
Canine Intestinal Organoids as a Novel In Vitro Model of Intestinal Drug Permeability: A Proof-of-Concept Study

 , ,
, ,  , ,
, , 
Abstract
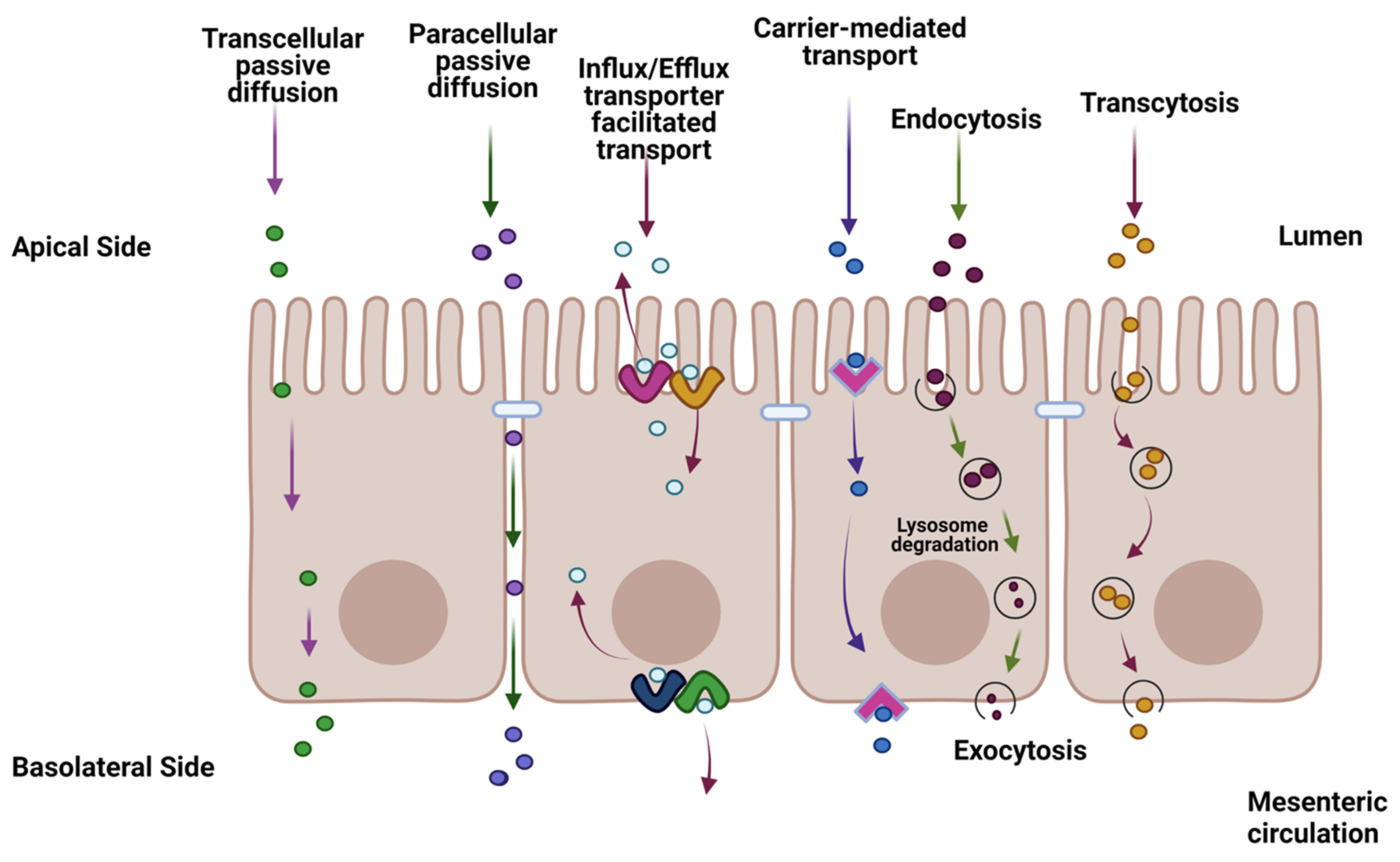
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Caco-2 Cell Culture and Maintenance
2.3. Caco-2 Two-Dimensional Monolayer Preparation and Maintenance
2.4. Maintenance of 3D Colonoids
2.5. Preparation of 2D Canine Colonic Monolayer and Maintenance
2.6. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction Analyses
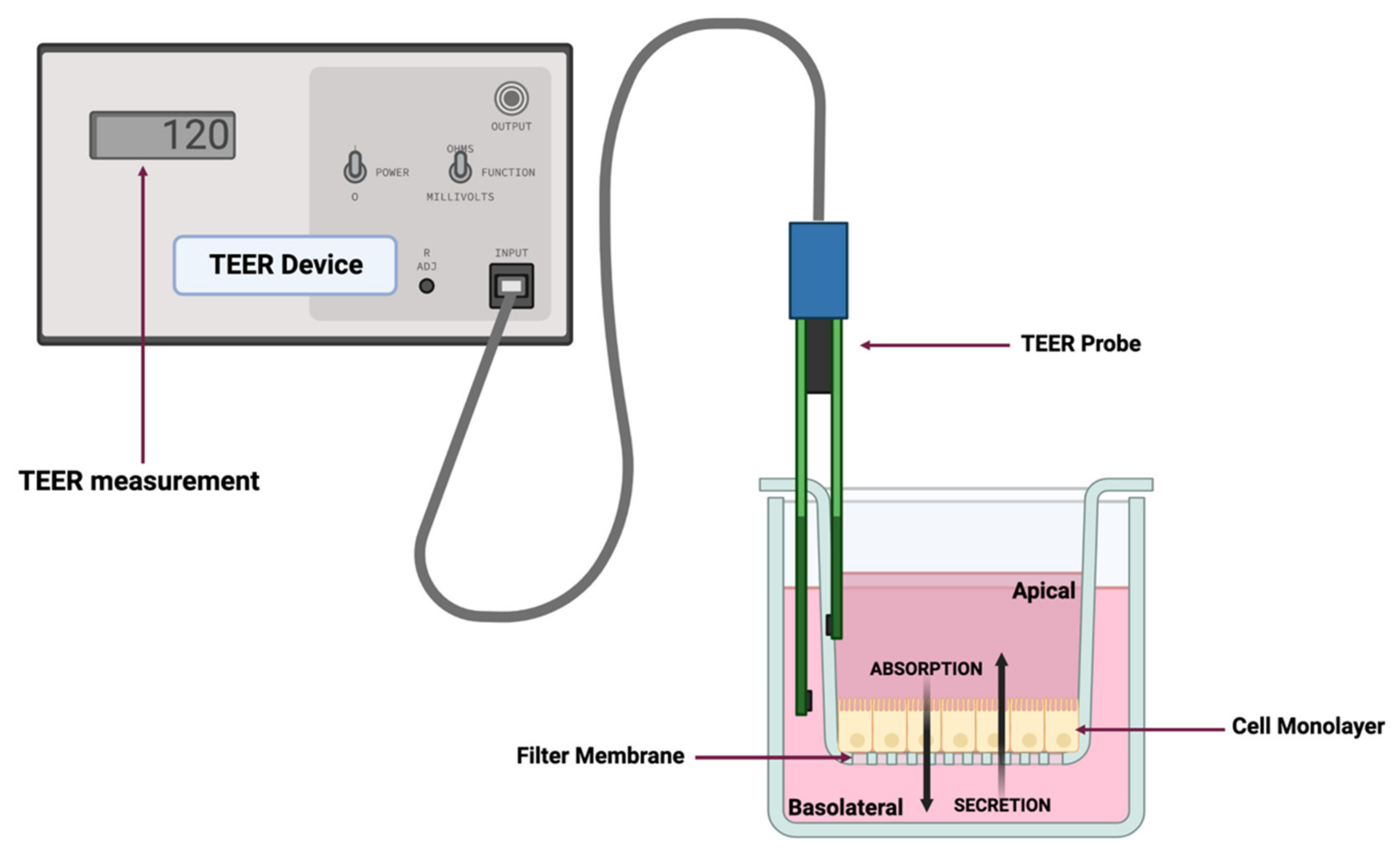
2.7. Assessment of Monolayer Integrity by TEER Measurement and Fluorescein Isothiocyanate-Dextran Leakage Assay
2.8. Preparation of Transport Buffer and Drug Solutions
2.9. Bidirectional Transport Experiments
2.9.1. Apical-to-Basolateral (AP→BL) Permeability
2.9.2. Basolateral-to-Apical (BL→AP) Permeability
2.10. Sample Preparation for Mass Spectrometer Analysis
2.10.1. Preparation of Standards and Solutions
2.10.2. Analytical Method
2.11. Papp Calculations
2.12. Statistics
3. Results
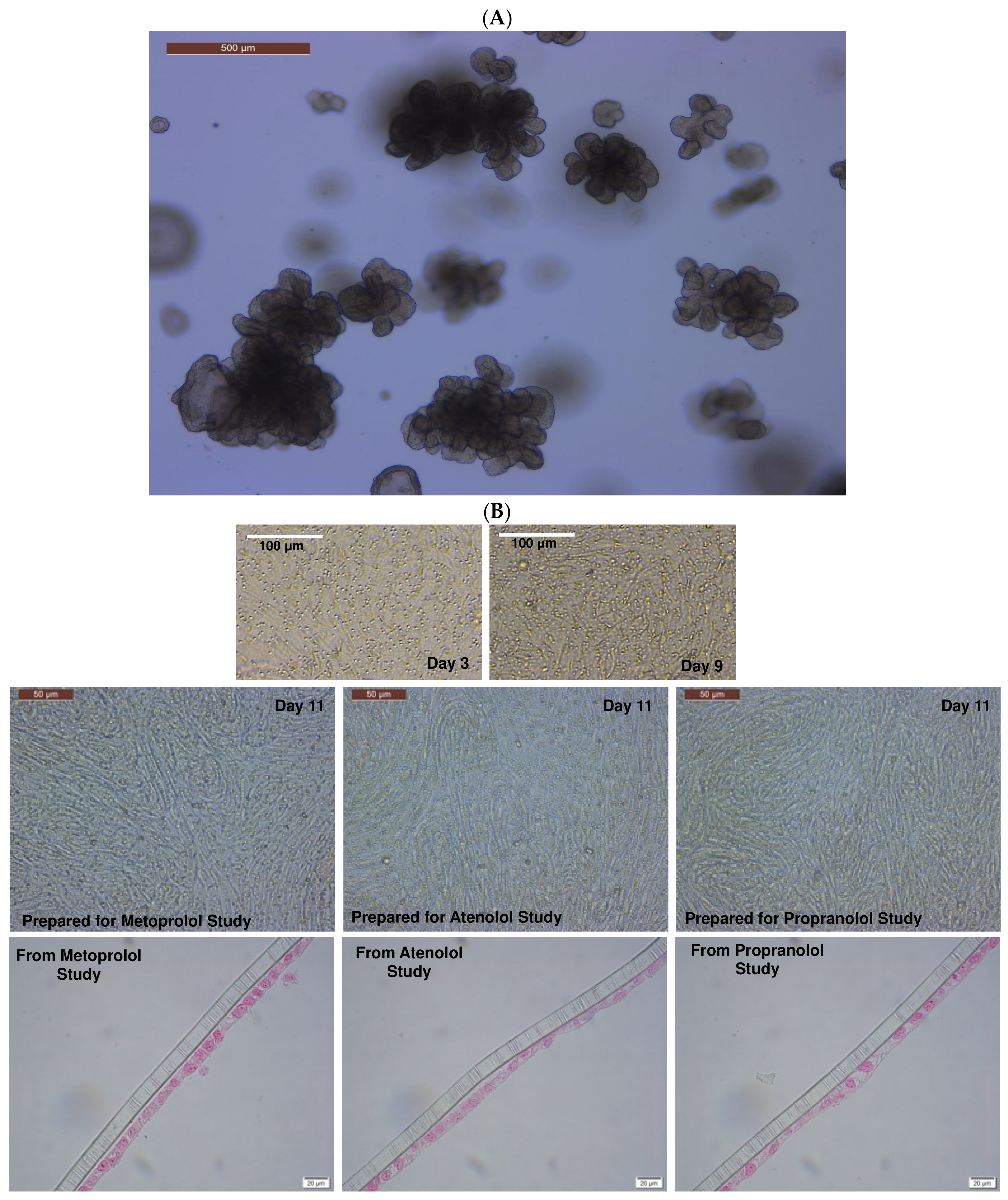
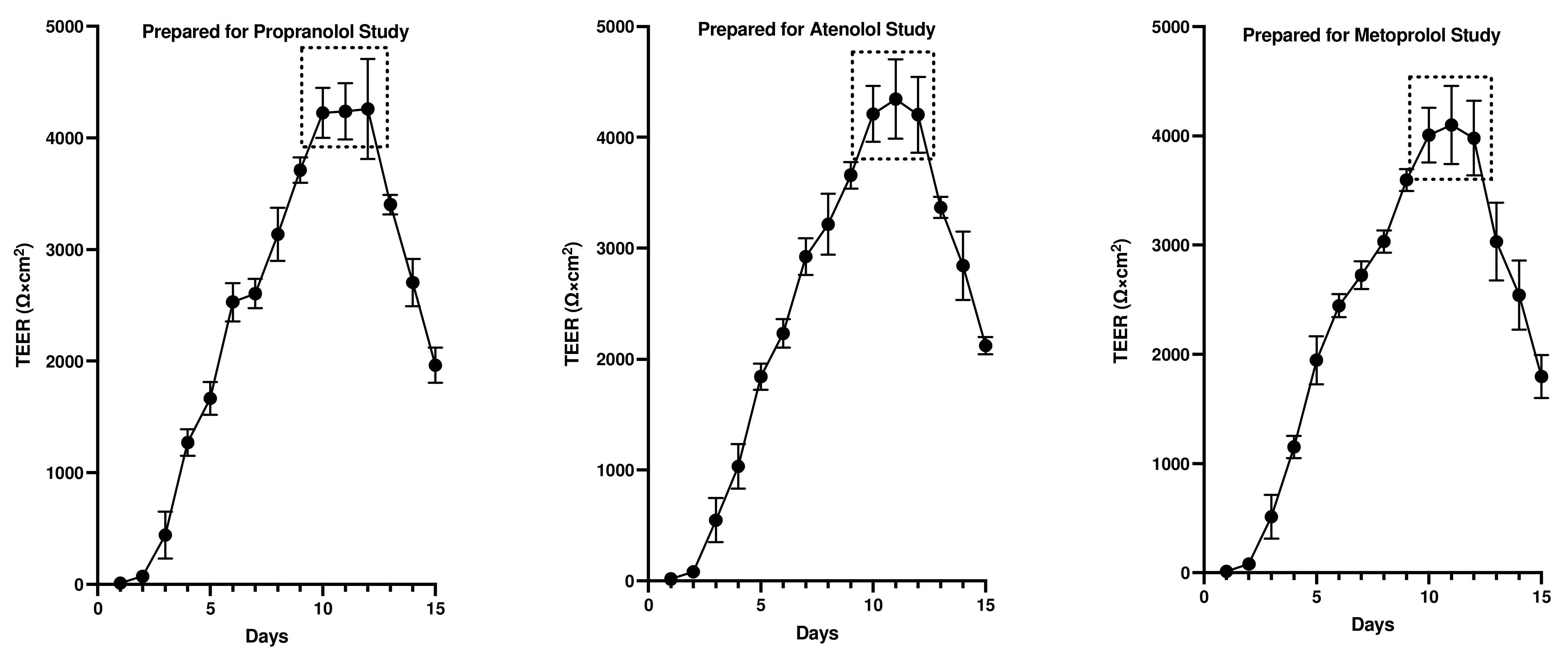
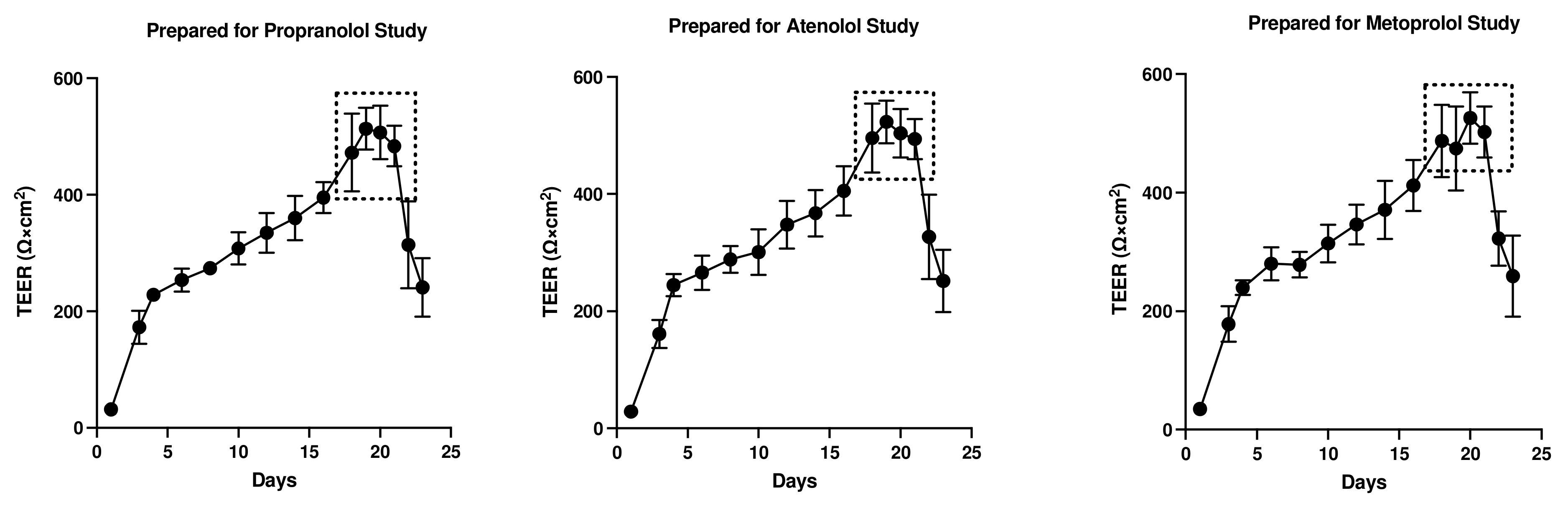
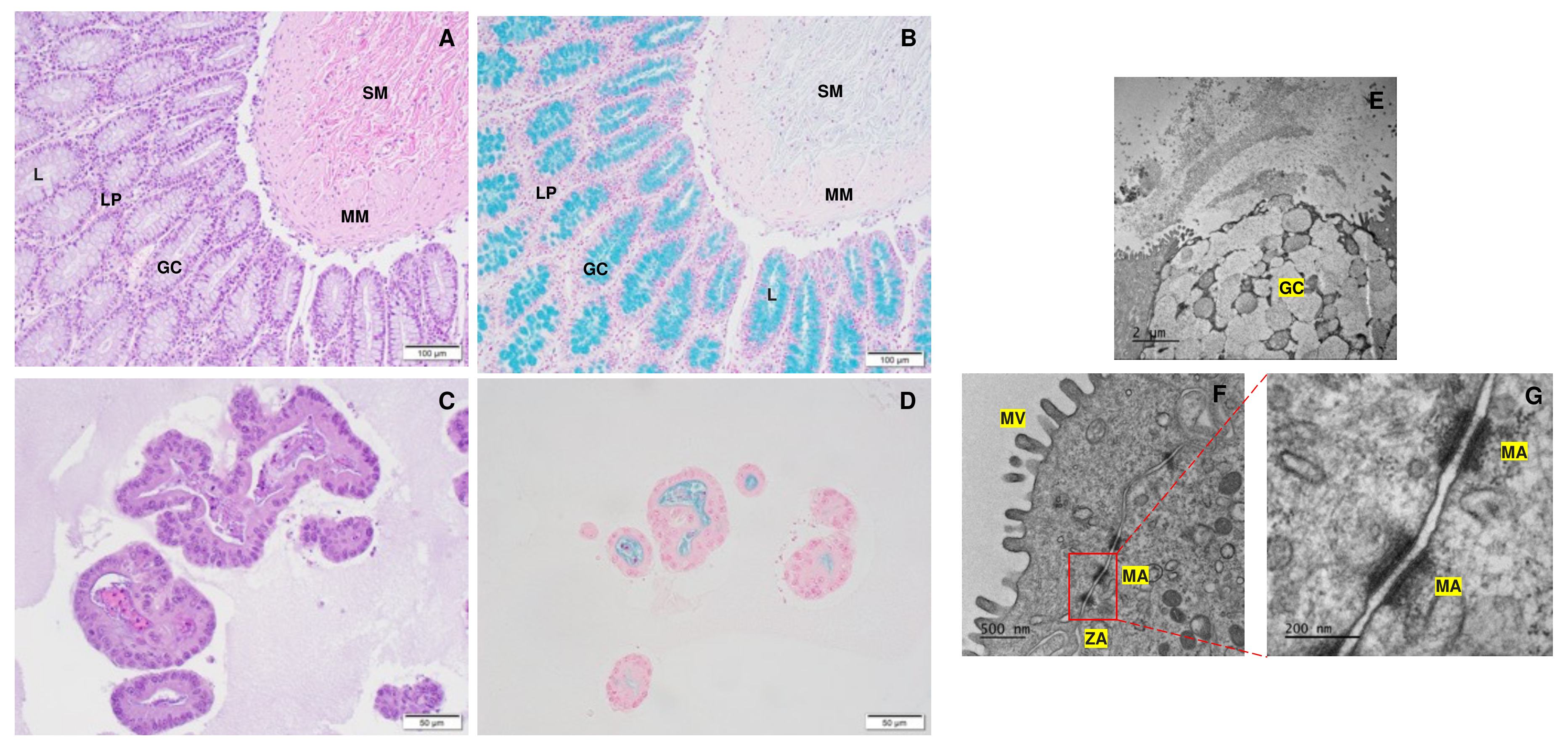
3.1. Assessment of Monolayer Integrity
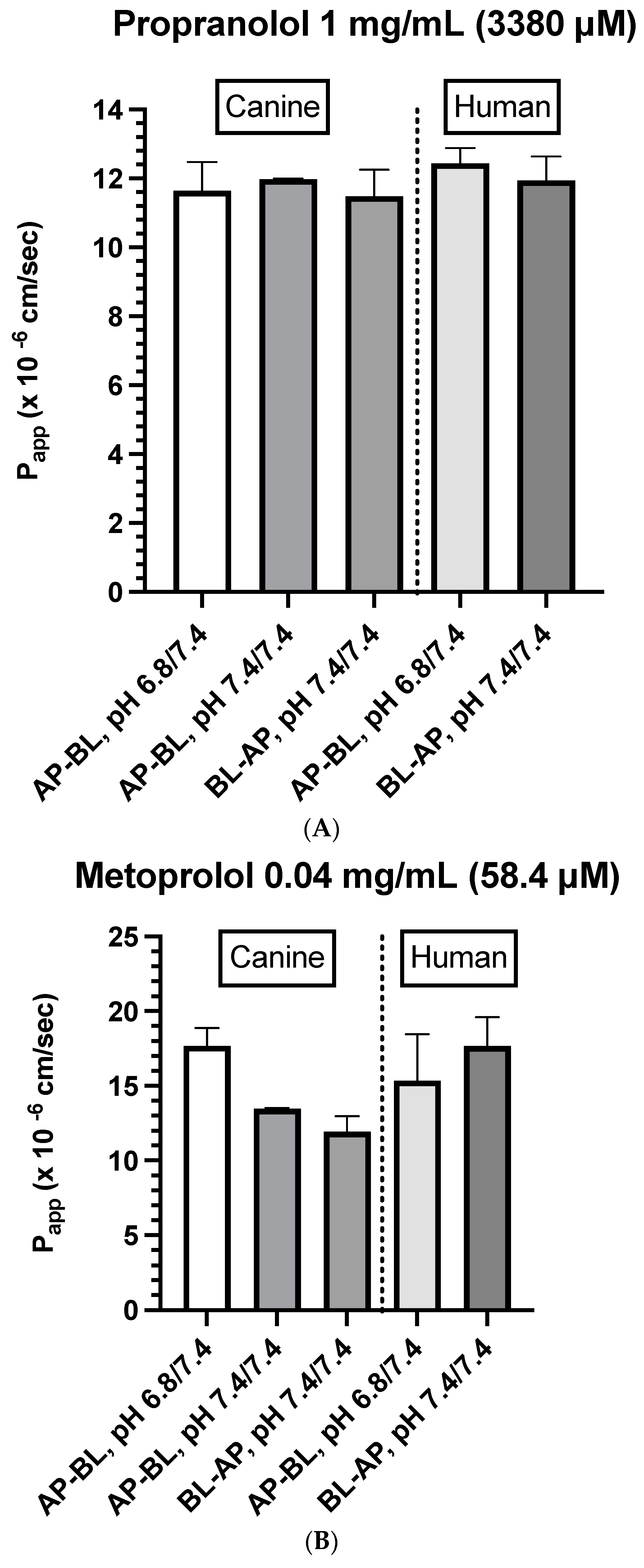
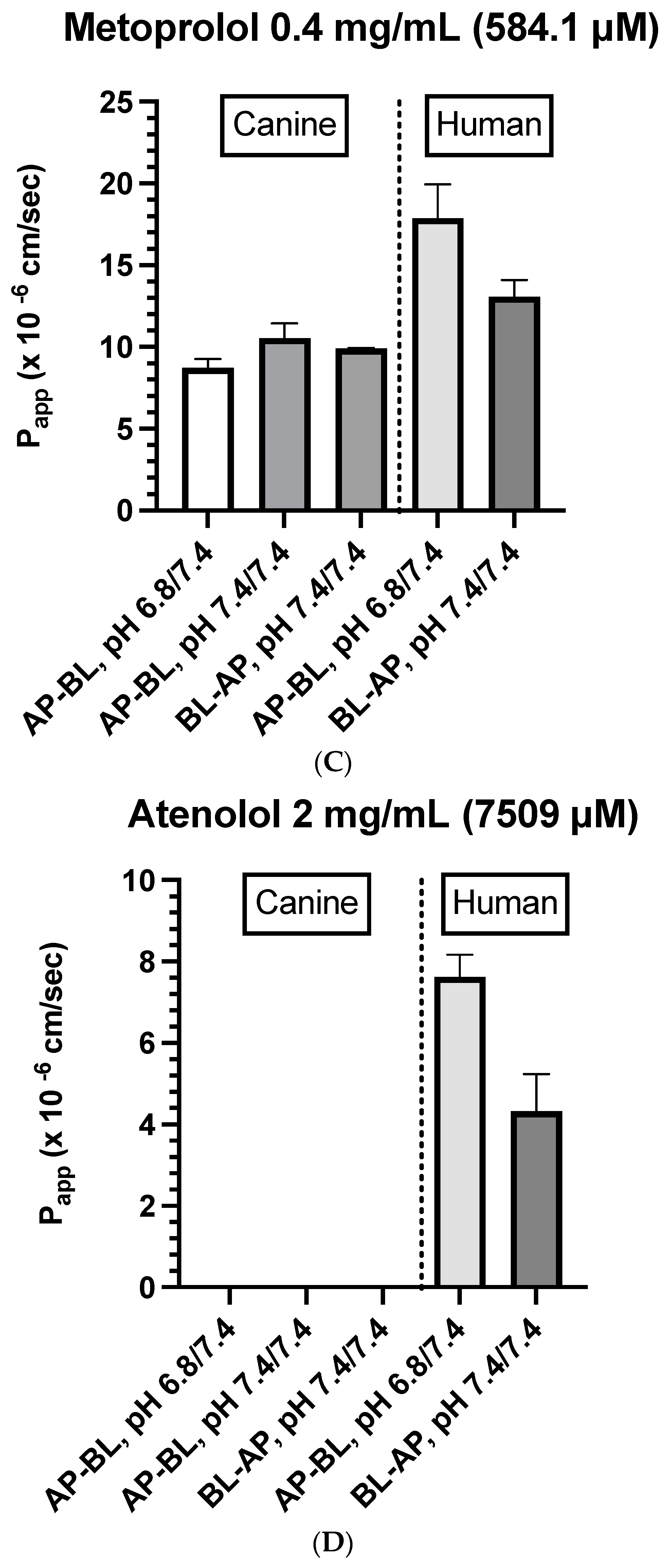
