
A Brief Review of Bone Cell Function and Importance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of Bone Physiology and Structure
2.1. Main Functions of Bone Tissue
- Structural support;
- Protection of internal organs and soft tissues from damage;
- Locomotion;
- Mineral storage;
- Production of blood cells;
- Endocrine regulation.
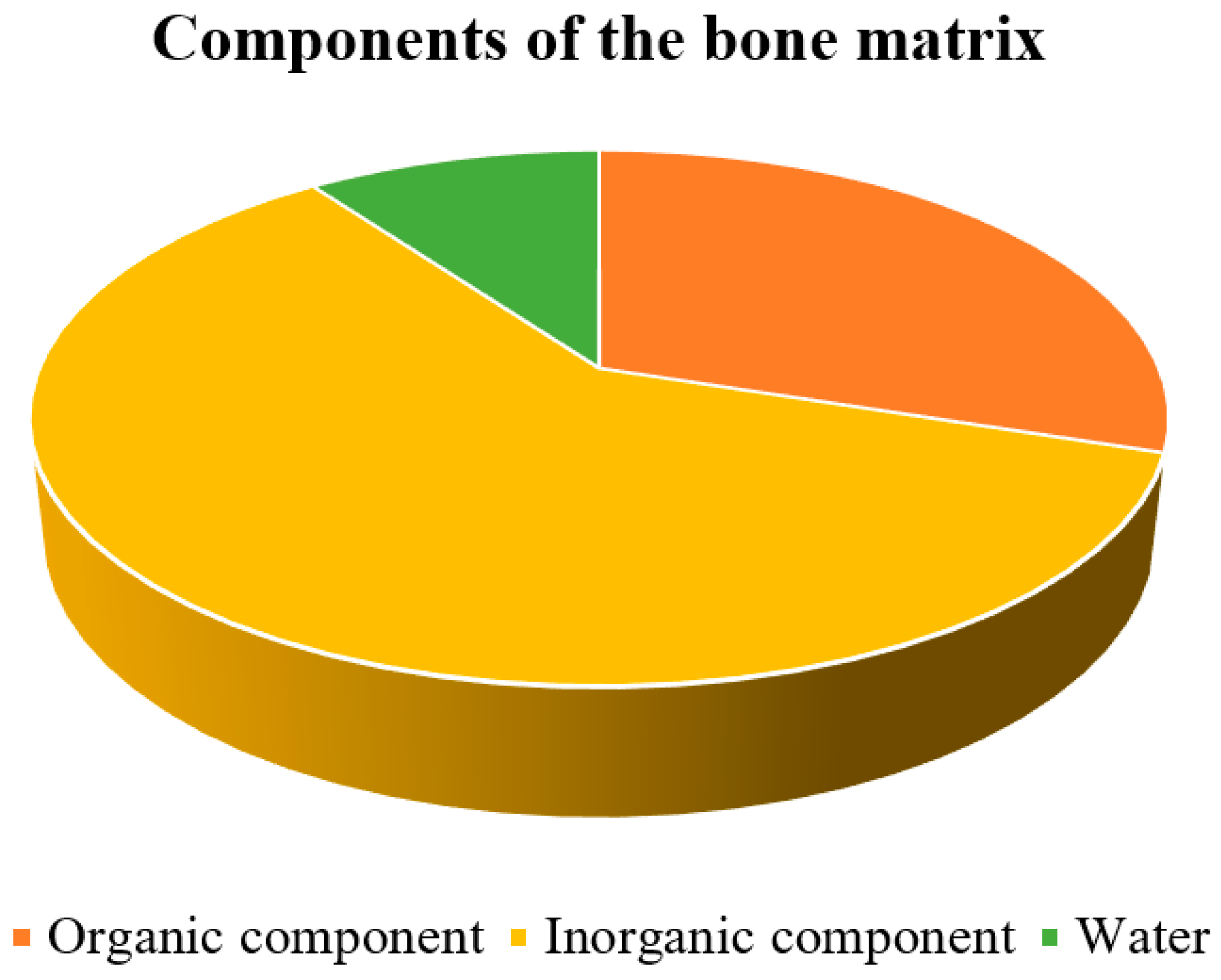
2.2. Microstructure of Bone Tissue
2.2.1. Organic Component of the Bone Matrix
- Osteocalcin—regulates bone formation and the activity of osteoclasts and their precursors; osteocalcin also plays a pivotal role in the hormonal regulation of insulin metabolism and as a mediator of testosterone secretion.
- Osteonectin—can modulate the activity of growth factors, influences cell adhesion, and plays a role in the mineralization and deposition of hydroxyapatite.
- Osteopontin—regulates bone formation, migration, adhesion, and mineralization processes.
- Bone sialoprotein—binds Ca2+ and is involved in mineralization.
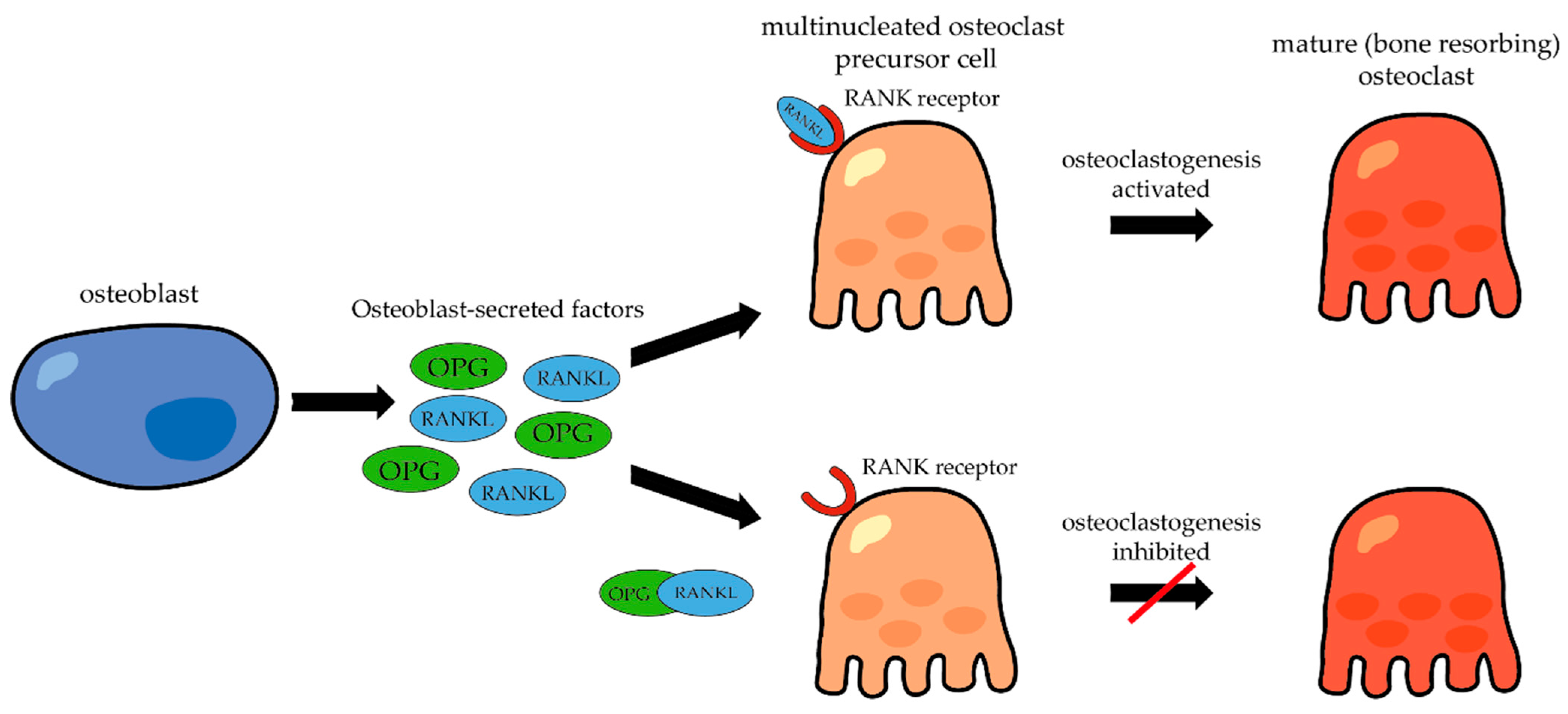
- RANKL (Receptor Activator of Nuclear Factor κ-B Ligand)—functions in the regulation of bone resorption and differentiation of osteoclasts from their precursors.
