
Potential Role of IL-37 in Atopic Dermatitis

Abstract
:1. Introduction
2. Discussion
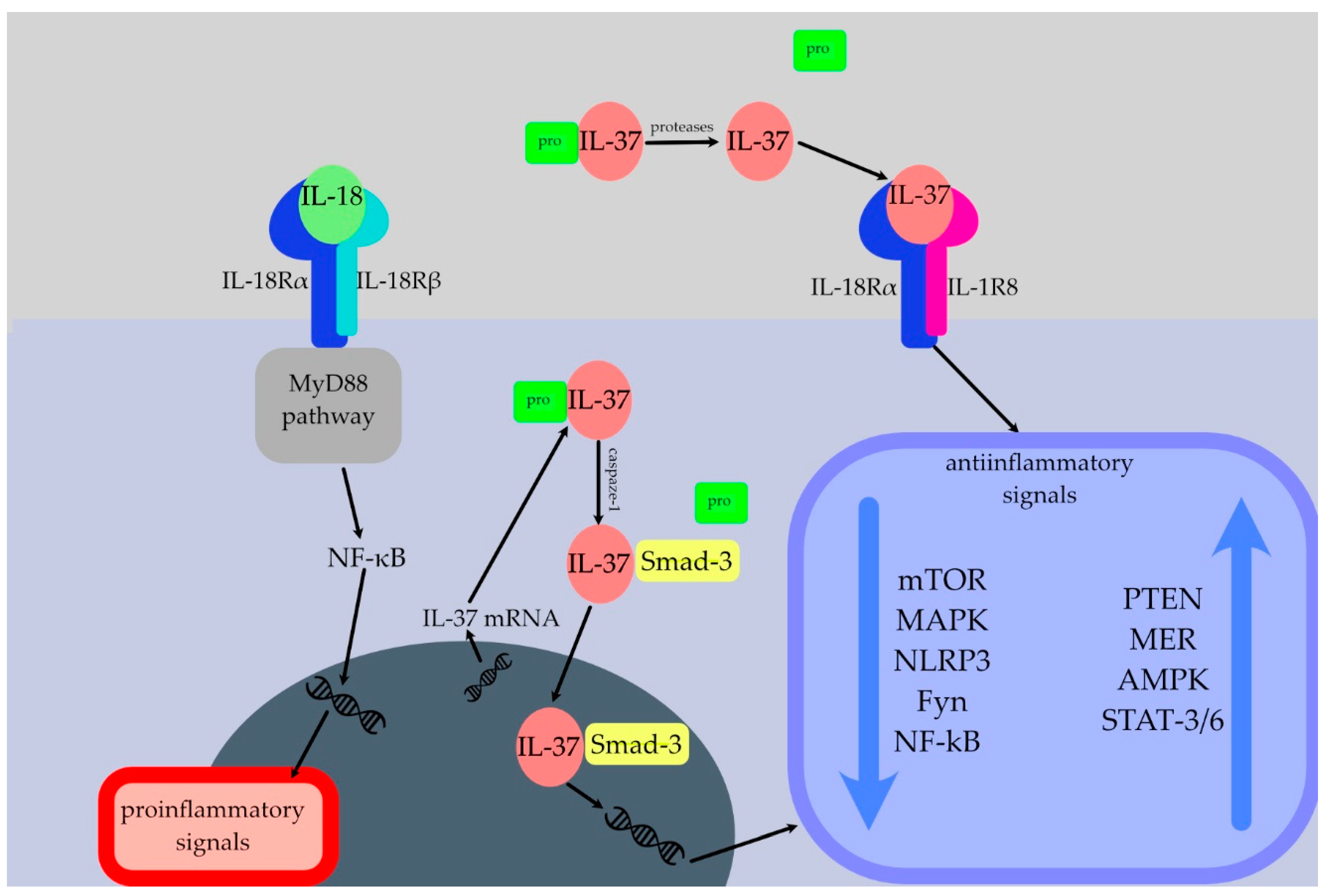
2.1. Brief Overview of the IL-37
2.1.1. Production and Processing
2.1.2. Release
2.1.3. Mechanism of Action
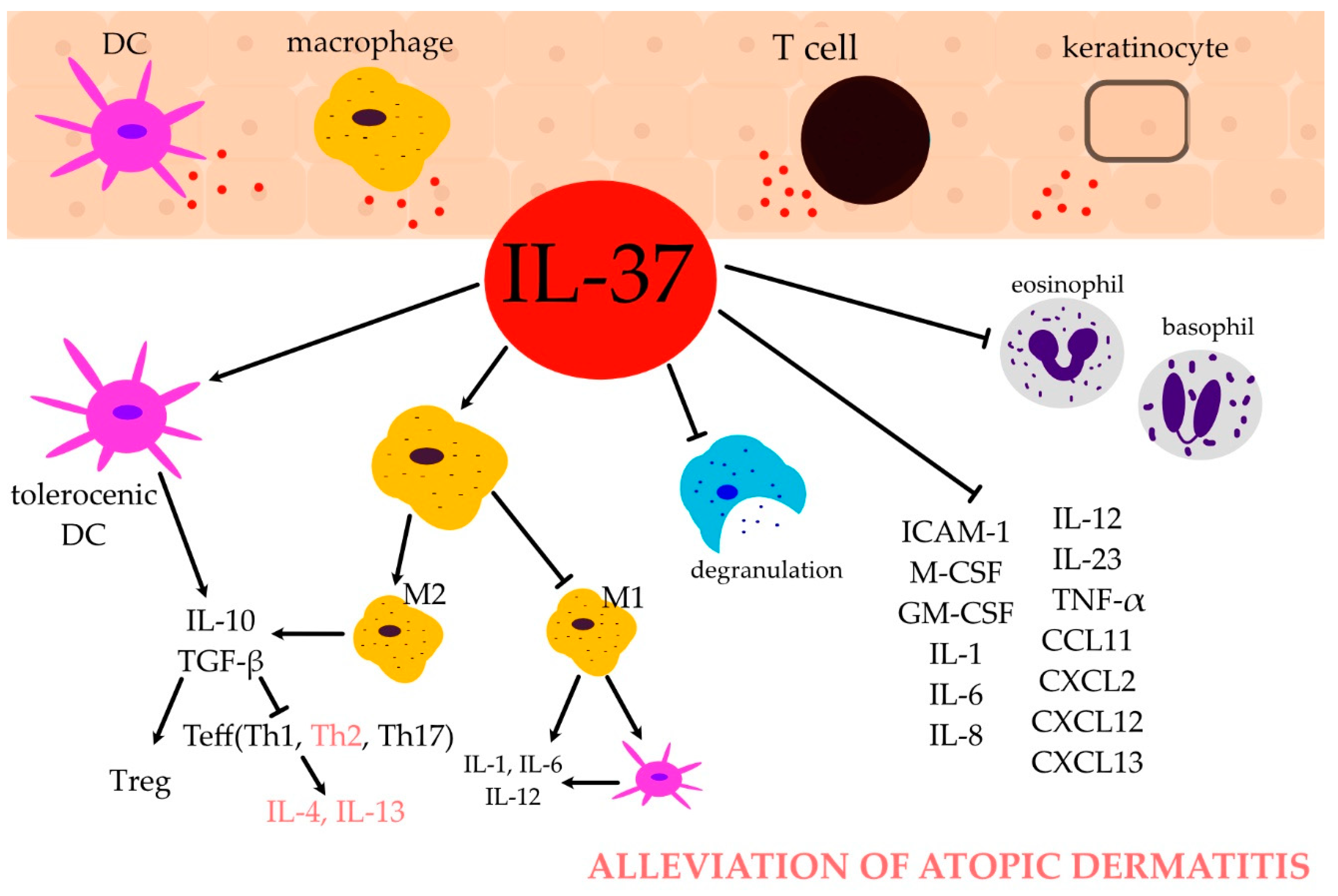
2.2. IL-37 and Immune Cells
2.2.1. Monocytes and Macrophages
2.2.2. Dendritic Cells (DCs)
2.3. Pathogenesis of AD
2.4. IL-37 and Immune Response in AD
2.4.1. T Helper (Th) Cells
2.4.2. Regulatory T Cells (Tregs)
2.4.3. Eosinophils
2.4.4. Basophils
2.4.5. Mast Cells
2.4.6. B Cells
2.5. IL-37 and Skin Barrier Disruption
2.6. IL-37 as a Potential Therapeutic Target in AD
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matarazzo, L.; Hernandez Santana, Y.E.; Walsh, P.T.; Fallon, P.G. The IL-1 cytokine family as custodians of barrier immunity. Cytokine 2022, 154, 155890. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; Dinarello, C.A. Suppression of inflammation and acquired immunity by IL-37. Immunol. Rev. 2018, 281, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Osborne, D.G.; Domenico, J.; Fujita, M. Expression of IL-37 Induces a Regulatory T-Cell-like Phenotype and Function in Jurkat Cells. Cells 2022, 11, 2565. [Google Scholar] [CrossRef]
- Conti, P.; Pregliasco, F.E.; Bellomo, R.G.; Gallenga, C.E.; Caraffa, A.; Kritas, S.K.; Lauritano, D.; Ronconi, G. Mast Cell Cytokines IL-1, IL-33, and IL-36 Mediate Skin Inflammation in Psoriasis: A Novel Therapeutic Approach with the Anti-Inflammatory Cytokines IL-37, IL-38, and IL-1Ra. Int. J. Mol. Sci. 2021, 22, 8076. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Baek, A.R.; Lee, J.H.; Jang, A.S.; Kim, D.J.; Chin, S.S.; Park, S.W. IL-37 Attenuates Lung Fibrosis by Inducing Autophagy and Regulating TGF-β1 Production in Mice. J. Immunol. 2019, 203, 2265–2275. [Google Scholar] [CrossRef]
- Schröder, A.; Lunding, L.P.; Zissler, U.M.; Vock, C.; Webering, S.; Ehlers, J.C.; Orinska, Z.; Chaker, A.; Schmidt-Weber, C.B.; Lang, N.J.; et al. IL-37 regulates allergic inflammation by counterbalancing pro-inflammatory IL-1 and IL-33. Allergy 2022, 77, 856–869. [Google Scholar] [CrossRef]
- Ballak, D.B.; Li, S.; van Diepen, J.A.; Moschen, A.R.; Jansen, H.; Hijmans, A.; Groenhof, G.-J.; Bufler, P.; Kersten, S.; Joosten, L.A.; et al. 12: IL-37 protects against obesity-induced inflammation and insulin resistance. Cytokine 2013, 63, 245–246. [Google Scholar] [CrossRef]
- Coll-Miró, M.; Francos-Quijorna, I.; Santos-Nogueira, E.; Torres-Espin, A.; Bufler, P.; Dinarello, C.A.; López-Vales, R. Beneficial effects of IL-37 after spinal cord injury in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 1411–1416. [Google Scholar] [CrossRef]
- Mantovani, A.; Barajon, I.; Garlanda, C. IL-1 and IL-1 Regulatory Pathways in Cancer Progression and Therapy. Immunol. Rev. 2018, 281, 57. [Google Scholar] [CrossRef]