3.2. Drug-Specific Papp Estimates
- Canine colonoids: The AP→BL Papp values were lower in the medium containing 0.4 vs. 0.04 mg/mL metoprolol. Moreover, at the 0.04 mg/mL concentration, the movement from AP→BL was slightly greater than that from BL→AP. The transport from AP→BL and BL→AP tended to be similar when evaluated at the 0.4 mg/mL concentration. Upon considering the data generated across the two metoprolol concentrations, the pH of the apical chamber did not consistently influence the magnitude of the Papp estimate (Figure 6B,C and Table 5).
- Caco-2: Unlike canine colonoids, the Papp values were not markedly influenced by metoprolol concentration, and at both concentrations, the movements from AP→BL and BL→AP were comparable (Figure 6B,C and Table 5). The concentration-associated differences in the Papp values seen with the colonoid were not observed with the Caco-2 monolayer. Moreover, although the two cell line monolayers exhibited similar AP→BL Papp values in the presence of 0.04 mg/mL metoprolol, the AP→BL Papp values for the Caco-2 monolayer tended to be higher than that of the canine colonoid when the donor concentration was increased to 0.4 mg/mL. Although the movement from AP→BL was somewhat greater than that seen in the BL→AP direction in the colonoid (0.04 mg/mL metoprolol, but not at the 0.4 mg/mL donor concentration), that difference was not seen with the 0.04 mg/mL concentration or was only minimally appreciated at the 0.4 mg/mL concentration when the Caco-2 monolayer was used. When considering the variability across observations and the small number of wells tested, statistical inferences should not be linked to these outcomes (Figure 6C,D and Table 5). Thus, unlike propranolol, differences in the behavior of metoprolol were seen when comparing the two cell line systems. However, these preliminary findings should be interpreted cautiously in light of our limited sample size and the background variability in our system.
- Caco-2: Incubation of Caco-2 cells with 2.0 mg/mL atenolol resulted in measurable concentrations in the receiver compartment for both AP→BL and BL→AP directions, with higher Papp estimates, reported after incubation in the apical chamber (Figure 6D and Table 5). These preliminary findings suggest a somewhat greater ability for atenolol to undergo paracellular transport across the Caco-2 monolayer as compared to that of the dog colonoid. However, at the lower atenolol concentration, neither system was associated with quantifiable movement from the donor to receiver compartment (irrespective of direction).