2.2.2. Inorganic Component of the Bone Matrix
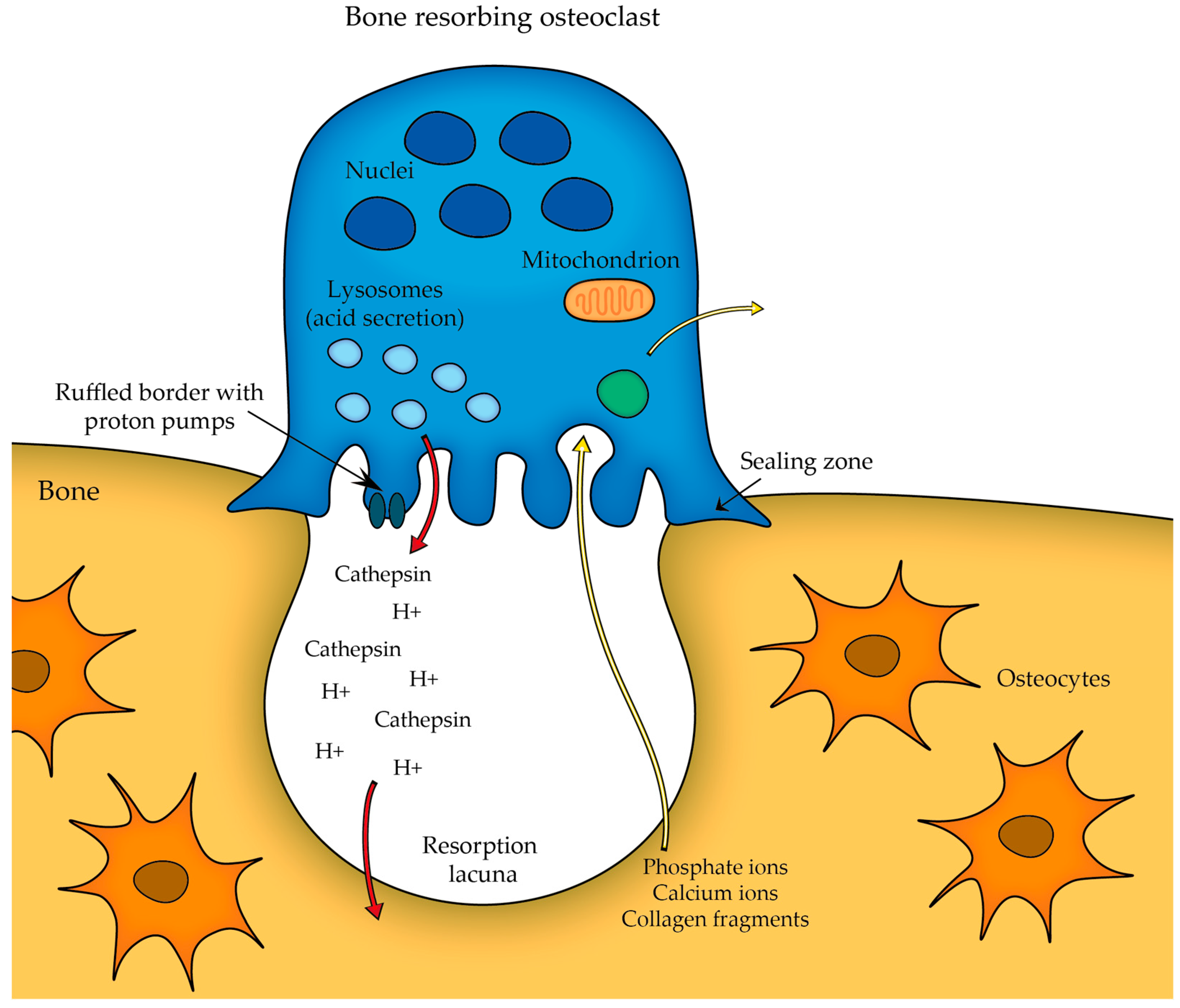
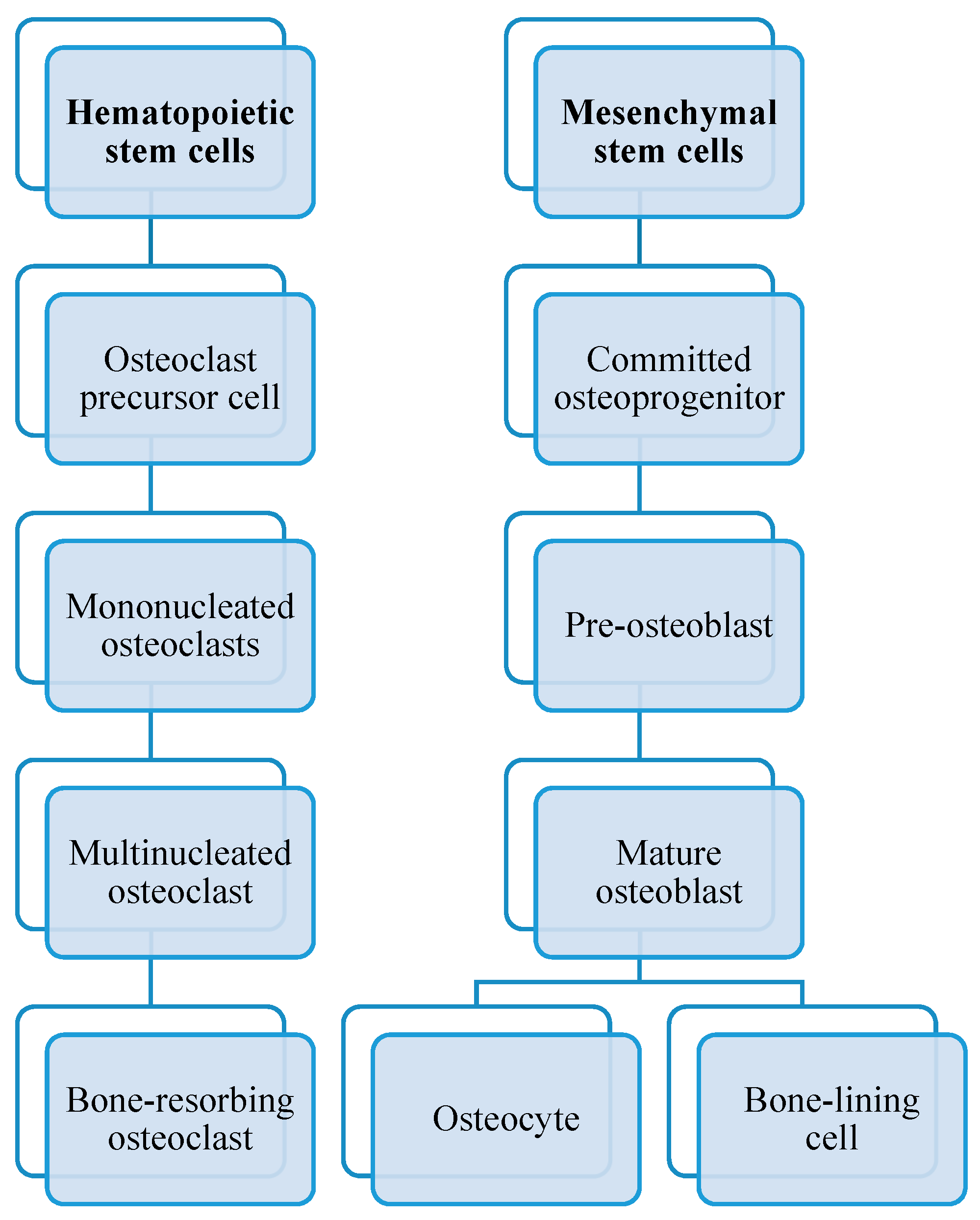