- Mesjasz, A.; Zawadzka, M.; Chałubiński, M.; Trzeciak, M. Is Atopic Dermatitis Only a Skin Disease? Int. J. Mol. Sci. 2023, 24, 837. [Google Scholar] [CrossRef]
- Hou, T.; Sun, X.; Zhu, J.; Hon, K.-L.; Jiang, P.; Chu, I.M.T.; Tsang, M.; Lam, C.; Zeng, H.; Wong, C. IL-37 Ameliorating Allergic Inflammation in Atopic Dermatitis Through Regulating Microbiota and AMPK-mTOR Signaling Pathway-Modulated Autophagy Mechanism. Front. Immunol. 2020, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Tsang, M.S.-M.; Kan, L.L.-Y.; Li, P.; Chu, I.M.-T.; Lam, C.W.-K.; Wong, C.-K. Il-37 targets tslp-primed basophils to alleviate atopic dermatitis. Int. J. Mol. Sci. 2021, 22, 7393. [Google Scholar] [CrossRef] [PubMed]
- Guttman-Yassky, E.; Diaz, A.; Pavel, A.B.; Fernandes, M.; Lefferdink, R.; Erickson, T.; Canter, T.; Rangel, S.; Peng, X.; Li, R.; et al. Use of Tape Strips to Detect Immune and Barrier Abnormalities in the Skin of Children With Early-Onset Atopic Dermatitis. JAMA Dermatol. 2019, 155, 1358. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Hashimoto-Hachiya, A.; Matsuda-Taniguchi, T.; Takai-Yumine, A.; Takemura, M.; Yan, X.; Furue, M.; Nakahara, T. Natural Compounds Tapinarof and Galactomyces Ferment Filtrate Downregulate IL-33 Expression via the AHR/IL-37 Axis in Human Keratinocytes. Front. Immunol. 2022, 13, 2354. [Google Scholar] [CrossRef]
- Zhou, J.; Gemperline, D.C.; Turner, M.J.; Oldach, J.; Molignano, J.; Sims, J.T.; Stayrook, K.R. Transcriptomic Analysis of Healthy and Atopic Dermatitis Samples Reveals the Role of IL-37 in Human Skin. Immunohorizons 2021, 5, 830–843. [Google Scholar] [CrossRef] [PubMed]
- Lossius, A.H.; Berents, T.L.; Sætre, F.; Nilsen, H.R.; Bradley, M.; Asad, S.; Haraldsen, G.; Sundnes, O.; Holm, J. Early transcriptional changes after UVB treatment in atopic dermatitis include inverse regulation of IL-36γ and IL-37. Exp. Dermatol. 2021, 30, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Tsang, M.S.; Chu, I.M.; Kan, L.L.; Hon, K.; Leung, T.; Lam, C.W.; Wong, C. Skewed inflammation is associated with aberrant interleukin-37 signaling pathway in atopic dermatitis. Allergy 2021, 76, 2102–2114. [Google Scholar] [CrossRef]
- Hu, T.; Todberg, T.; Ewald, D.A.; Hoof, I.; da Rosa, J.C.; Skov, L.; Litman, T. Assessment of Spatial and Temporal Variation in the Skin Transcriptome of Atopic Dermatitis by Use of 1.5 mm Minipunch Biopsies. J. Investig. Dermatol. 2023, 143, 612–620.e6. [Google Scholar] [CrossRef]
- Tsuji, G.; Yamamura, K.; Kawamura, K.; Kido-Nakahara, M.; Ito, T.; Nakahara, T. Regulatory Mechanism of the IL-33–IL-37 Axis via Aryl Hydrocarbon Receptor in Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2023, 24, 14633. [Google Scholar] [CrossRef]
