
Comprehensive Characterization of Tissues Derived from Animals at Different Regenerative Stages: A Comparative Analysis between Fetal and Adult Mouse Skin
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice Breeding
2.2. Skin Tissue Dissection and Biopsy Sampling
2.3. Macroscopic Visualization
2.4. Molecular Analysis
2.4.1. Nuclear Visualization and DNA Quantification
2.4.2. Lipid Visualization and Quantification
2.5. Histological Stains and Immunohistochemistry
2.6. Sulfated Glycosaminoglycans (sGAGs) Visualization and Quantification
2.7. Scanning Electron Microscopy (SEM) and Structural Analysis
2.8. Functional Analysis
2.8.1. Metabolic Activity
2.8.2. Water Absorption Capacity
2.8.3. Mechanical Characterization
2.9. Statistical Analysis
3. Results and Discussion
3.1. Macroscopic Analysis
3.2. Cellular and Molecular Features
3.3. ECM Composition and Structure
3.3.1. Fibrous Proteins
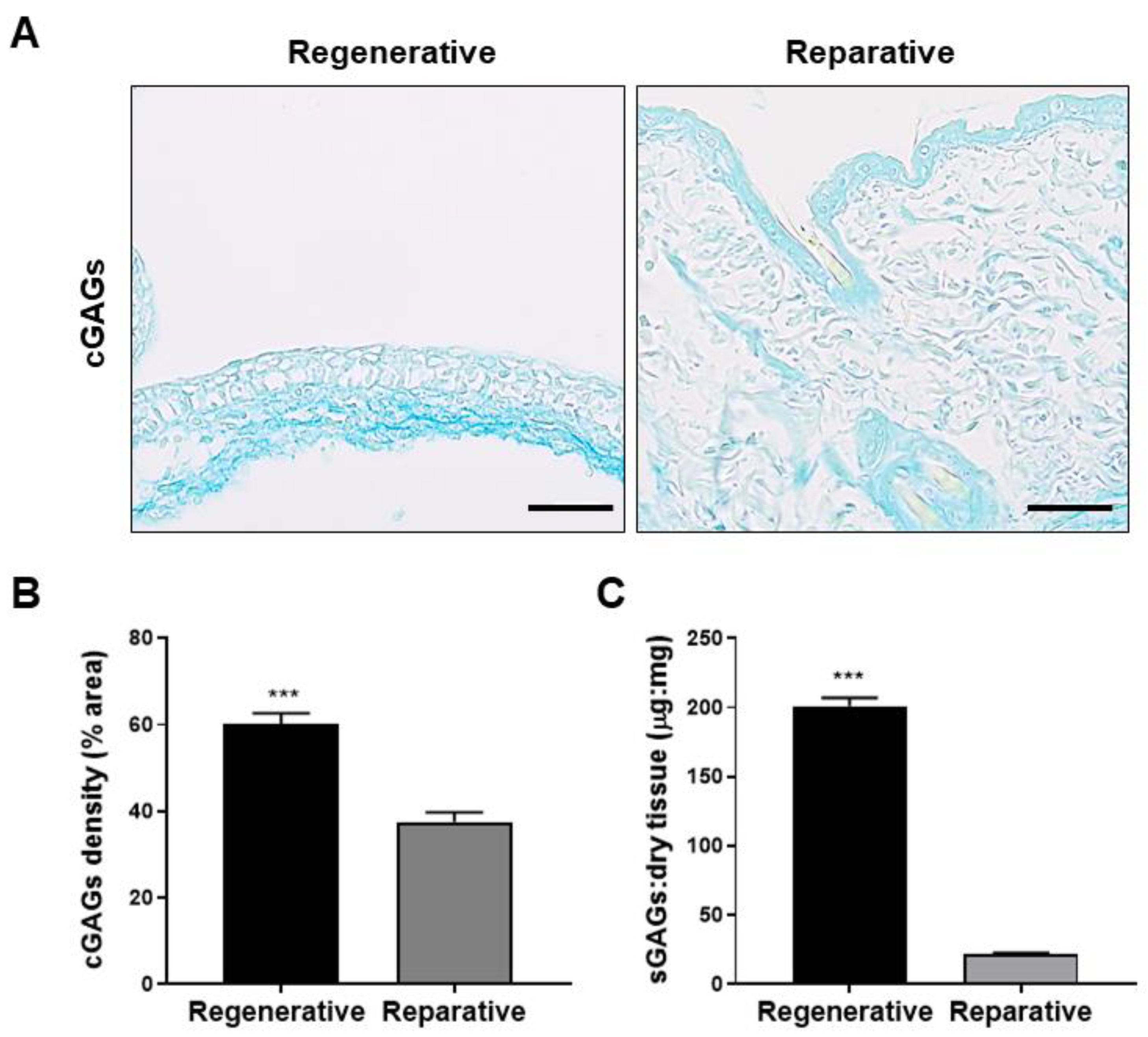
3.3.2. Glycosaminoglycans
3.3.3. Ultrastructural Analysis
3.3.4. Proteoglycans and Glycoproteins
3.4. Functional Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, R.S.; Newmark, P.A. The Cell Biology of Regeneration. J. Cell Biol. 2012, 196, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Hinman, V.; Cary, G.; Wolff, A.; Olga; Pattinato, J. Analysis of Sea Star Larval Regeneration Reveals Conserved Processes of Whole-Body Regeneration across the Metazoa. Dev. Biol. 2017, 118232. [Google Scholar] [CrossRef]
- Daponte, V.; Tylzanowski, P.; Forlino, A. Appendage Regeneration in Vertebrates: What Makes This Possible? Cells 2021, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.R.; Echeverri, K. Learning from Regeneration Research Organisms: The Circuitous Road to Scar Free Wound Healing. Dev. Biol. 2018, 433, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Larson, B.J.; Longaker, M.T.; Lorenz, H.P. Scarless Fetal Wound Healing: A Basic Science Review. Plast. Reconstr. Surg. 2010, 126, 1172–1180. [Google Scholar] [CrossRef]
- Wilgus, T.A. Regenerative Healing in Fetal Skin: A Review of the Literature. Ostomy Wound Manag. 2007, 53, 16–31; quiz 32–33. [Google Scholar]
- Wulff, B.C.; Parent, A.E.; Meleski, M.A.; DiPietro, L.A.; Schrementi, M.E.; Wilgus, T.A. Mast Cells Contribute to Scar Formation during Fetal Wound Healing. J. Investig. Dermatol. 2012, 132, 458–465. [Google Scholar] [CrossRef]
- Walmsley, G.G.; Hu, M.S.; Hong, W.X.; Maan, Z.N.; Lorenz, H.P.; Longaker, M.T. A Mouse Fetal Skin Model of Scarless Wound Repair. J. Vis. Exp. 2015, 52297. [Google Scholar] [CrossRef]
- Ashcroft, G.S.; Horan, M.A.; Ferguson, M.W. The Effects of Ageing on Cutaneous Wound Healing in Mammals. J. Anat. 1995, 187 Pt 1, 1–26. [Google Scholar]
- Coffman, J.A.; Rieger, S.; Rogers, A.N.; Updike, D.L.; Yin, V.P. Comparative Biology of Tissue Repair, Regeneration and Aging. Npj Regen. Med. 2016, 1, 16003. [Google Scholar] [CrossRef]