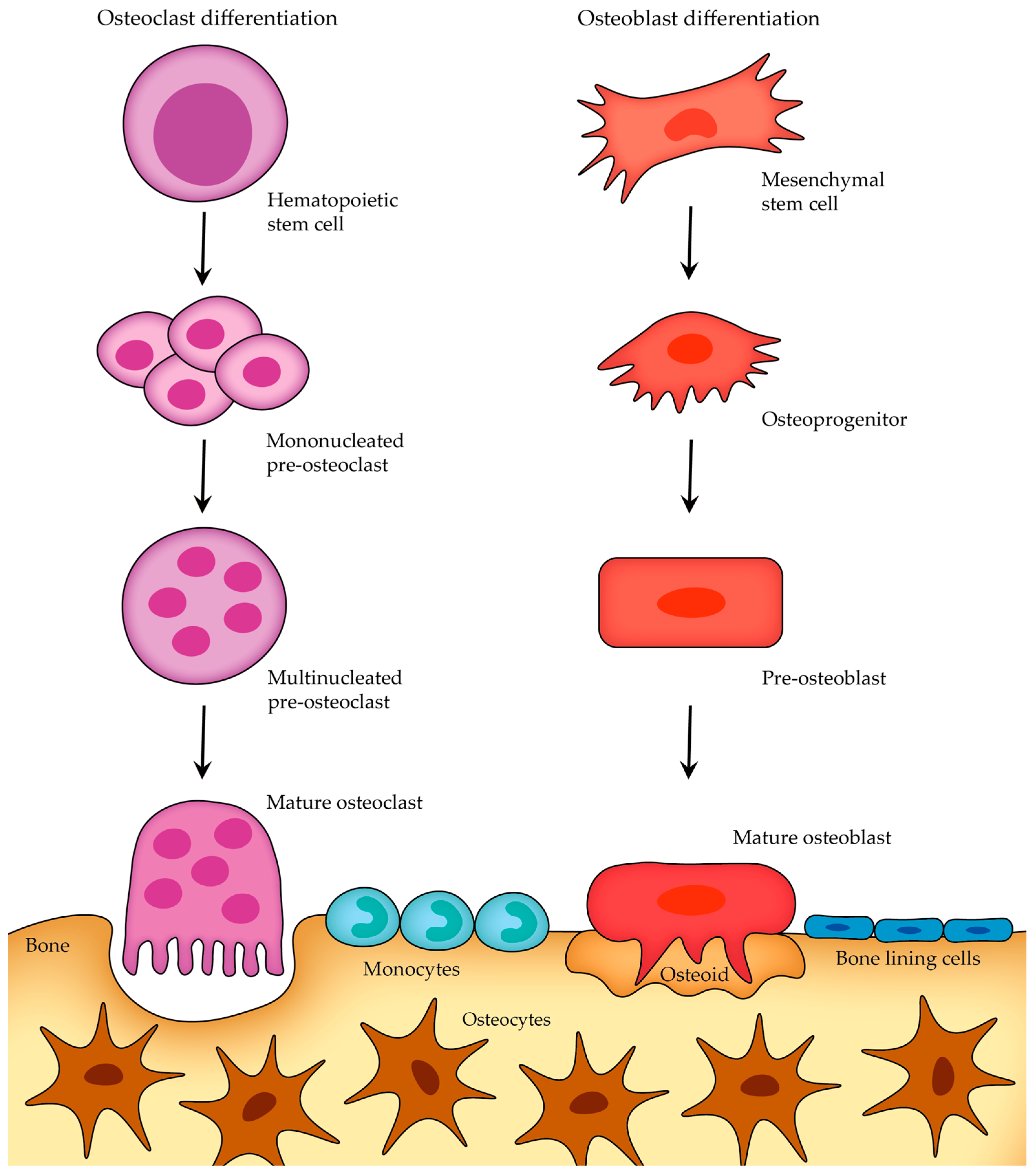
2.2.3. Cells of Bone
- Osteoclasts;
- Osteoblasts;
- Osteocytes;
- Lining cells.