- Borgia, F.; Custurone, P.; Li Pomi, F.; Vaccaro, M.; Alessandrello, C.; Gangemi, S. IL-33 and IL-37: A Possible Axis in Skin and Allergic Diseases. Int. J. Mol. Sci. 2023, 24, 372. [Google Scholar] [CrossRef]
- Pan, Y.; Wen, X.; Hao, D.; Wang, Y.; Wang, L.; He, G.; Jiang, X. The role of IL-37 in skin and connective tissue diseases. Biomed. Pharmacother. 2020, 122, 109705. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.; Luvsannyam, E.; Patel, D.; Nukala, S.; Puvvada, S.R.; Hamid, P. Review of Prominent Cytokines as Superior Therapeutic Targets for Moderate-to-Severe Atopic Dermatitis. Cureus 2020, 12, e9901. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, N.; Nicklin, M.J.; Di Giovine, F.S. Long-range DNA interactions at the IL-1/IL-36/IL-37 gene cluster (2q13) are induced by activation of monocytes. Cytokine 2014, 68, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Tao, X. Current Understanding of IL-37 in Human Health and Disease. Front. Immunol. 2021, 12, 696605. [Google Scholar] [CrossRef]
- Jia, H.; Liu, J.; Han, B. Reviews of Interleukin-37: Functions, Receptors, and Roles in Diseases. BioMed Res. Int. 2018, 2018, 3058640. [Google Scholar] [CrossRef]
- Christodoulou, P.; Kyriakou, T.-C.; Boutsikos, P.; Andreou, M.; Ji, Y.; Xu, D.; Papageorgis, P.; Christodoulou, M.-I. Aberrant Expression and Prognostic Potential of IL-37 in Human Lung Adenocarcinoma. Biomedicines 2022, 10, 3037. [Google Scholar] [CrossRef]
- Zeng, H.; Zhou, K.; Ye, Z. Biology of interleukin-37 and its role in autoimmune diseases (Review). Exp. Ther. Med. 2022, 24, 495. [Google Scholar] [CrossRef]
- Shilovskiy, I.P.; Dyneva, M.E.; Kurbacheva, O.M.; Kudlay, D.A.; Khaitov, M.R. The Role of Interleukin-37 in the Pathogenesis of Allergic Diseases. Acta Naturae 2019, 11, 54. [Google Scholar] [CrossRef]
- Tsutsumi, N.; Kimura, T.; Arita, K.; Ariyoshi, M.; Ohnishi, H.; Yamamoto, T.; Zuo, X.; Maenaka, K.; Park, E.Y.; Kondo, N.; et al. The structural basis for receptor recognition of human interleukin-18. Nat. Commun. 2014, 5, 5340. [Google Scholar] [CrossRef]
- Mao, X.; Zhu, R.; Zhang, F.; Zhong, Y.; Yu, K.; Wei, Y.; Sun, H.; Xu, W.; Luo, Q.; Wang, Y.; et al. IL-37 Plays a Beneficial Role in Patients with Acute Coronary Syndrome. Mediat. Inflamm 2019, 2019, 9515346. [Google Scholar] [CrossRef]
- Li, S.; Neff, C.P.; Barber, K.; Hong, J.; Luo, Y.; Azam, T.; Palmer, B.E.; Fujita, M.; Garlanda, C.; Mantovani, A.; et al. Extracellular forms of IL-37 inhibit innate inflammation in vitro and in vivo but require the IL-1 family decoy receptor IL-1R8. Proc. Natl. Acad. Sci. USA 2015, 112, 2497–2502. [Google Scholar] [CrossRef]
- Kaplanski, G. Interleukin-18: Biological properties and role in disease pathogenesis. Immunol. Rev. 2018, 281, 138. [Google Scholar] [CrossRef]