- Sgonc, R.; Gruber, J. Age-Related Aspects of Cutaneous Wound Healing: A Mini-Review. Gerontology 2013, 59, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.W.; Monaghan, J.R.; Voss, S.R.; Maden, M. Skin Regeneration in Adult Axolotls: A Blueprint for Scar-Free Healing in Vertebrates. PLoS ONE 2012, 7, e32875. [Google Scholar] [CrossRef]
- Richardson, R.; Slanchev, K.; Kraus, C.; Knyphausen, P.; Eming, S.; Hammerschmidt, M. Adult Zebrafish as a Model System for Cutaneous Wound-Healing Research. J. Investig. Dermatol. 2013, 133, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Namazi, M.R.; Fallahzadeh, M.K.; Schwartz, R.A. Strategies for Prevention of Scars: What Can We Learn from Fetal Skin? Int. J. Dermatol. 2011, 50, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.W.; Maden, M. New Insights into Vertebrate Skin Regeneration. Int. Rev. Cell Mol. Biol. 2014, 310, 129–169. [Google Scholar] [CrossRef] [PubMed]
- Walraven, M.; Talhout, W.; Beelen, R.H.J.; van Egmond, M.; Ulrich, M.M.W. Healthy Human Second-Trimester Fetal Skin Is Deficient in Leukocytes and Associated Homing Chemokines. Wound Repair Regen. 2016, 24, 533–541. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.; Kozin, E.D.; Keswani, S.G.; Vaikunth, S.S.; Katz, A.B.; Zoltick, P.W.; Favata, M.; Radu, A.P.; Soslowsky, L.J.; Herlyn, M.; et al. Permissive Environment in Postnatal Wounds Induced by Adenoviral-Mediated Overexpression of the Anti-Inflammatory Cytokine Interleukin-10 Prevents Scar Formation. Wound Repair Regen. 2008, 16, 70–79. [Google Scholar] [CrossRef]
- Jo, H.; Brito, S.; Kwak, B.M.; Park, S.; Lee, M.-G.; Bin, B.-H. Applications of Mesenchymal Stem Cells in Skin Regeneration and Rejuvenation. Int. J. Mol. Sci. 2021, 22, 2410. [Google Scholar] [CrossRef]
- Yao, B.; Huang, S.; Gao, D.; Xie, J.; Liu, N.; Fu, X. Age-Associated Changes in Regenerative Capabilities of Mesenchymal Stem Cell: Impact on Chronic Wounds Repair: MSCs Ageing Impact on Chronic Wounds Repair. Int. Wound J. 2016, 13, 1252–1259. [Google Scholar] [CrossRef]
- Rando, T.A.; Jones, D.L. Regeneration, Rejuvenation, and Replacement: Turning Back the Clock on Tissue Aging. Cold Spring Harb. Perspect. Biol. 2021, 13, a040907. [Google Scholar] [CrossRef]
- Rando, T.A.; Chang, H.Y. Aging, Rejuvenation, and Epigenetic Reprogramming: Resetting the Aging Clock. Cell 2012, 148, 46–57. [Google Scholar] [CrossRef]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719–1733.e12. [Google Scholar] [CrossRef]
- Chen, W.Y.; Grant, M.E.; Schor, A.M.; Schor, S.L. Differences between Adult and Foetal Fibroblasts in the Regulation of Hyaluronate Synthesis: Correlation with Migratory Activity. J. Cell Sci. 1989, 94, 577–584. [Google Scholar] [CrossRef]