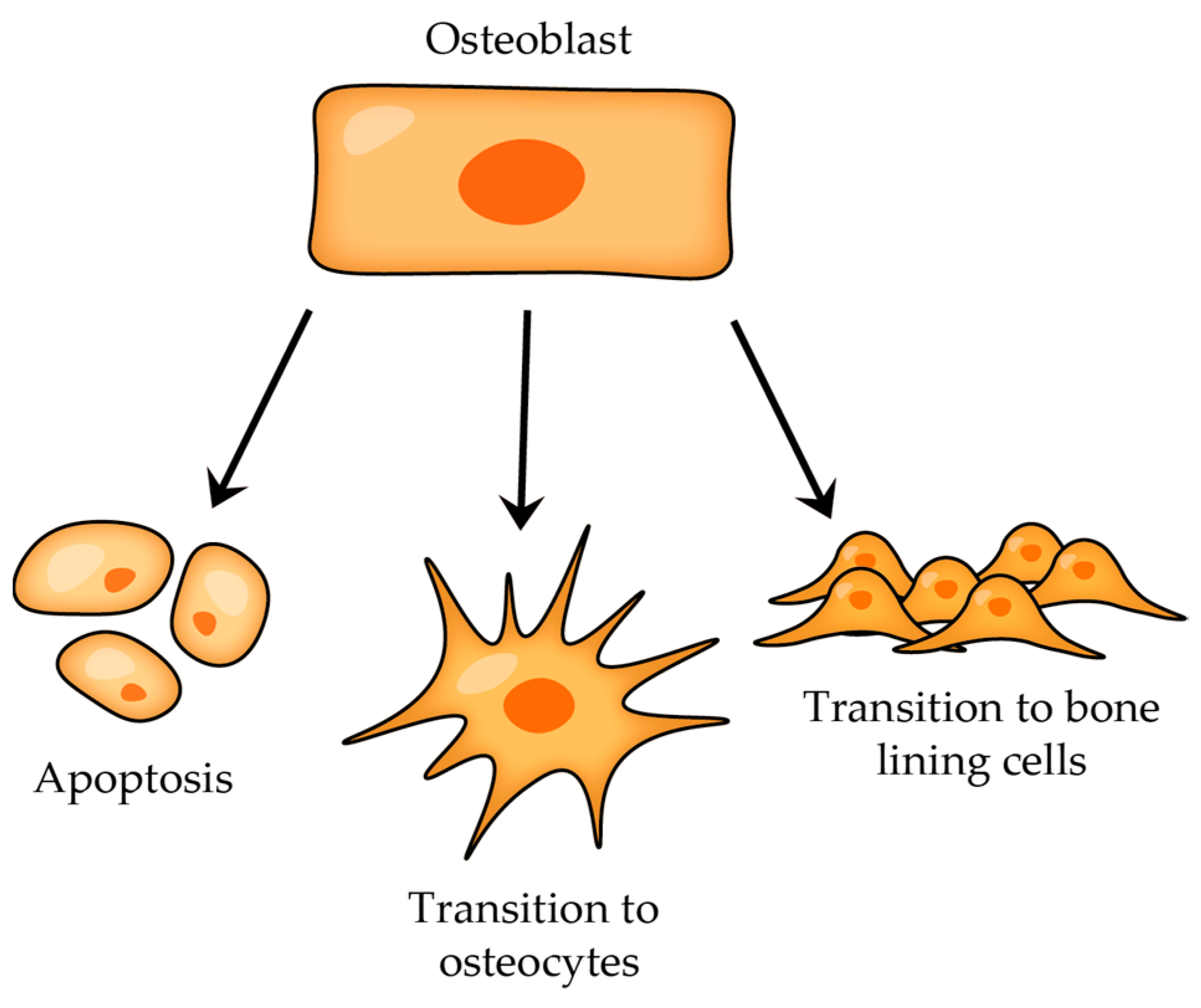
- Type I Pre-osteocyte (Osteoblastic Osteocyte)—it is not easy to distinguish these pre-osteocytes from osteoblasts due to their similar plump shape and the abundance of cytoplasmic organelles; also, these cells have moved away from the osteoblast layer, yet they remain in direct contact with this layer and connect to the osteoblasts via numerous cell–cell connections [22,23,25],
- Type II Pre-osteocyte (Osteoid-Osteocyte)—a type II pre-osteocyte is also called a osteoid-osteocyte, which refers to a cell positioned within the osteoid seam, and furthermore, the ratio of nucleus size to cytoplasmic content is increased, cytoplasmic processes are elongated, and they continue to exhibit a bone-forming role [26],
- Type III Pre-osteocyte—type III pre-osteocytes are characterized by being completely separated from the osteogenic lamina, and they appear to lose their bone-secreting activity, which were distinctive traits of the earlier stages; in contrast to type II pre-osteocytes, the type II pre-osteocyte has a smaller cell body with a clearly defined ellipsoid shape, the ratio of nucleus size to cytoplasmic volume is higher, and the content of cytoplasmic organelles is reduced [26]
2.2.4. Communication between Bone Cells
Osteoblast-Secreted Factors
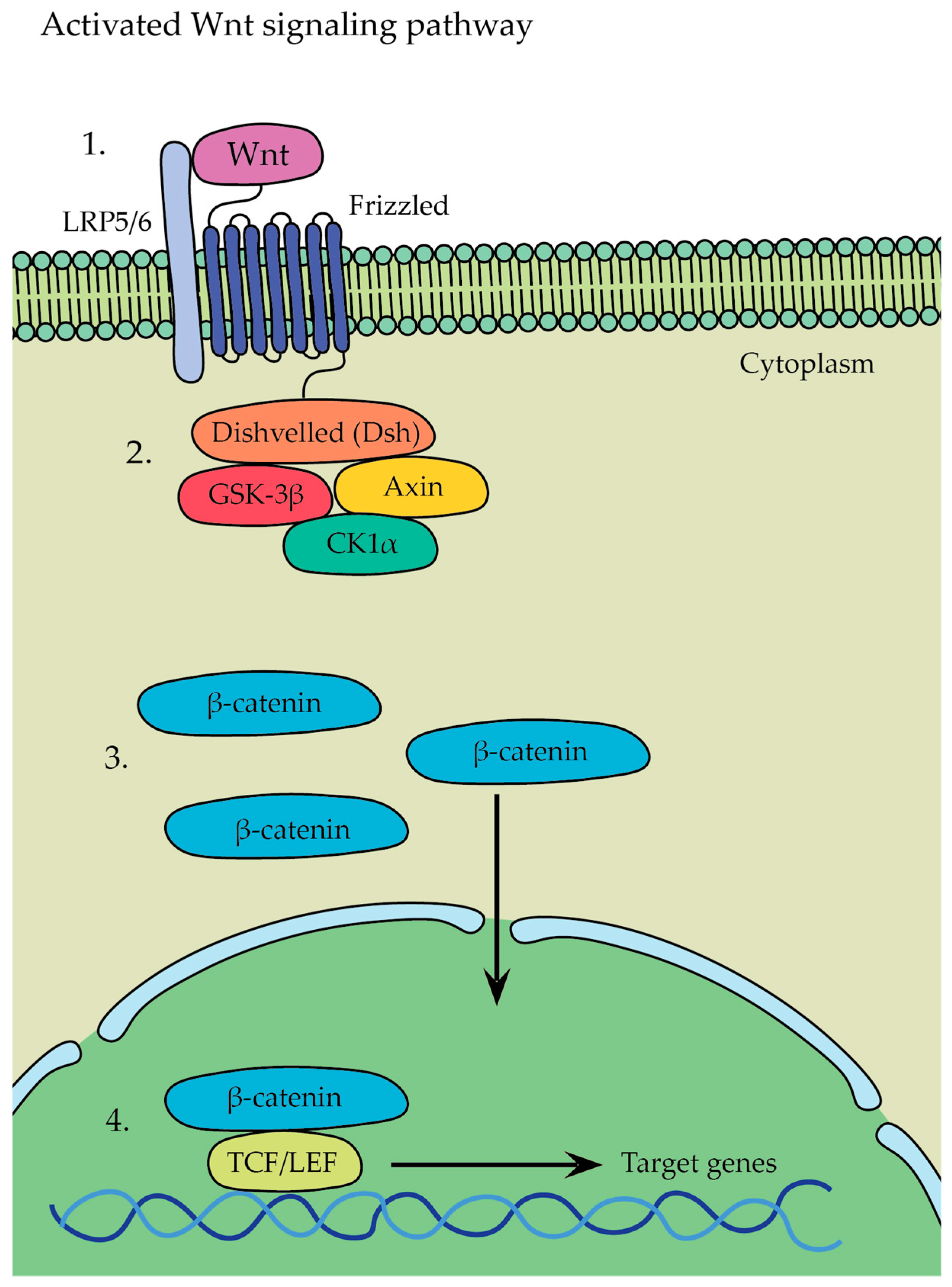
- β-catenin-dependent canonical pathway (Figure 8)—in this pathway, Wnt ligands bind to a receptor complex consisting of Frizzled and LRP5/6, which leads to the initiation of a signaling cascade where glycogen synthase kinase 3 stabilizes β-catenin → as a result, β-catenin is no longer targeted for degradation and accumulates in the cytoplasm, whereupon it translocates from there into the nucleus, leading to the activation of gene transcription program and initiation of the expression of target genes involved in various cellular processes, such as osteoblastogenesis, osteocyte formation, and bone tissue development.