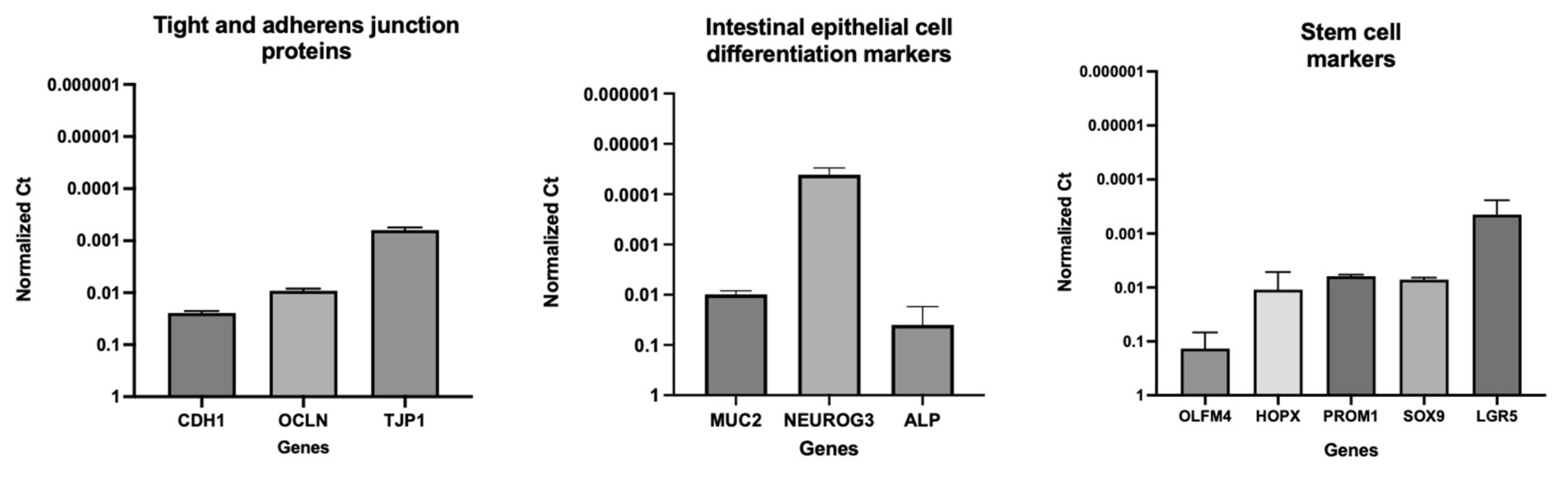
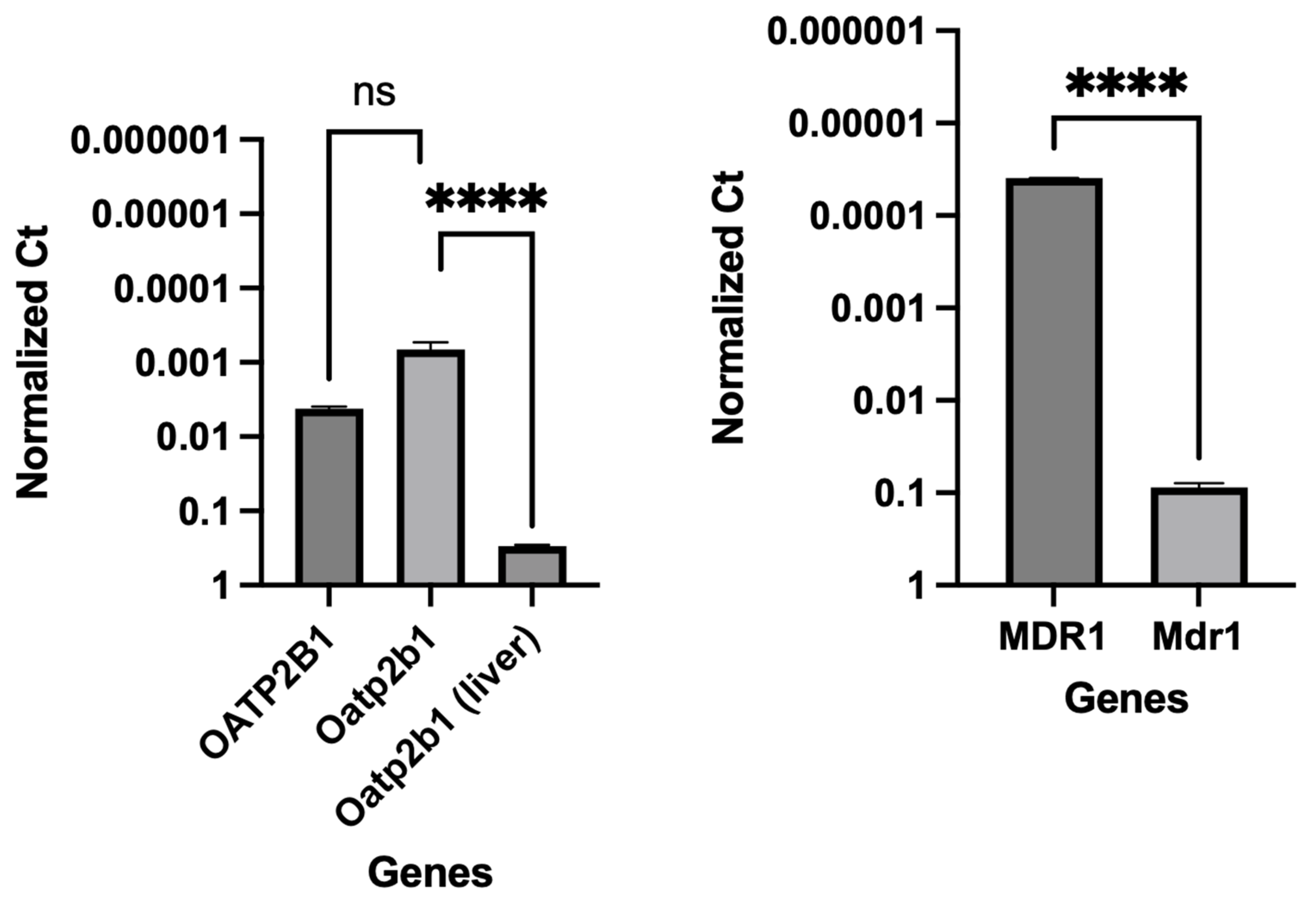
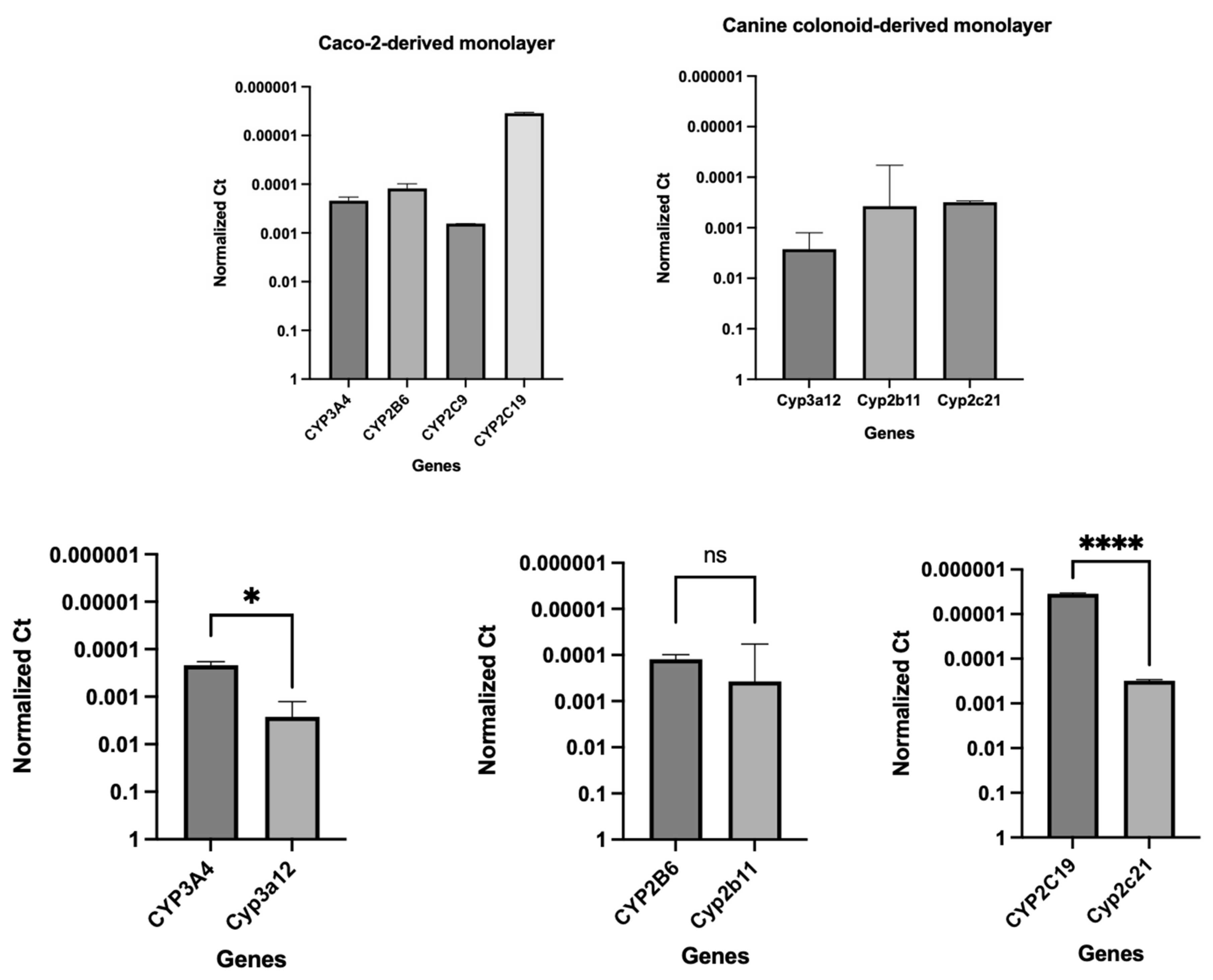
3.3. Gene Expression Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alqahtani, M.S.; Kazi, M.; Alsenaidy, M.A.; Ahmad, M.Z. Advances in Oral Drug Delivery. Front. Pharmacol. 2021, 12, 62. [Google Scholar] [CrossRef] [PubMed]
- Sugano, K.; Kansy, M.; Artursson, P.; Avdeef, A.; Bendels, S.; Di, L.; Ecker, G.F.; Faller, B.; Fischer, H.; Gerebtzoff, G.; et al. Coexistence of Passive and Carrier-Mediated Processes in Drug Transport. Nat. Rev. Drug Discov. 2010, 9, 597–614. [Google Scholar] [CrossRef] [PubMed]
- Roos, C. The Impact of Regional Permeability, Nanoparticles, and Absorption-Modifying Excipients. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 2018. [Google Scholar]
- Furuse, M. Molecular Basis of the Core Structure of Tight Junctions. Cold Spring Harb. Perspect. Biol. 2010, 2, a002907. [Google Scholar] [CrossRef] [PubMed]
- Fleth-James, J. Suitability of in Vitro, in Silico and in Vivo Methods to Predict Intestinal Absorption in Drug Development. Ph.D. Thesis, Freie Universität Berlin, Berlin, Germany, 2018. [Google Scholar] [CrossRef]
- Teksin, Z.S.; Seo, P.R.; Polli, J.E. Comparison of Drug Permeabilities and BCS Classification: Three Lipid-Component PAMPA System Method versus Caco-2 Monolayers. AAPS J. 2010, 12, 238. [Google Scholar] [CrossRef]
- Volpe, D.A. Variability in Caco-2 and MDCK Cell-Based Intestinal Permeability Assays. J. Pharm. Sci. 2008, 97, 712–725. [Google Scholar] [CrossRef]
- Volpe, D.A. Drug-Permeability and Transporter Assays in Caco-2 and MDCK Cell Lines. Future Med. Chem. 2011, 3, 2063–2077. [Google Scholar] [CrossRef]
- Volpe, D.A. Advances in Cell-Based Permeability Assays to Screen Drugs for Intestinal Absorption. Expert Opin. Drug Discov. 2020, 15, 539–549. [Google Scholar] [CrossRef]
- Volpe, D.A. Application of Method Suitability for Drug Permeability Classification. AAPS J. 2010, 12, 670–678. [Google Scholar] [CrossRef]
- Hill, D.R.; Huang, S.; Tsai, Y.H.; Spence, J.R.; Young, V.B. Real-Time Measurement of Epithelial Barrier Permeability in Human Intestinal Organoids. J. Vis. Exp. 2017, 2017, e56960. [Google Scholar] [CrossRef]
- Pade, D.; Jamei, M.; Rostami-Hodjegan, A.; Turner, D.B. Application of the MechPeff Model to Predict Passive Effective Intestinal Permeability in the Different Regions of the Rodent Small Intestine and Colon. Biopharm. Drug Dispos. 2017, 38, 94–114. [Google Scholar] [CrossRef]
- Macedo, M.H.; Martinez, E.; Barrias, C.C.; Sarmento, B. Development of an Improved 3D in Vitro Intestinal Model to Perform Permeability Studies of Paracellular Compounds. Front. Bioeng. Biotechnol. 2020, 8, 1076. [Google Scholar] [CrossRef]
- Lea, T. Caco-2 Cell Line. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland, 2015; pp. 103–111. [Google Scholar] [CrossRef]
- Tavelin, S.; Gråsjö, J.; Taipalensuu, J.; Ocklind, G.; Artursson, P. Applications of Epithelial Cell Culture in Studies of Drug Transport. Methods Mol. Biol. 2002, 188, 233–272. [Google Scholar] [CrossRef]
- Sai, Y.; Kaneko, Y.; Ito, S.; Mitsuoka, K.; Kato, Y.; Tamai, I.; Artursson, P.; Tsuji, A. Predominant Contribution of Organic Anion Transporting Polypeptide OATP-B (OATP2B1) to Apical Uptake of Estrone-3-Sulfate by Human Intestinal Caco-2 Cells. Drug Metab. Dispos. 2006, 34, 1423–1431. [Google Scholar] [CrossRef]
- Balimane, P.V.; Chong, S. Cell Culture-Based Models for Intestinal Permeability: A Critique. Drug Discov. Today 2005, 10, 335–343. [Google Scholar] [CrossRef]
- Ozawa, T.; Takayama, K.; Okamoto, R.; Negoro, R.; Sakurai, F.; Tachibana, M.; Kawabata, K.; Mizuguchi, H. Generation of Enterocyte-like Cells from Human Induced Pluripotent Stem Cells for Drug Absorption and Metabolism Studies in Human Small Intestine. Sci. Rep. 2015, 5, 16479. [Google Scholar] [CrossRef]
- Mochel, J.P.; Jergens, A.E.; Kingsbury, D.; Kim, H.J.; Martín, M.G.; Allenspach, K. Intestinal Stem Cells to Advance Drug Development, Precision, and Regenerative Medicine: A Paradigm Shift in Translational Research. AAPS J. 2018, 20, 17. [Google Scholar] [CrossRef]
- Maubon, N.; Le Vee, M.; Fossati, L.; Audry, M.; Le Ferrec, E.; Bolze, S.; Fardel, O. Analysis of Drug Transporter Expression in Human Intestinal Caco-2 Cells by Real-Time PCR. Fundam. Clin. Pharmacol. 2007, 21, 659–663. [Google Scholar] [CrossRef]
- Neuhoff, S.; Ungell, A.L.; Zamora, I.; Artursson, P. PH-Dependent Passive and Active Transport of Acidic Drugs across Caco-2 Cell Monolayers. Eur. J. Pharm. Sci. 2005, 25, 211–220. [Google Scholar] [CrossRef]
- Yamashita, T.; Inui, T.; Yokota, J.; Kawakami, K.; Morinaga, G.; Takatani, M.; Hirayama, D.; Nomoto, R.; Ito, K.; Cui, Y.; et al. Monolayer Platform Using Human Biopsy-Derived Duodenal Organoids for Pharmaceutical Research. Mol. Ther. Methods Clin. Dev. 2021, 22, 263–278. [Google Scholar] [CrossRef]
- Ito, K.; Suzuki, H.; Horie, T.; Sugiyama, Y. Apical/Basolateral Surface Expression of Drug Transporters and Its Role in Vectorial Drug Transport. Pharm. Res. 2005, 22, 1559–1577. [Google Scholar] [CrossRef]