- Ihim, S.A.; Abubakar, S.D.; Zian, Z.; Sasaki, T.; Saffarioun, M.; Maleknia, S.; Azizi, G. Interleukin-18 cytokine in immunity, inflammation, and autoimmunity: Biological role in induction, regulation, and treatment. Front. Immunol. 2022, 13, 919973. [Google Scholar] [CrossRef]
- McCurdy, S.; Liu, C.A.; Yap, J.; Boisvert, W.A. Potential Role of IL-37 in Atherosclerosis. Cytokine 2019, 122, 154169. [Google Scholar] [CrossRef]
- Nold, M.F.; Nold-Petry, C.A.; Zepp, J.A.; Palmer, B.E.; Bufler, P.; Dinarello, C.A. Interleukin 37 is a fundamental inhibitor of innate immunity. Nat. Immunol. 2010, 11, 1014. [Google Scholar] [CrossRef] [PubMed]
- Rudloff, I.; Cho, S.X.; Lao, J.C.; Ngo, D.; McKenzie, M.; Nold-Petry, C.A.; Nold, M.F. Monocytes and dendritic cells are the primary sources of interleukin 37 in human immune cells. J. Leukoc. Biol. 2017, 101, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Xie, B.; Wu, G.; Hu, J.; Wang, D.; Cai, X.; Li, J. Interleukin-37: The Effect of Anti-Inflammatory Response in Human Coronary Artery Endothelial Cells. Mediat. Inflamm. 2019, 2019, 2650590. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, S.; Baumer, Y.; Toulmin, E.; Lee, B.H.; Boisvert, W.A. Macrophage-Specific Expression of IL-37 in Hyperlipidemic Mice Attenuates Atherosclerosis. J. Immunol. 2017, 199, 3604–3613. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Li, Q.; Su, S.; Dong, W.; Zong, S.; Ma, Q.; Yang, X.; Zuo, D.; Zheng, S.; Meng, X.; et al. Interleukin 37 Suppresses M1 Macrophage Polarization Through Inhibition of the Notch1 and Nuclear Factor Kappa B Pathways. Front. Cell Dev. Biol. 2020, 8, 56. [Google Scholar] [CrossRef]
- Luo, Y.; Cai, X.; Liu, S.; Wang, S.; Nold-Petry, C.A.; Nold, M.F.; Bufler, P.; Norris, D.; Dinarello, C.A.; Fujita, M. Suppression of antigen-specific adaptive immunity by IL-37 via induction of tolerogenic dendritic cells. Proc. Natl. Acad. Sci. USA 2014, 111, 15178–15183. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, J.; Lin, Y.; Que, B.; Chang, C.; Zhang, J.; Liang, Z.; Gao, X.; Liu, S.; Liu, L.; et al. IL-37 inhibits the maturation of dendritic cells through the IL-1R8-TLR4-NF-κB pathway. Biochim. Et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2019, 1864, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular Mechanisms of Atopic Dermatitis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Reszka, E.; Gutowska-Owsiak, D.; Trzeciak, M.; Lange, M.; Jarczak, J.; Niedoszytko, M.; Jablonska, E.; Romantowski, J.; Strapagiel, D.; et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 6484. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Lange, M.; Sokołowska-Wojdyło, M.; Renke, J.; Trzonkowski, P.; Sobjanek, M.; Szczerkowska-Dobosz, A.; Niedoszytko, M.; Górska, A.; Romantowski, J.; et al. The role of regulatory T cells and genes involved in their differentiation in pathogenesis of selected inflammatory and neoplastic skin diseases. Part II: The Treg role in skin diseases pathogenesis. Adv. Dermatol. Allergol. Postȩpy Dermatol. I Alergol. 2017, 34, 405. [Google Scholar] [CrossRef] [PubMed]