- β-catenin-independent non-canonical pathway—Wnt5a, expressed in osteoblast-lineage cells, is a main ligand for non-canonical Wnt signaling that binds to its receptor complex of Frizzled and ROR2, present on the surface of osteoclasts, and it has been shown to activate both the Wnt-Ca2+ and Wnt-JNK pathways; Wnt5a was found to enhance RANKL-induced osteoclastogenesis due the upregulation of RANK expression in osteoclasts and activation of the Jun-N-terminal kinase MAPK pathway (JNK) [18,55,56].
- 1.
- Osteoclast-Secreted Factors
- 2.
- Examples of other factors influencing either osteoblast or osteoclast behavior
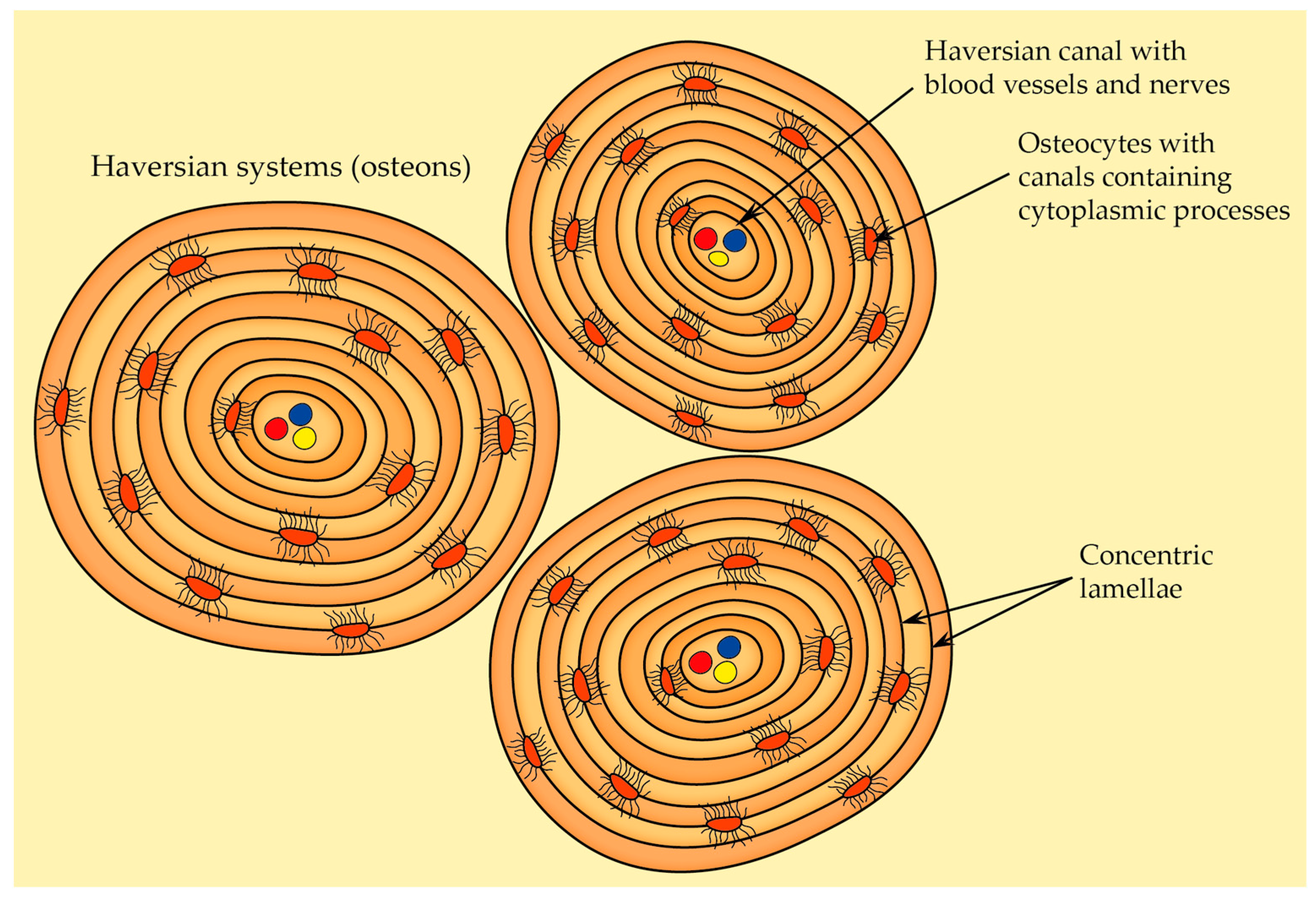
2.2.5. Osteon Structure (Haversian System)
- Primary osteons—they are situated near primary bone, and they usually contain fewer circular lamellae compared to the secondary osteons,
- Secondary osteons—also known as Haversian systems, they are the primary functional units of cortical bone that develop from primary osteons during bone remodeling. Each secondary osteon has concentric lamellae arranged around the central canal that houses blood vessels, nerves, and connective tissue, and these systems are distinguished from each other by clear cement lines that define secondary osteon boundaries.
2.2.6. Bone Blood Supply
2.3. Overview of Bone Types and Their Organization
- Stage I—the initial differentiation of pre-osteoblasts from their stem (mesenchymal) cells.
- Stage II—mesenchymal osteoblasts encircle themselves with matrix fibers oriented in a random manner.
- Stage III—the woven matrix serves as a scaffold where surface osteoblasts start synthesizing bone with a parallel-fibered lamellar structure.
- Stage IV—gradual reduction in woven bone and progression from woven to lamellar bone.
- In the trabecular bone and on the outer (periosteal) and inner (endosteal) surfaces of cortical bone, a few lamellae shape continuous circular layers that are approximately parallel to the bone surface.
- In the central regions of cortical bone, the lamellae form concentric cylinders and encircle Haversian canals, and this organization enhances the resilience of lamellar bone since the interfaces between lamellae can stop the expansion of cracks (and more energy is necessary to propagate cracks extensive enough to result in bone fractures).