- Hayeshi, R.; Hilgendorf, C.; Artursson, P.; Augustijns, P.; Brodin, B.; Dehertogh, P.; Fisher, K.; Fossati, L.; Hovenkamp, E.; Korjamo, T.; et al. Comparison of Drug Transporter Gene Expression and Functionality in Caco-2 Cells from 10 Different Laboratories. Eur. J. Pharm. Sci. 2008, 35, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; Zucco, F.; Felsani, A. Good Caco-2 Cell Culture Practices. Toxicol. Vitr. 2012, 26, 1243–1246. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.; Burmester, M.; Langeheine, M.; Brehm, R.; Empl, M.T.; Seeger, B.; Breves, G. Caco-2/HT29-MTX Co-Cultured Cells as a Model for Studying Physiological Properties and Toxin-Induced Effects on Intestinal Cells. PLoS ONE 2021, 16, e0257824. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chow, E.C.Y.; Liu, S.; Du, Y.; Pang, K.S. The Caco-2 Cell Monolayer: Usefulness and Limitations. Expert Opin. Drug Metab. Toxicol. 2008, 4, 395–411. [Google Scholar] [CrossRef]
- Thummel, K.E.; Brimer, C.; Yasuda, K.; Thottassery, J.; Senn, T.; Lin, Y.; Ishizuka, H.; Kharasch, E.; Schuetz, J.; Schuetz, E. Transcriptional Control of Intestinal Cytochrome P-4503A by 1α,25-Dihydroxy Vitamin D3. Mol. Pharmacol. 2001, 60, 1399–1406. [Google Scholar] [CrossRef]
- Minkler, S.; Lucien, F.; Kimber, M.J.; Sahoo, D.K.; Bourgois-Mochel, A.; Musser, M.; Johannes, C.; Frank, I.; Cheville, J.; Allenspach, K.; et al. Emerging Roles of Urine-Derived Components for the Management of Bladder Cancer: One Man’s Trash Is Another Man’s Treasure. Cancers 2021, 13, 422. [Google Scholar] [CrossRef]
- Shanahan, M.T.; Kanke, M.; Oyesola, O.O.; Hung, Y.H.; Koch-Laskowski, K.; Singh, A.P.; Peck, B.C.E.; Biraud, M.; Sheahan, B.; Cortes, J.E.; et al. Multiomic Analysis Defines the First MicroRNA Atlas across All Small Intestinal Epithelial Lineages and Reveals Novel Markers of Almost All Major Cell Types. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G668–G681. [Google Scholar] [CrossRef]
- Kopper, J.J.; Iennarella-Servantez, C.; Jergens, A.E.; Sahoo, D.K.; Guillot, E.; Bourgois-Mochel, A.; Martinez, M.N.; Allenspach, K.; Mochel, J.P. Harnessing the Biology of Canine Intestinal Organoids to Heighten Understanding of Inflammatory Bowel Disease Pathogenesis and Accelerate Drug Discovery: A One Health Approach. Front. Toxicol. 2021, 3, 773953. [Google Scholar] [CrossRef]
- Gabriel, V.; Zdyrski, C.; Sahoo, D.K.; Dao, K.; Bourgois-Mochel, A.; Kopper, J.; Zeng, X.-L.; Estes, M.K.; Mochel, J.P.; Allenspach, K. Standardization and Maintenance of 3D Canine Hepatic and Intestinal Organoid Cultures for Use in Biomedical Research. J. Vis. Exp. 2022, 179, e63515. [Google Scholar] [CrossRef]
- Zdyrski, C.; Iennarella-Servantez, C.A.; Sahoo, D.K.; Ward, J.; Long, E.; Gabriel, V.; Minkler, S.; Mao, S.; Bourgois-Mochel, A.; Jergens, A.; et al. Su124 homology directed repair in canine duodenal enteroids to mimic the wild-type p-glycoprotein mutation. Gastroenterology 2021, 160, S-625. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 Stem Cells Build Crypt-Villus Structures in Vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Chandra, L.; Borcherding, D.C.; Kingsbury, D.; Atherly, T.; Ambrosini, Y.M.; Bourgois-Mochel, A.; Yuan, W.; Kimber, M.; Qi, Y.; Wang, Q.; et al. Derivation of Adult Canine Intestinal Organoids for Translational Research in Gastroenterology. BMC Biol. 2019, 17, 33. [Google Scholar] [CrossRef]
- Ambrosini, Y.M.; Park, Y.; Jergens, A.E.; Shin, W.; Min, S.; Atherly, T.; Borcherding, D.C.; Jang, J.; Allenspach, K.; Mochel, J.P.; et al. Recapitulation of the Accessible Interface of Biopsy-Derived Canine Intestinal Organoids to Study Epithelial-Luminal Interactions. PLoS ONE 2020, 15, e0231423. [Google Scholar] [CrossRef]
- Gabriel, V.; Zdyrski, C.; Sahoo, D.K.; Dao, K.; Bourgois-Mochel, A.; Atherly, T.; Martinez, M.N.; Volpe, D.A.; Kopper, J.; Allenspach, K.; et al. Canine Intestinal Organoids in a Dual-Chamber Permeable Support System. J. Vis. Exp. 2022, 2022, e63612. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Borcherding, D.C.; Chandra, L.; Jergens, A.E.; Atherly, T.; Bourgois-Mochel, A.; Ellinwood, N.M.; Snella, E.; Severin, A.J.; Martin, M.; et al. Differential Transcriptomic Profiles Following Stimulation with Lipopolysaccharide in Intestinal Organoids from Dogs with Inflammatory Bowel Disease and Intestinal Mast Cell Tumor. Cancers 2022, 14, 3525. [Google Scholar] [CrossRef]
- Bedos, L.; Wickham, H.; Gabriel, V.; Zdyrski, C.; Allbaugh, R.A.; Sahoo, D.K.; Sebbag, L.; Mochel, J.P.; Allenspach, K. Culture and Characterization of Canine and Feline Corneal Epithelial Organoids: A New Tool for the Study and Treatment of Corneal Diseases. Front. Vet. Sci. 2022, 9, 1050467. [Google Scholar] [CrossRef]
- Kodama, N.; Iwao, T.; Katano, T.; Ohta, K.; Yuasa, H.; Matsunaga, T. Characteristic Analysis of Intestinal Transport in Enterocyte-Like Cells Differentiated from Human Induced Pluripotent Stem Cells. Drug Metab. Dispos. 2016, 44, 1662–1667. [Google Scholar] [CrossRef]
- Akazawa, T.; Yoshida, S.; Ohnishi, S.; Kanazu, T.; Kawai, M.; Takahashi, K. Application of Intestinal Epithelial Cells Differentiated from Human Induced Pluripotent Stem Cells for Studies of Prodrug Hydrolysis and Drug Absorption in the Small Intestine. Drug Metab. Dispos. 2018, 46, 1497–1506. [Google Scholar] [CrossRef]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocr. Rev. 2009, 30, 204–213. [Google Scholar] [CrossRef]
- Ergün, S.; Wörsdörfer, P. Organoids, Assembloids and Embryoids: New Avenues for Developmental Biology, Disease Modeling, Drug Testing and Toxicity Assessment without Animal Experimentation. Organoids 2022, 1, 37–40. [Google Scholar] [CrossRef]
- Liebau, S.; Parvin, B.; Ergün, S.; Richiardone, E.; Van Den Bossche, V.; Corbet, C. Metabolic Studies in Organoids: Current Applications, Opportunities and Challenges. Organoids 2022, 1, 85–105. [Google Scholar] [CrossRef]