- Estes, J.M.; Berg, J.S.V.; Adzick, N.S.; MacGillivray, T.E.; Desmoulière, A.; Gabbiani, G. Phenotypic and Functional Features of Myofibroblasts in Sheep Fetal Wounds. Differentiation 1994, 56, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef]
- Van Exan, R.J.; Hardy, M.H. The Differentiation of the Dermis in the Laboratory Mouse. Am. J. Anat. 1984, 169, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A.; Kretzschmar, K.; Watt, F.M. Reprogramming Adult Dermis to a Neonatal State through Epidermal Activation of β-Catenin. Development 2011, 138, 5189–5199. [Google Scholar] [CrossRef]
- Whitby, D.J.; Ferguson, M.W.J. Immunohistochemical Localization of Growth Factors in Fetal Wound Healing. Dev. Biol. 1991, 147, 207–215. [Google Scholar] [CrossRef]
- Dang, C.M.; Beanes, S.R.; Soo, C.; Ting, K.; Benhaim, P.; Hedrick, M.H.; Lorenz, P.H. Decreased Expression of Fibroblast and Keratinocyte Growth Factor Isoforms and Receptors during Scarless Repair. Plast. Reconstr. Surg. 2003, 111, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Wilgus, T.A.; Ferreira, A.M.; Oberyszyn, T.M.; Bergdall, V.K.; DiPietro, L.A. Regulation of Scar Formation by Vascular Endothelial Growth Factor. Lab. Investig. 2008, 88, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Peled, Z.M.; Rhee, S.J.; Hsu, M.; Chang, J.; Krummel, T.M.; Longaker, M.T. The Ontogeny of Scarless Healing II: EGF and PDGF-B Gene Expression in Fetal Rat Skin and Fibroblasts as a Function of Gestational Age. Ann. Plast. Surg. 2001, 47, 417–424. [Google Scholar] [CrossRef]
- Alaish, S.M.; Yager, D.; Diegelmann, R.F.; Cohen, I.K. Biology of Fetal Wound Healing: Hyaluronate Receptor Expression in Fetal Fibroblasts. J. Pediatr. Surg. 1994, 29, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Longaker, M.T.; Chiu, E.S.; Harrison, M.R.; Crombleholme, T.M.; Langer, J.C.; Duncan, B.W.; Adzick, N.S.; Verrier, E.D.; Stern, R. Studies in Fetal Wound Healing IV. Hyaluronic Acid-Stimulating Activity Distinguishes Fetal Wound Fluid from Adult Wound Fluid. Ann. Surg. 1989, 210, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Mast, B.A.; Flood, L.C.; Haynes, J.H.; Depalma, R.L.; Cohen, I.K.; Diegelmann, R.F.; Krummel, T.M. Hyaluronic Acid Is a Major Component of the Matrix of Fetal Rabbit Skin and Wounds: Implications for Healing by Regeneration. Matrix 1991, 11, 63–68. [Google Scholar] [CrossRef]
- Ouyang, X.; Panetta, N.J.; Talbott, M.D.; Payumo, A.Y.; Halluin, C.; Longaker, M.T.; Chen, J.K. Hyaluronic Acid Synthesis Is Required for Zebrafish Tail Fin Regeneration. PLoS ONE 2017, 12, e0171898. [Google Scholar] [CrossRef]
- de Oliveira, J.D.; Carvalho, L.S.; Gomes, A.M.V.; Queiroz, L.R.; Magalhães, B.S.; Parachin, N.S. Genetic Basis for Hyper Production of Hyaluronic Acid in Natural and Engineered Microorganisms. Microb. Cell Factories 2016, 15, 119. [Google Scholar] [CrossRef]