2.4. Overview of Macroscopic Properties of Bone
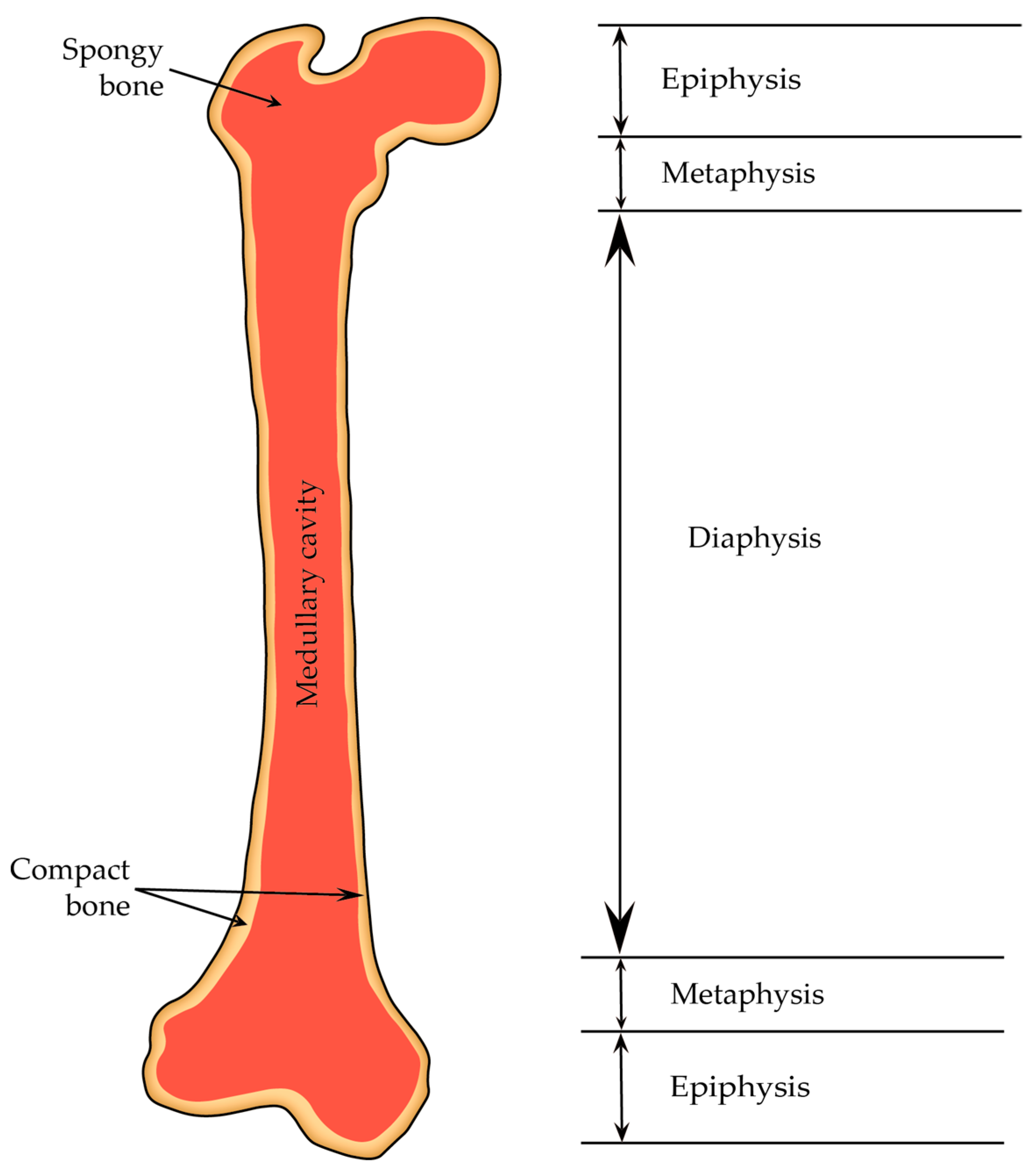
2.4.1. Diaphysis and Epiphyses
2.4.2. Periosteum and Endosteum
- Nourishment of bone tissue—blood vessels in the periosteum supply the bone’s outer surface and inner osseous tissue to some extent; these vessels are accompanied by nerves and vasomotor fibers, which control blood circulation.
- Bone growth—the periosteum is crucial for the growth of bone through surface apposition; the endosteum contributes to internal matrix formation by absorbing and depositing tissue.
- Repair—osteoprogenitor cells within the periosteum are multipotent stem cells that can prompt bone regeneration when needed, such as after a bone fracture.
- Calcium regulation—the endosteum aids in depositing calcium into the bone matrix and acts as a conduit for calcium transfer between the bone matrix and blood.
2.4.3. Bone Marrow
2.5. Bone Development and Growth
2.5.1. Intramembranous Ossification
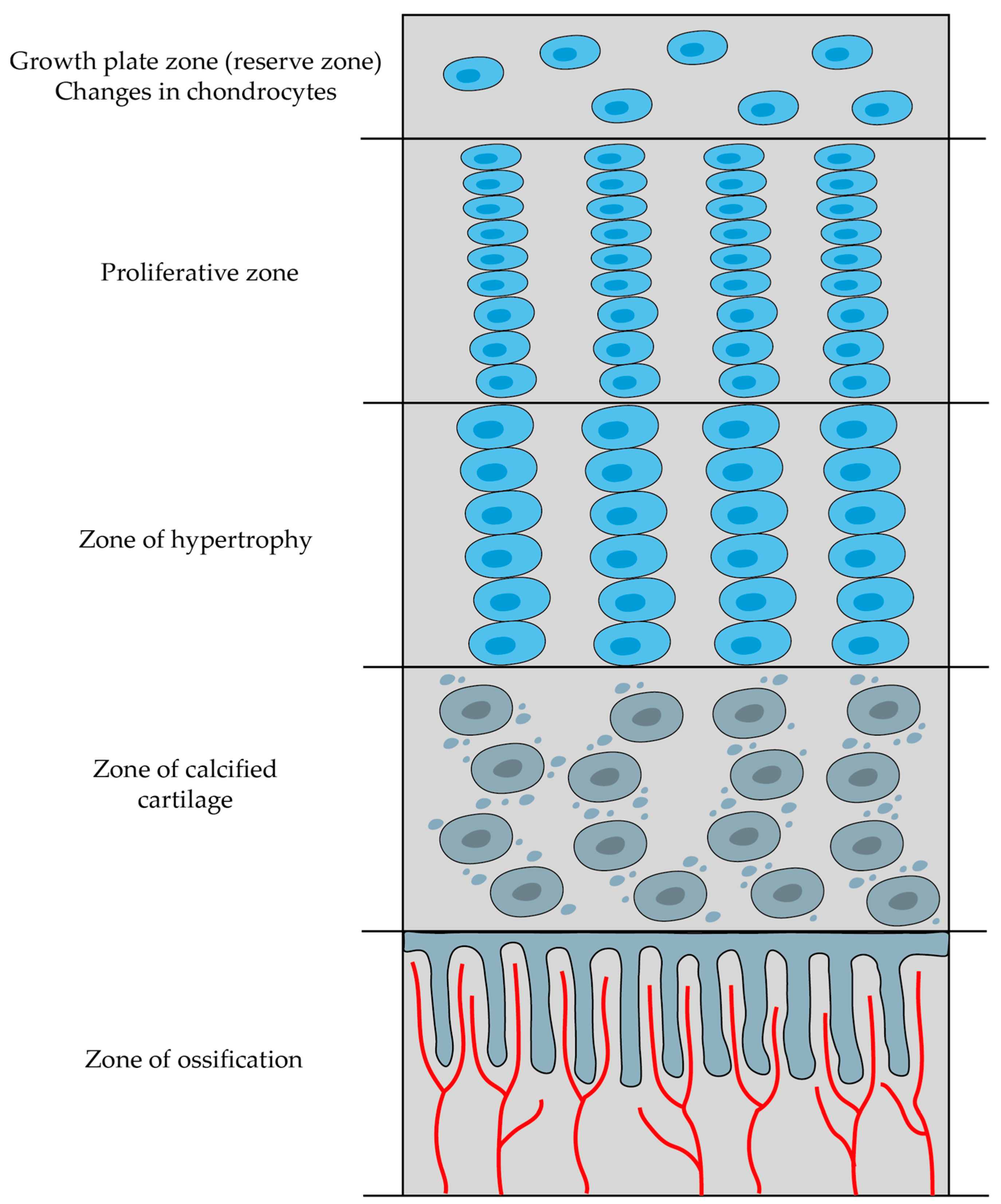
2.5.2. Endochondral Ossification
- Zone of reserve cartilage—consists of typical hyaline cartilage and serves as a reservoir of chondrocytes that can potentially contribute to growth.