- Ewald, D.A.; Malajian, D.; Krueger, J.G.; Workman, C.T.; Wang, T.; Tian, S.; Litman, T.; Guttman-Yassky, E.; Suárez-Fariñas, M. Meta-analysis derived atopic dermatitis (MADAD) transcriptome defines a robust AD signature highlighting the involvement of atherosclerosis and lipid metabolism pathways. BMC Med. Genom. 2015, 8, 60. [Google Scholar] [CrossRef]
- Fujita, H.; Inoue, Y.; Seto, K.; Komitsu, N.; Aihara, M. Interleukin-37 is elevated in subjects with atopic dermatitis. J. Dermatol. Sci. 2013, 69, 173–175. [Google Scholar] [CrossRef]
- Thijs, J.L.; Strickland, I.; Bruijnzeel-Koomen, C.A.; Nierkens, S.; Giovannone, B.; Csomor, E.; Sellman, B.R.; Mustelin, T.; Sleeman, M.A.; de Bruin-Weller, M.S.; et al. Moving toward endotypes in atopic dermatitis: Identification of patient clusters based on serum biomarker analysis. J. Allergy Clin. Immunol. 2017, 140, 730–737. [Google Scholar] [CrossRef]
- He, H.; Bissonnette, R.; Wu, J.; Diaz, A.; Proulx, E.S.-C.; Maari, C.; Jack, C.; Louis, M.; Estrada, Y.; Krueger, J.G.; et al. Tape strips detect distinct immune and barrier profiles in atopic dermatitis and psoriasis. J. Allergy Clin. Immunol. 2021, 147, 199–212. [Google Scholar] [CrossRef]
- Ahmed, M.B.; Ad’hiah, A.H. Reduced levels of interleukin-37 in serum of patients with allergic rhinitis or asthma. Rev. Fr. Allergol. 2021, 61, 410–414. [Google Scholar] [CrossRef]
- Lunding, L.; Webering, S.; Vock, C.; Schröder, A.; Raedler, D.; Schaub, B.; Fehrenbach, H.; Wegmann, M. IL-37 requires IL-18Rα and SIGIRR/IL-1R8 to diminish allergic airway inflammation in mice. Allergy 2015, 70, 366–373. [Google Scholar] [CrossRef]
- Lei, H.; Sun, Y.; Quan, S. IL-37 relieves allergic inflammation by inhibiting the CCL11 signaling pathway in a mouse model of allergic rhinitis. Exp. Ther. Med. 2020, 20, 3114. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Bozza, S.; Oikonomou, V.; Renga, G.; Casagrande, A.; Iannitti, R.G.; Puccetti, M.; Garlanda, C.; Kim, S.; Li, S.; et al. IL-37 Inhibits Inflammasome Activation and Disease Severity in Murine Aspergillosis. PLoS Pathog. 2014, 10, 1004462. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, Y.; Li, C.; Liu, C.; Wang, Z.-H.; Li, Y.-S.; Ke, X.; Hu, G.-H. IL-37 attenuates allergic process via STAT6/STAT3 pathways in murine allergic rhinitis. Int. Immunopharmacol. 2019, 69, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Xiong, Y.; Li, W.; Cui, X.; Cheng, X.; Leng, Q.; He, R. IL-37 inhibits IL-4/IL-13-induced CCL11 production and lung eosinophilia in murine allergic asthma. Allergy 2018, 73, 1642–1652. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Liao, B.; Yu, Q.-H.; Wang, H.; Liu, Y.-B.; Guo, C.-L.; Wang, Z.