- Cullen, B.; Silcock, D.; Brown, L.J.; Gosiewska, A.; Geesin, J.C. The Differential Regulation and Secretion of Proteinases from Fetal and Neonatal Fibroblasts by Growth Factors. Int. J. Biochem. Cell Biol. 1997, 29, 241–250. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Dela Justina, V.; dos Passos Junior, R.R.; Bressan, A.F.; Tostes, R.C.; Carneiro, F.S.; Soares, T.S.; Volpato, G.T.; Lima, V.V.; Martin, S.S.; Giachini, F.R. O-Linked N-Acetyl-Glucosamine Deposition in Placental Proteins Varies According to Maternal Glycemic Levels. Life Sci. 2018, 205, 18–25. [Google Scholar] [CrossRef]
- Farshi Azhar, F.; Olad, A.; Salehi, R. Fabrication and Characterization of Chitosan–Gelatin/Nanohydroxyapatite–Polyaniline Composite with Potential Application in Tissue Engineering Scaffolds. Des. Monomers Polym. 2014, 17, 654–667. [Google Scholar] [CrossRef]
- Gilliver, S.C.; Ruckshanthi, J.P.D.; Hardman, M.J.; Nakayama, T.; Ashcroft, G.S. Sex Dimorphism in Wound Healing: The Roles of Sex Steroids and Macrophage Migration Inhibitory Factor. Endocrinology 2008, 149, 5747–5757. [Google Scholar] [CrossRef] [PubMed]
- Troncoso, F.; Herlitz, K.; Acurio, J.; Aguayo, C.; Guevara, K.; Castro, F.O.; Godoy, A.S.; San Martin, S.; Escudero, C. Advantages in Wound Healing Process in Female Mice Require Upregulation A2A-Mediated Angiogenesis under the Stimulation of 17β-Estradiol. Int. J. Mol. Sci. 2020, 21, 7145. [Google Scholar] [CrossRef]
- Coolen, N.A.; Schouten, K.C.W.M.; Middelkoop, E.; Ulrich, M.M.W. Comparison between Human Fetal and Adult Skin. Arch. Dermatol. Res. 2010, 302, 47–55. [Google Scholar] [CrossRef]
- Cuttle, L.; Nataatmadja, M.; Fraser, J.F.; Kempf, M.; Kimble, R.M.; Hayes, M.T. Collagen in the Scarless Fetal Skin Wound: Detection with Picrosirius-Polarization. Wound Repair Regen. 2005, 13, 198–204. [Google Scholar] [CrossRef]
- Jarc, E.; Petan, T. Lipid Droplets and the Management of Cellular Stress. Yale J. Biol. Med. 2019, 92, 435–452. [Google Scholar]
- Knox, S.; O’Boyle, N.M. Skin Lipids in Health and Disease: A Review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef] [PubMed]
- Welte, M.A. Expanding Roles for Lipid Droplets. Curr. Biol. 2015, 25, R470–R481. [Google Scholar] [CrossRef]
- Koh, T.J.; DiPietro, L.A. Inflammation and Wound Healing: The Role of the Macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Martins Cardoso, R.; Absalah, S.; Van Eck, M.; Bouwstra, J.A. Barrier Lipid Composition and Response to Plasma Lipids: A Direct Comparison of Mouse Dorsal Back and Ear Skin. Exp. Dermatol. 2020, 29, 548–555. [Google Scholar] [CrossRef]