- Proliferative zone—cartilage cells undergo repeated division, enlargement, and increased secretion of type II collagen and proteoglycans.
- Zone of hypertrophy—chondrocytes become enlarged and terminally differentiated; they then compress the matrix into aligned spicules and stiffen it through the secretion of type X collagen.
- Zone of calcified cartilage—chondrocytes release matrix vesicles and osteocalcin, and these substances initiate the calcification of the matrix by forming hydroxyapatite crystals.
- Zone of ossification—bone tissue first emerges in this zone; osteoblasts settle in a layer over the spicules of the calcified cartilage matrix and secrete osteoid, which subsequently transforms into woven bone (and this woven bone is later remodeled into lamellar bone).
2.5.3. Appositional Growth
2.6. Bone Remodeling
2.6.1. Bone Remodeling Cycle
Activation and Bone Resorption
Reversal and Formation Stage
2.6.2. Dynamics of Bone Remodeling
2.7. Pathophysiology of Major Bone Disorders
2.7.1. Osteoporosis
2.7.2. Osteopetrosis and Osteosclerosis
2.7.3. Hyperthyroidism and Hyperparathyroidism
2.7.4. Orthopedic Disorders
2.7.5. Paget Disease
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Standring, S. Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed.Churchill Livingstone Elsevier: London, UK, 2008. [Google Scholar]
- Bordoni, B.; Black, A.C.; Varacallo, M. Anatomy, Tendons. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA. Available online: https://www.ncbi.nlm.nih.gov/books/NBK513237 (accessed on 30 August 2023).
- U.S. Department of Health and Human Services. Bone Health and Osteoporosis: A Report of the Surgeon General; U.S. Department of Health and Human Services, Office of the Surgeon General: Rockville, MD, USA, 2004. Available online: https://www.ncbi.nlm.nih.gov/books/NBK45513 (accessed on 30 August 2023).
- Shaker, J.L.; Deftos, M.D. Calcium and Phosphate Homeostasis. Endocr. Reprod. Physiol. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279023 (accessed on 30 August 2023).
- Kovacs, E.J. Wheater’s Functional Histology: A Text and Colour Atlas. Arch. Pathol. Lab. Med. 2001, 125, 708. [Google Scholar] [CrossRef]
- Zhou, R.; Guo, Q.; Xiao, Y.; Guo, Q.; Huang, Y.; Li, C.; Luo, X. Endocrine role of bone in the regulation of energy metabolism. Bone Res. 2021, 9, 25. [Google Scholar] [CrossRef]
- Guo, Y.C.; Yuan, Q. Fibroblast growth factor 23 and bone mineralisation. Int. J. Oral Sci. 2015, 7, 8–13. [Google Scholar] [CrossRef]
- Guntur, A.R.; Rosen, C.J. Bone as an endocrine organ. Endocr. Pract. 2012, 18, 758–762. [Google Scholar] [CrossRef]
- Feng, X. Chemical and Biochemical Basis of Cell-Bone Matrix Interaction in Health and Disease. Curr. Chem. Biol. 2009, 3, 189–196. [Google Scholar] [CrossRef]
- Dalla Pria Bankoff, A. Biomechanical Characteristics of the Bone. Hum. Musculoskelet. Biomech. 2012, 61, 86. [Google Scholar]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3, S131–S139. [Google Scholar] [CrossRef]
- Mescher, L.A. Junquiera’s Basic Histology Text and Atlas, 15th ed.McGraw Hill/Medical: Chicago, IL, USA, 2018. [Google Scholar]
- Cowin, S.C.; Telega, J.J. Bone Mechanics Handbook, 2nd Edition. Appl. Mech. Rev. 2003, 56, B61–B63. [Google Scholar] [CrossRef]
- Lü, X.; Wang, J.; Li, B.; Zhang, Z.; Zhao, L. Gene expression profile study on osteoinductive effect of natural hydroxyapatite. J. Biomed. Mater. Res. Part A 2014, 102, 2833–2841. [Google Scholar] [CrossRef]
- Albrektsson, T.; Johansson, C. Osteoinduction, osteoconduction and osseointegration. Eur. Spine J. 2001, 10, S96–S101. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; Sasso, G.R.D.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. The RANKL/RANK/OPG pathway. Curr. Osteoporos. Rep. 2007, 5, 98–104. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef]
- Caetano-Lopes, J.; Canhão, H.; Fonseca, J.E. Osteoblasts and bone formation. Acta Reum. Port. 2007, 32, 103–110. [Google Scholar]
- Schaffler, M.B.; Kennedy, O.D. Osteocyte signaling in bone. Curr. Osteoporos. Rep. 2012, 10, 118–125. [Google Scholar] [CrossRef]
- Aarden, E.M.; Nijweide, P.J.; Burger, E.H. Function of osteocytes in bone. J. Cell. Biochem. 1994, 55, 287–299. [Google Scholar] [CrossRef]
- Franz-Odendaal, T.A.; Hall, B.K.; Witten, P.E. Buried alive: How osteoblasts become osteocytes. Dev. Dyn. 2006, 235, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Mullen, C.A.; Haugh, M.G.; Schaffler, M.B.; Majeska, R.J.; McNamara, L.M. Osteocyte differentiation is regulated by extracellular matrix stiffness and intercellular separation. J. Mech. Behav. Biomed. Mater. 2013, 28, 183–194. [Google Scholar] [CrossRef]
- Marotti, G.; Canè, V.; Palazzini, S.; Palumbo, C. Structure-function relationships in the osteocyte. Ital. J. Miner. Electrolyte Metab. 1990, 4, 93–106. [Google Scholar]
- Palumbo, C.; Palazzini, S.; Zaffe, D.; Marotti, G. Osteocyte differentiation in the tibia of newborn rabbit: An ultrastructural study of the formation of cytoplasmic processes. Acta Anat. 1990, 137, 350–358. [Google Scholar] [CrossRef]
- Pérez-Campo, F.M.; Santurtún, A.; García-Ibarbia, C.; Pascual, M.A.; Valero, C.; Garcés, C.; Sañudo, C.; Zarrabeitia, M.T.; Riancho, J.A. Osterix and RUNX2 are Transcriptional Regulators of Sclerostin in Human Bone. Calcif. Tissue Int. 2016, 99, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by runx2. In Osteoimmunology: Interactions of the Immune and Skeletal Systems II; Springer: Berlin/Heidelberg, Germany, 2010; Volume 658. [Google Scholar]
- Yang, F.; Tang, W.; So, S.; de Crombrugghe, B.; Zhang, C. Sclerostin is a direct target of osteoblast-specific transcription factor osterix. Biochem. Biophys. Res. Commun. 2010, 400, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, F.G.F.; Torres, J.; López-Quiles, J.; Hernández, G.; Vega, J.A.; Tresguerres, I.F. The osteocyte: A multifunctional cell within the bone. Ann. Anat. 2020, 227, 151422. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
- Wysolmerski, J.J. Osteocytic osteolysis: Time for a second look? Bonekey Rep. 2012, 1, 229. [Google Scholar] [CrossRef]