- Burgess, T.; Barker, N.; Torresi, J.; Munro, M.J.; Tan, S.T.; Gray, C. Applications for Colon Organoid Models in Cancer Research. Organoids 2023, 2, 37–49. [Google Scholar] [CrossRef]
- Schutgens, F.; Rookmaaker, M.B.; Margaritis, T.; Rios, A.; Ammerlaan, C.; Jansen, J.; Gijzen, L.; Vormann, M.; Vonk, A.; Viveen, M.; et al. Tubuloids Derived from Human Adult Kidney and Urine for Personalized Disease Modeling. Nat. Biotechnol. 2019, 37, 303–313. [Google Scholar] [CrossRef]
- Usui, T.; Sakurai, M.; Nishikawa, S.; Umata, K.; Nemoto, Y.; Haraguchi, T.; Itamoto, K.; Mizuno, T.; Noguchi, S.; Mori, T.; et al. Establishment of a Dog Primary Prostate Cancer Organoid Using the Urine Cancer Stem Cells. Cancer Sci. 2017, 108, 2383–2392. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Ding, B.; Wan, M.; Chen, L.; Jackson, J.; Atala, A. Formation and Optimization of Three-Dimensional Organoids Generated from Urine-Derived Stem Cells for Renal Function in Vitro. Stem Cell Res. Ther. 2020, 11, 309. [Google Scholar] [CrossRef]
- Zhang, Y.; McNeill, E.; Tian, H.; Soker, S.; Andersson, K.E.; Yoo, J.J.; Atala, A. Urine Derived Cells Are a Potential Source for Urological Tissue Reconstruction. J. Urol. 2008, 180, 2226–2233. [Google Scholar] [CrossRef]
- Yu, F.; Hunziker, W.; Choudhury, D. Engineering Microfluidic Organoid-on-a-Chip Platforms. Micromachines 2019, 10, 165. [Google Scholar] [CrossRef]
- Danku, A.E.; Dulf, E.H.; Braicu, C.; Jurj, A.; Berindan-Neagoe, I. Organ-On-A-Chip: A Survey of Technical Results and Problems. Front. Bioeng. Biotechnol. 2022, 10, 94. [Google Scholar] [CrossRef]
- van Berlo, D.; Nguyen, V.V.T.; Gkouzioti, V.; Leineweber, K.; Verhaar, M.C.; van Balkom, B.W.M. Stem Cells, Organoids, and Organ-on-a-Chip Models for Personalized in Vitro Drug Testing. Curr. Opin. Toxicol. 2021, 28, 7–14. [Google Scholar] [CrossRef]
- Menon-Andersen, D.; Florian, J.; Madabushi, R. Clinical Pharmacology Review NDA Number 205410 Submission Type; Code: Seattle, WA, USA, 2013. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2014/205410Orig1s000ClinPharmR.pdf (accessed on 11 January 2023).
- Yang, Y.; Faustino, P.J.; Volpe, D.A.; Ellison, C.D.; Lyon, R.C.; Yu, L.X. Biopharmaceutics Classification of Selected β-Blockers: Solubility and Permeability Class Membership. Mol. Pharm. 2007, 4, 608–614. [Google Scholar] [CrossRef]
- Martinez, M.N.; Mochel, J.P.; Neuhoff, S.; Pade, D. Comparison of Canine and Human Physiological Factors: Understanding Interspecies Differences That Impact Drug Pharmacokinetics. AAPS J. 2021, 23, 59. [Google Scholar] [CrossRef]
- Volpe, D.A.; Faustino, P.J.; Ciavarella, A.B.; Asafu-Adjaye, E.B.; Ellison, C.D.; Yu, L.X.; Hussain, A.S. Classification of Drug Permeability with a Caco-2 Cell Monolayer Assay. Ceased 2008, 24, 39–47. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Roy, A.; Bhanja, S.; Chainy, G.B.N. Hypothyroidism Impairs Antioxidant Defence System and Testicular Physiology during Development and Maturation. Gen. Comp. Endocrinol. 2008, 156, 63–70. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Roy, A.; Chainy, G.B.N. Protective Effects of Vitamin E and Curcumin on L-Thyroxine-Induced Rat Testicular Oxidative Stress. Chem. Biol. Interact. 2008, 176, 121–128. [Google Scholar] [CrossRef]
- Zdyrski, C.; Gabriel, V.; Ospina, O.; Wickham, H.; Sahoo, D.K.; Dao, K.; Aguilar Meza, L.S.; Bedos, L.; Honold, S.; Pineyro, P.; et al. Establishment and Characterization of Novel Canine Organoids with Organ-Specific Physiological Similarity. bioRxiv. 2022, 2022-07, e63612. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, X.; Hu, B.; Xing, M.; Cao, Q.; Ji, A.; Song, S. Transport Mechanisms of Polymannuronic Acid and Polyguluronic Acid Across Caco-2 Cell Monolayers. Pharmaceutics 2020, 12, 167. [Google Scholar] [CrossRef]
- Stockdale, T.P.; Challinor, V.L.; Lehmann, R.P.; de Voss, J.J.; Blanchfield, J.T. Caco-2 Monolayer Permeability and Stability of Chamaelirium Luteum (False Unicorn) Open-Chain Steroidal Saponins. ACS Omega 2019, 4, 7658–7666. [Google Scholar] [CrossRef]
- Frost, T.S.; Jiang, L.; Lynch, R.M.; Zohar, Y. Permeability of Epithelial/Endothelial Barriers in Transwells and Microfluidic Bilayer Devices. Micromachines 2019, 10, 533. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 22 December 2022).
- Falavigna, M.; Stein, P.C.; Flaten, G.E.; di Cagno, M.P. Impact of Mucin on Drug Diffusion: Development of a Straightforward In Vitro Method for the Determination of Drug Diffusivity in the Presence of Mucin. Pharmaceutics 2020, 12, 168. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, U.D.; Mahalingam, R.; Li, X.; Pather, I.; Jasti, B. Effect of Experimental Temperature on the Permeation of Model Diffusants Across Porcine Buccal Mucosa. AAPS PharmSciTech 2011, 12, 579. [Google Scholar] [CrossRef] [PubMed]
- Flaten, G.E.; Dhanikula, A.B.; Luthman, K.; Brandl, M. Drug Permeability across a Phospholipid Vesicle Based Barrier: A Novel Approach for Studying Passive Diffusion. Eur. J. Pharm. Sci. 2006, 27, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Caco-2 Permeability Assay. Available online: https://www.cyprotex.com/admepk/in-vitro-permeability/caco-2-permeability (accessed on 13 March 2022).
- US Food and Drug Administration. In Vitro Drug Interaction Studies—Cytochrome P450 Enzyme- and Transporter-Mediated Drug Interactions. Guidance for Industry. 2020. Available online: https://www.fda.gov/media/134581/download (accessed on 30 March 2023).