- Hardman, M.J.; Sisi, P.; Banbury, D.N.; Byrne, C. Patterned Acquisition of Skin Barrier Function during Development. Development 1998, 125, 1541–1552. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. PERSPECTIVE ARTICLE: Growth Factors and Cytokines in Wound Healing: Growth Factors and Cytokines in Wound Healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Alexander, C.M.; Kasza, I.; Yen, C.-L.E.; Reeder, S.B.; Hernando, D.; Gallo, R.L.; Jahoda, C.A.B.; Horsley, V.; MacDougald, O.A. Dermal White Adipose Tissue: A New Component of the Thermogenic Response. J. Lipid Res. 2015, 56, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Merkel, J.R.; DiPaolo, B.R.; Hallock, G.G.; Rice, D.C. Type I and Type III Collagen Content of Healing Wounds in Fetal and Adult Rats. Exp. Biol. Med. 1988, 187, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Mathew-Steiner, S.S.; Roy, S.; Sen, C.K. Collagen in Wound Healing. Bioengineering 2021, 8, 63. [Google Scholar] [CrossRef]
- Hu, M.S.; Borrelli, M.R.; Hong, W.X.; Malhotra, S.; Cheung, A.T.M.; Ransom, R.C.; Rennert, R.C.; Morrison, S.D.; Lorenz, H.P.; Longaker, M.T. Embryonic Skin Development and Repair. Organogenesis 2018, 14, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Volk, S.W.; Wang, Y.; Mauldin, E.A.; Liechty, K.W.; Adams, S.L. Diminished Type III Collagen Promotes Myofibroblast Differentiation and Increases Scar Deposition in Cutaneous Wound Healing. Cells Tissues Organs 2011, 194, 25–37. [Google Scholar] [CrossRef]
- Park, S. Building vs. Rebuilding Epidermis: Comparison Embryonic Development and Adult Wound Repair. Front. Cell Dev. Biol. 2022, 9, 796080. [Google Scholar] [CrossRef]
- Egging, D.F.; van Vlijmen, I.; Starcher, B.; Gijsen, Y.; Zweers, M.C.; Blankevoort, L.; Bristow, J.; Schalkwijk, J. Dermal Connective Tissue Development in Mice: An Essential Role for Tenascin-X. Cell Tissue Res. 2006, 323, 465–474. [Google Scholar] [CrossRef]
- Senior, R.M.; Griffin, G.L.; Mecham, R.P.; Wrenn, D.S.; Prasad, K.U.; Urry, D.W. Val-Gly-Val-Ala-Pro-Gly, a Repeating Peptide in Elastin, Is Chemotactic for Fibroblasts and Monocytes. J. Cell Biol. 1984, 99, 870–874. [Google Scholar] [CrossRef]
- Longaker, M.T.; Chiu, E.S.; Adzick, N.S.; Stern, M.; Harrison, M.R.; Stern, R. Studies in Fetal Wound Healing V. A Prolonged Presence of Hyaluronic Acid Characterizes Fetal Wound Fluid. Ann. Surg. 1991, 213, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; Maytin, E.V.; Mack, J.A.; Hascall, V.C.; Atanelishvili, I.; Moreno Rodriguez, R.; Markwald, R.R.; Misra, S. Roles of Proteoglycans and Glycosaminoglycans in Wound Healing and Fibrosis. Int. J. Cell Biol. 2015, 2015, 834893. [Google Scholar] [CrossRef]