-C.; Li, Z.-Y.; Wang, Z.-Z.; Ruan, J.-W.; et al. The IL-37–Mex3B–Toll-like receptor 3 axis in epithelial cells in patients with eosinophilic chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2020, 145, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Jiang, B.; Deng, J.; Du, J.; Xiong, W.; Guan, Y.; Wen, Z.; Huang, K.; Huang, Z. IL-37 Alleviates Rheumatoid Arthritis by Suppressing IL-17 and IL-17–Triggering Cytokine Production and Limiting Th17 Cell Proliferation. J. Immunol. 2015, 194, 5110–5119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Hao, F.; Qian, T.; Cheng, H.X. Expression of Helper and Regulatory T Cells in Atopic Dermatitis: A Meta-Analysis. Front. Pediatr. 2022, 10, 777992. [Google Scholar] [CrossRef] [PubMed]
- Shuai, X.; Wei-Min, L.; Tong, Y.L.; Dong, N.; Sheng, Z.Y.; Yao, Y.M. Expression of IL-37 contributes to the immunosuppressive property of human CD4+CD25+ regulatory T cells. Sci. Rep. 2015, 5, 14478. [Google Scholar] [CrossRef]
- Wang, D.W.; Dong, N.; Wu, Y.; Zhu, X.M.; Wang, C.T.; Yao, Y.M. Interleukin-37 Enhances the Suppressive Activity of Naturally Occurring CD4+CD25+ Regulatory T Cells. Sci. Rep. 2016, 6, 38955. [Google Scholar] [CrossRef]
- Ghelani, A.; Bates, D.; Conner, K.; Wu, M.-Z.; Lu, J.; Hu, Y.-L.; Li, C.-M.; Chaudhry, A.; Sohn, S.J. Defining the Threshold IL-2 Signal Required for Induction of Selective Treg Cell Responses Using Engineered IL-2 Muteins. Front. Immunol. 2020, 11, 1106. [Google Scholar] [CrossRef]
- Liu, F.T.; Goodarzi, H.; Chen, H.Y. IgE, mast cells, and eosinophils in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef]
- Zeng-Yun-Ou, Z.; Zhong-Yu, J.; Wei, L. Bidirectional associations between eosinophils, basophils, and lymphocytes with atopic dermatitis: A multivariable Mendelian randomization study. Front. Immunol. 2022, 13, 1001911. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Z.X.; Lian, D.; Haig, A.; Bhattacharjee, R.N.; Jevnikar, A.M. IL-37 inhibits IL-18-induced tubular epithelial cell expression of pro-inflammatory cytokines and renal ischemia-reperfusion injury. Kidney Int. 2015, 87, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Ziegler, S.F. TSLP: From allergy to cancer. Nat. Immunol. 2019, 20, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, Y.; Mogi, K.; Takahashi, K.; Miyake, K.; Yoshikawa, S.; Karasuyama, H. Skin-infiltrating basophils promote atopic dermatitis-like inflammation via IL-4 production in mice. Allergy 2020, 75, 2613–2622. [Google Scholar] [CrossRef]