- Katneni, K.; Pham, T.; Saunders, J.; Chen, G.; Patil, R.; White, K.L.; Abla, N.; Chiu, F.C.K.; Shackleford, D.M.; Charman, S.A. Using Human Plasma as an Assay Medium in Caco-2 Studies Improves Mass Balance for Lipophilic Compounds. Pharm. Res. 2018, 35, 210. [Google Scholar] [CrossRef]
- Heinlein, A.; Metzger, M.; Walles, H.; Buettner, A. Transport of Hop Aroma Compounds across Caco-2 Monolayers. Food Funct. 2014, 5, 2719. [Google Scholar] [CrossRef]
- Quan, Y.; Jin, Y.; Faria, T.N.; Tilford, C.A.; He, A.; Wall, D.A.; Smith, R.L.; Vig, B.S. Expression Profile of Drug and Nutrient Absorption Related Genes in Madin-Darby Canine Kidney (MDCK) Cells Grown under Differentiation Conditions. Pharmaceutics 2012, 4, 314–333. [Google Scholar] [CrossRef]
- Almeida, L.D.; Quaglio, A.E.V.; De Almeida Costa, C.A.R.; Di Stasi, L.C. Intestinal Anti-Inflammatory Activity of Ground Cherry (Physalis angulata L.) Standardized CO2 Phytopharmaceutical Preparation. World J. Gastroenterol. 2017, 23, 4369–4380. [Google Scholar] [CrossRef]
- Youhanna, S.; Lauschke, V.M. The Past, Present and Future of Intestinal In Vitro Cell Systems for Drug Absorption Studies. J. Pharm. Sci. 2021, 110, 50–65. [Google Scholar] [CrossRef]
- He, Y.L.; Murby, S.; Warhurst, G.; Gifford, L.; Walker, D.; Ayrton, J.; Eastmond, R.; Rowland, M. Species Differences in Size Discrimination in the Paracellular Pathway Reflected by Oral Bioavailability of Poly(Ethylene Glycol) and D-Peptides. J. Pharm. Sci. 1998, 87, 626–633. [Google Scholar] [CrossRef]
- Incecayir, T.; Tsume, Y.; Amidon, G.L. Comparison of the Permeability of Metoprolol and Labetalol in Rat, Mouse, and Caco-2 Cells: Use as a Reference Standard for BCS Classification. Mol. Pharm. 2013, 10, 958–966. [Google Scholar] [CrossRef]
- Zur, M.; Gasparini, M.; Wolk, O.; Amidon, G.L.; Dahan, A. The Low/High BCS Permeability Class Boundary: Physicochemical Comparison of Metoprolol and Labetalol. Mol. Pharm. 2014, 11, 1707–1714. [Google Scholar] [CrossRef]
- Dahlgren, D.; Roos, C.; Sjögren, E.; Lennernäs, H. Direct In Vivo Human Intestinal Permeability (Peff) Determined with Different Clinical Perfusion and Intubation Methods. J. Pharm. Sci. 2015, 104, 2702–2726. [Google Scholar] [CrossRef]
- Dahlgren, D.; Roos, C.; Johansson, P.; Lundqvist, A.; Tannergren, C.; Abrahamsson, B.; Sjögren, E.; Lennernäs, H. Regional Intestinal Permeability in Dogs: Biopharmaceutical Aspects for Development of Oral Modified-Release Dosage Forms. Mol. Pharm. 2016, 13, 3022–3033. [Google Scholar] [CrossRef]
- Machen, T.E.; Erlij, D.; Wooding, F.B.P. PERMEABLE JUNCTIONAL COMPLEXES: The Movement of Lanthanum across Rabbit Gallbladder and Intestine. J. Cell Biol. 1972, 54, 302. [Google Scholar] [CrossRef]
- Frömter, E.; Diamond, J. Route of Passive Ion Permeation in Epithelia. Nat. New Biol. 1972, 235, 9–13. [Google Scholar] [CrossRef]
- Monaco, A.; Ovryn, B.; Axis, J.; Amsler, K. The Epithelial Cell Leak Pathway. Int. J. Mol. Sci. 2021, 22, 7677. [Google Scholar] [CrossRef]
- Amidon, G.; Lee, P.; Topp, E. Transport Processes in Pharmaceutical Systems; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Saaby, L.; Helms, H.C.C.; Brodin, B. IPEC-J2 MDR1, a Novel High-Resistance Cell Line with Functional Expression of Human P-Glycoprotein (ABCB1) for Drug Screening Studies. Mol. Pharm. 2016, 13, 640–652. [Google Scholar] [CrossRef]
- Dahlgren, D.; Lennernäs, H. Intestinal Permeability and Drug Absorption: Predictive Experimental, Computational and In Vivo Approaches. Pharmaceutics 2019, 11, 411. [Google Scholar] [CrossRef]
- Li, J.; Volpe, D.A.; Wang, Y.; Zhang, W.; Bode, C.; Owen, A.; Hidalgo, I.J. Use of Transporter Knockdown Caco-2 Cells to Investigate the In Vitro Efflux of Statin Drugs. Drug Metab. Dispos. 2011, 39, 1196–1202. [Google Scholar] [CrossRef]
- Meneses, A.M.C.; Schneeberger, K.; Kruitwagen, H.S.; Penning, L.C.; van Steenbeek, F.G.; Burgener, I.A.; Spee, B. Intestinal Organoids—Current and Future Applications. Vet. Sci. 2016, 3, 31. [Google Scholar] [CrossRef]
- Reineking, W.; Schauerte, I.E.; Junginger, J.; Hewicker-Trautwein, M. Sox9, Hopx, and Survivin and Tuft Cell Marker DCLK1 Expression in Normal Canine Intestine and in Intestinal Adenoma and Adenocarcinoma. Vet. Pathol. 2022, 59, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Karim, B.O.; Rhee, K.J.; Liu, G.; Yun, K.; Brant, S.R. Prom1 Function in Development, Intestinal Inflammation, and Intestinal Tumorigenesis. Front. Oncol. 2014, 4, 323. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Chen, S.H.; Zhang, Y.N.; Xu, C.F. Olfactomedin-4 in Digestive Diseases: A Mini-Review. World J. Gastroenterol. 2018, 24, 1881. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.B.; Hwangbo, S.; Jang, S.; Jo, Y.K. Bioengineered Co-Culture of Organoids to Recapitulate Host-Microbe Interactions. Mater. Today Bio 2022, 16, 100345. [Google Scholar] [CrossRef]
- Ahn, J.S.; Kang, M.J.; Seo, Y.; Kim, H.S. Intestinal Organoids as Advanced Modeling Platforms to Study the Role of Host-Microbiome Interaction in Homeostasis and Disease. BMB Rep. 2023, 56, 15–23. [Google Scholar] [CrossRef]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the Dog and Human Gut Microbiomes in Gene Content and Response to Diet. Microbiome 2018, 6, 72. [Google Scholar] [CrossRef]
- Sawant-Basak, A.; Rodrigues, A.D.; Lech, M.; Doyonnas, R.; Kasaian, M.; Prasad, B.; Tsamandouras, N. Physiologically Relevant, Humanized Intestinal Systems to Study Metabolism and Transport of Small Molecule Therapeutics. Drug Metab. Dispos. 2018, 46, 1581–1587. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Category | Gene Full Name | Symbol | Forward Primer Sequence (5′–3′) | Reverse Primer Sequence (5′–3′) |
|---|---|---|---|---|---|
| Canis lupus familiaris (dog) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | TCAACGGATTTGGCCGTATTGG | TGAAGGGGTCATTGATGGCG |
| Tight and adherens junction proteins | Cadherin 1 | CDH1 | GACCCAGTAACTAACGACG | CTTCATTCACATCTTCCACG | |
| Occludin | OCLN | CACTACTGTGTGGTGGATCC | CCTTGTCCCACAATATATTCG | ||
| Tight junction protein 1 | TJP1 | GAGGGTGATCAAATTCTCAGG | CTGATTCTACAATGCGACG | ||
| Intestinal epithelial cell differentiation markers | Mucin 2 | MUC2 | CCTGTGCCCCATATTCTGC | GAGATGTTGGAATGGATGCC | |
| Neurogenin 3 | NEUROG3 | GAATGCACAACCTCAACTCG | GTAGAGGCTGTGGTCCGC | ||
| Intestinal alkaline phosphatase | ALP | CGTAGTAAACCGCAACTGG | GGAAACATGTACTTTCGGC | ||
| Stem cell markers | Olfactomedin 4 | OLFM4 | GTATCATGAATGTCAGCAAGC | CTGTAATATTCCAGAATTCTTCC | |
| HOP homeobox | HOPX | GACCAGGTGGAGATTCTGG | GCCAGACGCTGCTTAAACC | ||
| Prominin 1 | PROM1 | GATTATTATTTGTGCTGTCC | GAGACTGTAAAGTATTTCCTC | ||
| SRY-box transcription factor 9 | SOX9 | GTCATCTCCAACATAGAGACC | CTGCTTGGACATCCACACG | ||
| Leucine-rich repeat-containing G-protein-coupled receptor 5 | LGR5 | GCTAGATCTGTCTTACAACC | GTTCCAGGCTAAATTCAGC | ||
| Transporters | Organic anion transporting polypeptide | Oatp2b1 | GATGACTTTGCCCACAACAGC | CAGCAGCAGAGATGAGGAAGC | |
| Multidrug resistance p-glycoprotein | Mdr1 | GTAGCTGAAGAAGTCTTAGCAGC | GCGGCACCAATAGAAATGTTGGC | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P-450 3a12 | Cyp3a12 | GATCATGAACATGAAACTTGC | CTTTTCAGGTTGAATAATCCC | |
| Cytochrome P450 2b11 | Cyp2b11 | CTGAGGGAGTCCTCCAGGACCC | CACATAGAACAAGTTCATCAGG | ||