- Samourides, A.; Browning, L.; Hearnden, V.; Chen, B. The Effect of Porous Structure on the Cell Proliferation, Tissue Ingrowth and Angiogenic Properties of Poly(Glycerol Sebacate Urethane) Scaffolds. Mater. Sci. Eng. C 2020, 108, 110384. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Li, Q.; Wang, X.; Cao, X.; Zgonis, M.H.; Esterhai, J.L.; Shenoy, V.B.; Han, L.; Mauck, R.L. Maturation State and Matrix Microstructure Regulate Interstitial Cell Migration in Dense Connective Tissues. Sci. Rep. 2018, 8, 3295. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.M.; O’Brien, F.J. Understanding the Effect of Mean Pore Size on Cell Activity in Collagen-Glycosaminoglycan Scaffolds. Cell Adhes. Migr. 2010, 4, 377–381. [Google Scholar] [CrossRef]
- Buckley, M.R.; Evans, E.B.; Matuszewski, P.E.; Chen, Y.-L.; Satchel, L.N.; Elliott, D.M.; Soslowsky, L.J.; Dodge, G.R. Distributions of Types I, II and III Collagen by Region in the Human Supraspinatus Tendon. Connect. Tissue Res. 2013, 54, 374–379. [Google Scholar] [CrossRef]
- Hayes, A.J.; Farrugia, B.L.; Biose, I.J.; Bix, G.J.; Melrose, J. Perlecan, A Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan With Roles in Tissue Development Has Relevance to Connective Tissue Repair and Regeneration. Front. Cell Dev. Biol. 2022, 10, 856261. [Google Scholar] [CrossRef]
- Bassat, E.; Mutlak, Y.E.; Genzelinakh, A.; Shadrin, I.Y.; Baruch Umansky, K.; Yifa, O.; Kain, D.; Rajchman, D.; Leach, J.; Riabov Bassat, D.; et al. The Extracellular Matrix Protein Agrin Promotes Heart Regeneration in Mice. Nature 2017, 547, 179–184. [Google Scholar] [CrossRef]
- Chakraborty, S.; Sampath, D.; Yu Lin, M.O.; Bilton, M.; Huang, C.-K.; Nai, M.H.; Njah, K.; Goy, P.-A.; Wang, C.-C.; Guccione, E.; et al. Agrin-Matrix Metalloproteinase-12 Axis Confers a Mechanically Competent Microenvironment in Skin Wound Healing. Nat. Commun. 2021, 12, 6349. [Google Scholar] [CrossRef]
- Seppinen, L.; Sormunen, R.; Soini, Y.; Elamaa, H.; Heljasvaara, R.; Pihlajaniemi, T. Lack of Collagen XVIII Accelerates Cutaneous Wound Healing, While Overexpression of Its Endostatin Domain Leads to Delayed Healing. Matrix Biol. 2008, 27, 535–546. [Google Scholar] [CrossRef]
- Maeba, T.; Yonezawa, T.; Ono, M.; Tomono, Y.; Heljasvaara, R.; Pihlajaniemi, T.; Inagawa, K.; Oohashi, T. Collagen XVIII Deposition in the Basement Membrane Zone beneath the Newly Forming Epidermis during Wound Healing in Mice. Acta Med. Okayama 2019, 73, 135–146. [Google Scholar]
- Zheng, Z.; Zhang, X.; Dang, C.; Beanes, S.; Chang, G.X.; Chen, Y.; Li, C.-S.; Lee, K.S.; Ting, K.; Soo, C. Fibromodulin Is Essential for Fetal-Type Scarless Cutaneous Wound Healing. Am. J. Pathol. 2016, 186, 2824–2832. [Google Scholar] [CrossRef]