- Mali, S.S.; Bautista, D.M. Basophils add fuel to the flame of eczema itch. Cell 2021, 184, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Zhan, M.; Wang, L.; Chen, D.; Zhao, N.; Wang, J.; Huang, Y.; Zhang, X.; He, S.; Zhang, H. Upregulation of the expression of Toll-like receptor 9 in basophils in patients with allergic rhinitis: An enhanced expression by allergens. Scand. J. Immunol. 2021, 93, e13003. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Caraffa, A.; Ronconi, G.; Frydas, I.; Theoharides, T.C. Recent progress on pathophysiology, inflammation and defense mechanism of mast cells against invading microbes: Inhibitory effect of IL-37. Cent. Eur. J. Immunol. 2019, 44, 447. [Google Scholar] [CrossRef]
- Li, W.; Ding, F.; Zhai, Y.; Tao, W.; Bi, J.; Fan, H.; Yin, N.; Wang, Z. IL-37 is protective in allergic contact dermatitis through mast cell inhibition. Int. Immunopharmacol. 2020, 83, 106476. [Google Scholar] [CrossRef]
- Saluja, R.; Khan, M.; Church, M.K.; Maurer, M. The role of IL-33 and mast cells in allergy and inflammation. Clin. Transl. Allergy 2015, 5, 33. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Tsilioni, I.; Conti, P. Mast Cells May Regulate The Anti-Inflammatory Activity of IL-37. Int. J. Mol. Sci. 2019, 20, 3701. [Google Scholar] [CrossRef] [PubMed]
- Kader, H.A.; Azeem, M.; Jwayed, S.A.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.A.; Hetta, H.F.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Brahmanti, H.; Rofiq, A.; Cholis, M.; Trijayanti, U.; Purwanti, S. Association of LL-37 and IL-31 serum levels with SCORing Atopic Dermatitis (SCORAD) score in atopic dermatitis patients. Iran. J. Dermatol. 2018, 21, 7–13. [Google Scholar] [CrossRef]
- Furue, M. Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic Implications in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Skin Barrier Abnormalities and Immune Dysfunction in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef] [PubMed]
- Paller, A.S.; Kong, H.H.; Seed, P.; Naik, S.; Scharschmidt, T.C.; Gallo, R.L.; Luger, T.; Irvine, A.D. The microbiome in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 26. [Google Scholar] [CrossRef]
- Lachner, J.; Mlitz, V.; Tschachler, E.; Eckhart, L. Epidermal cornification is preceded by the expression of a keratinocyte-specific set of pyroptosis-related genes. Sci. Rep. 2017, 7, 17446. [Google Scholar] [CrossRef]
- Yan, X.; Tsuji, G.; Hashimoto-Hachiya, A.; Furue, M. Galactomyces Ferment Filtrate Potentiates an Anti-Inflammaging System in Keratinocytes. J. Clin. Med. 2022, 11, 6338. [Google Scholar] [CrossRef]
- Nold-Petry, C.A.; Nold, M.F. Rationale for IL-37 as a novel therapeutic agent in inflammation. Expert Rev. Clin. Immunol. 2022, 18, 1203–1206. [Google Scholar] [CrossRef]
- Abulkhir, A.; Samarani, S.; Amre, D.; Duval, M.; Haddad, E.; Sinnett, D.; Leclerc, J.-M.; Diorio, C.; Ahmad, A. A protective role of IL-37 in cancer: A new hope for cancer patients. J. Leukoc. Biol. 2017, 101, 395–406. [Google Scholar] [CrossRef]