| Cytochrome P450 2C21 | Cyp2c21 (Cyp2c18) | CAAGCACCTCCTGGATACAGC | CTTCGTGTTCTTTTATTTTTTCC | ||
| Homo sapiens (human) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | TGCACCACCAACTGCTTAGC | GGCATGGACTGTGGTCATGAG |
| Transporters | Organic anion transporting polypeptide 2B1/solute carrier organic anion transporter family member 2B1 | OATP2B1 (SLCO2B1) | CAAACCTGACTGTGATCCAG | GAGCAGGTTGGCGTATGAGG | |
| ATP binding cassette subfamily B member 1 | ABCB1 (MDR1) | CAGTAGCTGAAGAGGTCTTGGC | CTGTAATAGCTTTCTTTATCCC | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P450 family 3 subfamily A member 4 | CYP3A4 | GAGATGGTCCCTATCATTGCC | GATGTTCACTCCAAATGATGTGC | |
| Cytochrome P450 family 2 subfamily B member 6 | CYP2B6 | GAAACCGCTGGAAGGTGCTTCG | CTCCTCTATCAGACACTGAGC | ||
| Cytochrome P450 family 2 subfamily C member 9 | CYP2C9 | GAAGGAGATCCGGCGTTTCTCC | CTTGGTTTTTCTCAACTCCTCC | ||
| Cytochrome P450 family 2 subfamily C member 19 | CYP2C19 | GATCTGCTCCATTATTTTCC | GTTTTTAAGTAATTTGTTATGG |
| Drug | Molecular Formula (MF) | Molecular Weight (MW) | Aqueous Solubility (25 °C) | Log P | Dissociation Constants (Basic pKa) |
|---|---|---|---|---|---|
| Metoprolol | C15H25NO3 | 267.36 g/mol | >1000 mg/mL | 2.15 | 9.56 |
| Atenolol | C14H22N2O3 | 266.34 g/mol | 13.3 mg/mL | 0.16 | 9.58 |
| Propranolol | C16H21NO2 | 259.339 g/mol | 0.0617 mg/L | 3.48 | 9.53 |
| Compound Name | pH | QC Level (ppm) | Average Measured Value (ppm) | Bias (%) |
|---|---|---|---|---|
| Atenolol | 6.8 | 25 | 23.1 | −7.41 |
| 7.4 | 25 | 23.8 | −4.60 | |
| Metoprolol | 6.8 | 25 | 25.5 | 1.93 |
| 7.4 | 25 | 24.9 | −0.22 | |
| Propranolol | 6.8 | 40 | 40.2 | 0.61 |
| 7.4 | 40 | 40.9 | 2.33 |
| Compound | Time (min) | Donor (%) | Receiver (%) |
|---|---|---|---|
| FITC-Dextran (200 µg/mL) | 0 | 100 ± 0.0 | 0.020 ± 0.001 |
| 15 | 0.021 ± 0.003 | ||
| 30 | 0.024 ± 0.004 | ||
| 45 | 0.022 ± 0.002 | ||
| 60 | 0.024 ± 0.003 | ||
| 90 | 0.023 ± 0.004 | ||
| 120 | 93.7 ± 7.8 | 0.022 ± 0.002 | |
| Metoprolol (0.4 mg/mL) + | 0 | 100 ± 0.0 | 0.021 ± 0.001 |
| FITC-Dextran (200 µg/mL) | 15 | 0.023 ± 0.001 | |
| 30 | 0.020 ± 0.00 | ||
| 45 | 0.020 ± 0.00 | ||
| 60 | 0.022 ± 0.001 | ||
| 90 | 0.023 ± 0.00 | ||
| 120 | 94.6 ± 1.4 | 0.020 ± 0.001 |
| Drug | Species | Direction | Well | Papp × 10−6 | Avg | SD | %CV |
|---|---|---|---|---|---|---|---|
| Metoprolol (0.4 mg/mL or 584.1 μM) | Human | AP→BL pH 6.8/7.4 | 1 | 19.34 | |||
| 2 | 16.45 | 17.89 | 2.05 | 11.43 | |||
| BL→AP pH 7.4/7.4 | 1 | 13.79 | |||||
| 2 | 12.37 | 13.08 | 1.01 | 7.72 | |||
| Dog * | AP→BL pH 6.8/7.4 | 1 | 8.33 | ||||
| 2 | 9.10 | 8.72 | 0.55 | 6.32 | |||
| AP→BL pH 7.4/7.4 | 1 | 11.19 | |||||
| 2 | 9.90 | 10.54 | 0.91 | 8.66 | |||
| BL→AP pH 7.4/7.4 | 1 | 9.88 | |||||
| 2 | 9.94 | 9.91 | 0.05 | 0.46 | |||
| Metoprolol (0.04 mg/mL or 58.4 μM) | Human | AP→BL pH 6.8/7.4 | 1 | 17.54 | |||
| 2 | 13.13 | 15.33 | 3.12 | 20.38 | |||
| BL→AP pH 7.4/7.4 | 1 | 16.30 | |||||
| 2 | 19.03 | 17.67 | 1.93 | 10.92 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | 16.83 | ||||
| 2 | 18.52 | 17.68 | 1.19 | 6.75 | |||
| AP→BL pH 7.4/7.4 | 1 | 13.44 | |||||
| 2 | 13.50 | 13.47 | 0.04 | 0.32 | |||
| BL→AP pH 7.4/7.4 | 1 | 11.19 | |||||
| 2 | 12.67 | 11.93 | 1.04 | 8.76 | |||
| Atenolol (0.2 mg/mL or 750.9 μM) | Human | AP→BL pH 6.8/7.4 | 1 | BLQ | |||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Dog | AP→BL pH 6.8/7.4 | 1 | BLQ | ||||
| 2 | BLQ | ||||||
| AP→BL pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Atenolol (2 mg/mL or 7509 μM) | Human | AP→BL pH 6.8/7.4 | 1 | 7.25 | |||
| 2 | 8.01 | 7.63 | 0.54 | 7.09 | |||
| BL→AP pH 7.4/7.4 | 1 | 3.69 | |||||
| 2 | 4.97 | 4.33 | 0.90 | 20.89 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | BLQ | ||||
| 2 | BLQ | ||||||
| AP→BL pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Propranolol (1 mg/mL or 3380 μM) | Human | AP→BL pH 6.8/7.4 | 1 | 12.12 | |||
| 2 | 12.75 | 12.44 | 0.45 | 3.59 | |||
| BL→AP pH 7.4/7.4 | 1 | 11.45 | |||||
| 2 | 12.43 | 11.94 | 0.70 | 5.84 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | 12.23 | ||||
| 2 | 11.05 | 11.64 | 0.83 | 7.15 | |||
| AP→BL pH 7.4/7.4 | 1 | 11.99 | |||||
| 2 | 11.97 | 11.98 | 0.01 | 0.09 | |||
| BL→AP pH 7.4/7.4 | 1 | 10.94 | |||||
| 2 | 12.03 | 11.49 | 0.77 | 6.70 |
| Species | Category | Gene Full Name | Symbol | Cycle Threshold (Ct) | |
|---|---|---|---|---|---|
| Mean | SD | ||||
| Canis lupus familiaris (dog) (colonoid-derived monolayer) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | 20.67 | 1.52 |
| Tight and adherens junction proteins | Cadherin 1 | CDH1 | 24.49 | 0.12 | |
| Occludin | OCLN | 25.92 | 0.12 | ||
| Tight junction protein 1 | TJP1 | 29.80 | 0.15 | ||
| Intestinal epithelial cell differentiation markers | Mucin 2 | MUC2 | 25.83 | 0.22 | |
| Neurogenin 3 | NEUROG3 | 33.75 | 0.37 | ||
| Intestinal alkaline phosphatase | ALP | 26.91 | 0.76 | ||
| Stem cell markers | Olfactomedin 4 | OLFM4 | 22.19 | 0.91 | |
| HOP homeobox | HOPX | 25.85 | 1.01 | ||
| Prominin 1 | PROM1 | 26.48 | 0.10 | ||
| SRY-box transcription factor 9 | SOX9 | 26.27 | 0.12 | ||
| Leucine-rich repeat-containing G-protein-coupled receptor 5 | LGR5 | 30.38 | 0.68 | ||
| Transporters | Organic anion transporting polypeptide | Oatp2b1 | 32.41 | 0.28 | |
| Oatp2b1 (liver tissue) (used as positive control) | 21.41 | 0.07 | |||
| Multidrug resistance p-glycoprotein | Mdr1 | 25.36 | 0.14 | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P-450 3a12 | Cyp3a12 | 27.40 | 0.70 | |
| Cytochrome P450 2b11 | Cyp2b11 | 30.48 | 1.29 | ||
| Cytochrome P450 2C21 | Cyp2c21 (Cyp2c18) | 31.24 | 0.08 | ||
| Homo sapiens (human) (Caco-2-derived monolayer) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | 20.21 | 0.08 |
| Transporters | Organic anion transporting polypeptide 2B1/solute carrier organic anion transporter family member 2B1 | OATP2B1 (SLCO2B1) | 28.11 | 0.10 | |
| ATP binding cassette subfamily B member 1 | ABCB1 (MDR1) | 35.53 | 1.76 | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P450 family 3 subfamily A member 4 | CYP3A4 | 32.39 | 0.22 | |
| Cytochrome P450 family 2 subfamily B member 6 | CYP2B6 | 33.22 | 0.30 | ||
| Cytochrome P450 family 2 subfamily C member 9 | CYP2C9 | 31.34 | 1.46 | ||
| Cytochrome P450 family 2 subfamily C member 19 | CYP2C19 | 38.34 | 0.06 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahoo, D.K.; Martinez, M.N.; Dao, K.; Gabriel, V.; Zdyrski, C.; Jergens, A.E.; Atherly, T.; Iennarella-Servantez, C.A.; Burns, L.E.; Schrunk, D.; et al. Canine Intestinal Organoids as a Novel In Vitro Model of Intestinal Drug Permeability: A Proof-of-Concept Study. Cells 2023, 12, 1269. https://doi.org/10.3390/cells12091269
Sahoo DK, Martinez MN, Dao K, Gabriel V, Zdyrski C, Jergens AE, Atherly T, Iennarella-Servantez CA, Burns LE, Schrunk D, et al. Canine Intestinal Organoids as a Novel In Vitro Model of Intestinal Drug Permeability: A Proof-of-Concept Study. Cells. 2023; 12(9):1269. https://doi.org/10.3390/cells12091269
Chicago/Turabian StyleSahoo, Dipak Kumar, Marilyn N. Martinez, Kimberly Dao, Vojtech Gabriel, Christopher Zdyrski, Albert E. Jergens, Todd Atherly, Chelsea A. Iennarella-Servantez, Laura E. Burns, Dwayne Schrunk, and et al. 2023. "Canine Intestinal Organoids as a Novel In Vitro Model of Intestinal Drug Permeability: A Proof-of-Concept Study" Cells 12, no. 9: 1269. https://doi.org/10.3390/cells12091269