- Zheng, Z.; James, A.W.; Li, C.; Jiang, W.; Wang, J.Z.; Chang, G.X.; Lee, K.S.; Chen, F.; Berthiaume, E.A.; Chen, Y.; et al. Fibromodulin Reduces Scar Formation in Adult Cutaneous Wounds by Eliciting a Fetal-like Phenotype. Signal Transduct. Target. Ther. 2017, 2, 17050. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, J.M.; Carrino, D.A.; Baber, M.A.; Caplan, A.I. Versican in Human Fetal Skin Development. Anat. Embryol. 1999, 199, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Soo, C.; Hu, F.-Y.; Zhang, X.; Wang, Y.; Beanes, S.R.; Lorenz, H.P.; Hedrick, M.H.; Mackool, R.J.; Plaas, A.; Kim, S.-J.; et al. Differential Expression of Fibromodulin, a Transforming Growth Factor-β Modulator, in Fetal Skin Development and Scarless Repair. Am. J. Pathol. 2000, 157, 423–433. [Google Scholar] [CrossRef]
- du Cros, D.L.; LeBaron, R.G.; Couchman, J.R. Association of Versican with Dermal Matrices and Its Potential Role in Hair Follicle Development and Cycling. J. Investig. Dermatol. 1995, 105, 426–431. [Google Scholar] [CrossRef]
- Choi, Y.E.; Song, M.J.; Hara, M.; Imanaka-Yoshida, K.; Lee, D.H.; Chung, J.H.; Lee, S.-T. Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging. Int. J. Mol. Sci. 2020, 21, 8693. [Google Scholar] [CrossRef]
- Whitby, D.J.; Longaker, M.T.; Harrison, M.R.; Adzick, N.S.; Ferguson, M.W. Rapid Epithelialisation of Fetal Wounds Is Associated with the Early Deposition of Tenascin. J. Cell Sci. 1991, 99, 583–586. [Google Scholar] [CrossRef]
- Midwood, K.S.; Orend, G. The Role of Tenascin-C in Tissue Injury and Tumorigenesis. J. Cell Commun. Signal. 2009, 3, 287–310. [Google Scholar] [CrossRef] [PubMed]
- Silver, F.H.; Siperko, L.M.; Seehra, G.P. Mechanobiology of Force Transduction in Dermal Tissue: Mechanobiology of Dermal Tissue. Skin Res. Technol. 2003, 9, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ding, Z.; Xu, F.; Lu, Q.; Kaplan, D.L. Subtle Regulation of Scaffold Stiffness for the Optimized Control of Cell Behavior. ACS Appl. Bio Mater. 2019, 2, 3108–3119. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Xu, Q.; Liu, W. An Overview of Substrate Stiffness Guided Cellular Response and Its Applications in Tissue Regeneration. Bioact. Mater. 2022, 15, 82–102. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo, V.; Díaz-Astudillo, P.; Corrales-Orovio, R.; San Martín, S.; Egaña, J.T. Comprehensive Characterization of Tissues Derived from Animals at Different Regenerative Stages: A Comparative Analysis between Fetal and Adult Mouse Skin. Cells 2023, 12, 1215. https://doi.org/10.3390/cells12091215
Castillo V, Díaz-Astudillo P, Corrales-Orovio R, San Martín S, Egaña JT. Comprehensive Characterization of Tissues Derived from Animals at Different Regenerative Stages: A Comparative Analysis between Fetal and Adult Mouse Skin. Cells. 2023; 12(9):1215. https://doi.org/10.3390/cells12091215
Chicago/Turabian StyleCastillo, Valentina, Pamela Díaz-Astudillo, Rocío Corrales-Orovio, Sebastián San Martín, and José Tomás Egaña. 2023. "Comprehensive Characterization of Tissues Derived from Animals at Different Regenerative Stages: A Comparative Analysis between Fetal and Adult Mouse Skin" Cells 12, no. 9: 1215. https://doi.org/10.3390/cells12091215