- Xu, J.; Chen, J.; Li, W.; Lian, W.; Huang, J.; Lai, B.; Li, L.; Huang, Z. Additive Therapeutic Effects of Mesenchymal Stem Cells and IL-37 for Systemic Lupus Erythematosus. J. Am. Soc. Nephrol. 2020, 31, 54–65. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, S.; Yap, J.; Irei, J.; Lozano, J.; Boisvert, W.A. IL-37—A putative therapeutic agent in cardiovascular diseases. QJM Int. J. Med. 2022, 115, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, L.; Lu, Z.; Chen, H.; Fan, L.; Xue, Q.; Shi, J.; Li, M.; Li, H.; Gong, J.; et al. IL-37 Represses the Autoimmunity in Myasthenia Gravis via Directly Targeting Follicular Th and B Cells. J. Immunol. 2020, 204, 1736–1745. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Country and Authors | Study Subject or Model | Most Important Findings |
|---|---|---|
| China 2020; Hou et al. [11] | CRISPR/Cas9 human IL-37b knock-in mice |
|
| China 2021; Hou et al. [12] | CRISPR/Cas9 human IL-37b knock-in mice or mice with direct treatment with human IL-37b antibody |
|
| United States of America 2019; Guttman-Yassky et al. [13] | 51 children (less than 5 years); 21 with moderate-to-severe AD with less than 6 months of disease duration; 30 did not have AD; RNA extracted from tape strips; quantitative RT-PCR |
|
| Japan 2022; Tsuji et al. [14] | normal human epidermal keratinocytes |
|
| United States of America 2021; Zhou et al. [15] | skin and blood samples from moderate-to-severe AD treated with topical corticosteroid and those with no topical medications applied to skin for a period of at least 1 week |
|
| Norway 2020; Lossius et al. [16] | 16 adult patients (5 males, 11 females; mean age 32, range 20–73); patients received standard full-body nb-UVB therapy 3×/week for 6–8 weeks with a starting dose of 0.1–0.2 J/cm2 and progressive dose increase; analysis of different questionnaire of AD severity |
|
| China 2021; Hou et al. [17] | blood samples from 20 healthy control subjects (aged 14–57 years) without history of skin, allergic, or inflammatory disease, and 34 subjects (aged 11–49 years) with moderate-to-severe AD; human HaCaT keratinocytes and 3D keratinocytes |
|
| Denmark 2022; Hu et al. [18] | 30 patients with AD and 30 healthy controls; 393 skin samples from multiple anatomical regions and time points; 1.5 mm minipunch biopsies for obtaining multiple full-thickness skin samples from the same subject over time |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesjasz, A.; Trzeciak, M.; Gleń, J.; Jaskulak, M. Potential Role of IL-37 in Atopic Dermatitis. Cells 2023, 12, 2766. https://doi.org/10.3390/cells12232766
Mesjasz A, Trzeciak M, Gleń J, Jaskulak M. Potential Role of IL-37 in Atopic Dermatitis. Cells. 2023; 12(23):2766. https://doi.org/10.3390/cells12232766
Chicago/Turabian StyleMesjasz, Alicja, Magdalena Trzeciak, Jolanta Gleń, and Marta Jaskulak. 2023. "Potential Role of IL-37 in Atopic Dermatitis" Cells 12, no. 23: 2766. https://doi.org/10.3390/cells12232766