9 × 106 ± 7.1 × 105 (c) | 1.7 × 106 ± 2.1 × 105 (bc) | 2.3 × 106 ± 3.7 × 105 (bc) | 1.2 × 105 ± 2.4 × 104 (a) | 1.6 × 106 ± 2.6 × 105 (b) | 1.1 × 106 ± 8.1 × 104 (ab) | 1.7 × 106 ± 1.7 × 105 (b) |
| 2-Ethylthiophene | 1.2 × 108 ± 2.6 × 107 (a) | 1.1 × 108 ± 9.3 × 106 (a) | nd | 1.4 × 108 ± 3.5 × 107 (a) | nd | nd | nd | nd |
| Benzofuran | 1.8 × 106 ± 1.5 × 105 (b) | 2.6 × 106 ± 4.9 × 105 (b) | 1.7 × 106 ± 7.3 × 105 (b) | 1.7 × 106 ± 4.5 × 105 (b) | 1.4 × 105 ± 8.4 × 103 (a) | 1.2 × 106 ± 3.5 × 105 (b) | 2.1 × 106 ± 1.9 × 105 (b) | 3.2 × 105 ± 3.5 × 104 (ab) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawrocka, J.; Szymczak, K.; Skwarek-Fadecka, M.; Małolepsza, U. Toward the Analysis of Volatile Organic Compounds from Tomato Plants (Solanum lycopersicum L.) Treated with Trichoderma virens or/and Botrytis cinerea. Cells 2023, 12, 1271. https://doi.org/10.3390/cells12091271
Nawrocka J, Szymczak K, Skwarek-Fadecka M, Małolepsza U. Toward the Analysis of Volatile Organic Compounds from Tomato Plants (Solanum lycopersicum L.) Treated with Trichoderma virens or/and Botrytis cinerea. Cells. 2023; 12(9):1271. https://doi.org/10.3390/cells12091271
Chicago/Turabian StyleNawrocka, Justyna, Kamil Szymczak, Monika Skwarek-Fadecka, and Urszula Małolepsza. 2023. "Toward the Analysis of Volatile Organic Compounds from Tomato Plants (Solanum lycopersicum L.) Treated with Trichoderma virens or/and Botrytis cinerea" Cells 12, no. 9: 1271. https://doi.org/10.3390/cells12091271