- Tazawa, K.; Hoshi, K.; Kawamoto, S.; Tanaka, M.; Ejiri, S.; Ozawa, H. Osteocytic osteolysis observed in rats to which parathyroid hormone was continuously administered. J. Bone Miner. Metab. 2004, 22, 524–529. [Google Scholar] [CrossRef]
- Neuman, W.F.; Ramp, W.K. The concept of a bone membrane: Some implications. This work was supported in part by Public Health Service Training Grant No. 1 Tl DE-175 and in part by the United States Atomic Energy Commission Contract No. At-30-1-49 and has been assigned Report No. U. Cell. Mech. Calcium Transf. Homeost. 1971, 197–209. [Google Scholar] [CrossRef]
- Klein-Nulend, J.; Bakker, A.D.; Bacabac, R.G.; Vatsa, A.; Weinbaum, S. Mechanosensation and transduction in osteocytes. Bone 2013, 54, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. Mechanosensation and transduction in osteocytes. BoneKEy-Osteovision 2006, 3, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular mechanosensors in osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Lu, Y.; Williams, E.A.; Lai, F.; Lee, J.Y.; Enishi, T.; Balani, D.H.; Ominsky, M.S.; Ke, H.Z.; Kronenberg, H.M.; et al. Sclerostin Antibody Administration Converts Bone Lining Cells Into Active Osteoblasts. J. Bone Miner. Res. 2017, 32, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.F.; Herrero, M.A.; Echeverri, L.F.; Oleaga, G.E.; López, J.M. Bone remodeling: A tissue-level process emerging from cell-level molecular algorithms. PLoS ONE 2018, 13, e0204171. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.; Heine, G.H.; Fliser, D. Clinical relevance of FGF-23 in chronic kidney disease. Kidney Int. 2009, 76, S34–S42. [Google Scholar] [CrossRef] [PubMed]
- Jüppner, H. Phosphate and FGF-23. Kidney Int. 2011, 79, S24–S27. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Jiang, Y.; Zhao, X.; Sato, T.; Densmore, M.; Schüler, C.; Erben, R.G.; McKee, M.D.; Lanske, B. Increased osteopontin contributes to inhibition of bone mineralization in FGF23-deficient mice. J. Bone Miner. Res. 2014, 29, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Song, Y.; Wu, Y.; Huang, Y.; Zhong, Q.; Zhang, Y.; Fan, Z.; Xu, C. The protective role of Ephrin-B2/EphB4 signaling in osteogenic differentiation under inflammatory environment. Exp. Cell Res. 2021, 400, 112505. [Google Scholar] [CrossRef] [PubMed]
- Vrahnas, C.; Sims, N.A. EphrinB2 Signalling in Osteoblast Differentiation, Bone Formation and Endochondral Ossification. Curr. Mol. Biol. Rep. 2015, 1, 148–156. [Google Scholar] [CrossRef]
- Li, Z.; Hao, J.; Duan, X.; Wu, N.; Zhou, Z.; Yang, F.; Li, J.; Zhao, Z.; Huang, S. The role of semaphorin 3A in bone remodeling. Front. Cell. Neurosci. 2017, 11, 40. [Google Scholar] [CrossRef]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukoc. Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef]
- Wiktor-Jedrzejczak, W.; Bartocci, A.; Ferrante, A.W.; Ahmed-Ansari, A.; Sell, K.W.; Pollard, J.W.; Stanley, E.R. Total absence of colony-stimulating factor 1 in the macrophage-deficient osteopetrotic (op/op) mouse. Proc. Natl. Acad. Sci. USA 1990, 87, 4828–4832. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K. RANK Signaling Pathways and Key Molecules Inducing Osteoclast Differentiation. Biomed. Sci. Lett. 2017, 23, 295–302. [Google Scholar] [CrossRef]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Lüthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-López, N.; Martínez-Arias, L.; Fernández-Villabrille, S.; Ruiz-Torres, M.P.; Dusso, A.; Cannata-Andía, J.B.; Naves-Díaz, M.; Panizo, S. Role of the RANK/RANKL/OPG and Wnt/β-Catenin Systems in CKD Bone and Cardiovascular Disorders. Calcif. Tissue Int. 2021, 108, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zhou, W.; Zhou, X.; Li, D.; Weng, J.; Yi, Z.; Cho, S.G.; Li, C.; Yi, T.; Wu, X.; et al. Regulation of bone formation and remodeling by G-protein-coupled receptor 48. Development 2009, 136, 2747–2756. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Z.; Ma, Y.; Yue, Z.; Lin, H.; Qu, G.; Huang, J.; Dai, W.; Li, C.; Zheng, C.; et al. LGR4 is a receptor for RANKL and negatively regulates osteoclast differentiation and bone resorption. Nat. Med. 2016, 22, 539–546. [Google Scholar] [CrossRef]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2019, 6, 170. [Google Scholar] [CrossRef]
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 signaling between osteoblast-lineage cells and osteoclast precursors enhances osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef]
- Lojk, J.; Marc, J. Roles of non-canonical wnt signalling pathways in bone biology. Int. J. Mol. Sci. 2021, 22, 10840. [Google Scholar] [CrossRef]
- Lee, S.H.; Rho, J.; Jeong, D.; Sul, J.Y.; Kim, T.; Kim, N.; Kang, J.S.; Miyamoto, T.; Suda, T.; Lee, S.K.; et al. V-ATPase V0 subunit d2-deficient mice exhibit impaired osteoclast fusion and increased bone formation. Nat. Med. 2006, 12, 1403–1409. [Google Scholar] [CrossRef]
- Lontos, K.; Adamik, J.; Tsagianni, A.; Galson, D.L.; Chirgwin, J.M.; Suvannasankha, A. The role of Semaphorin 4D in bone remodeling and cancer metastasis. Front. Endocrinol. 2018, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Kim, H.J.; Chang, E.J.; Huang, H.; Banno, Y.; Kim, H.H. Sphingosine 1-phosphate as a regulator of osteoclast differentiation and osteoclast-osteoblast coupling. EMBO J. 2006, 25, 5840–5851. [Google Scholar] [CrossRef]
- Takeshita, S.; Fumoto, T.; Matsuoka, K.; Park, K.A.; Aburatani, H.; Kato, S.; Ito, M.; Ikeda, K. Osteoclast-secreted CTHRC1 in the coupling of bone resorption to formation. J. Clin. Investig. 2013, 123, 3914–3924. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Tong, M.; Tang, B.; Shi, X. Increased level of Complement C3 in patients with osteoporosis and enhanced osteogenic ability in osteoblasts with Complement component 3 knockdown. Res. Sq. 2022, 1–15. [Google Scholar] [CrossRef]
- Lewiecki, E.M. Role of sclerostin in bone and cartilage and its potential as a therapeutic target in bone diseases. Ther. Adv. Musculoskelet. Dis. 2014, 6, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Patil, S.; Qian, A. The role of micrornas in bone metabolism and disease. Int. J. Mol. Sci. 2020, 21, 6081. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef]
- Qin, L.; Raggatt, L.J.; Partridge, N.C. Parathyroid hormone: A double-edged sword for bone metabolism. Trends Endocrinol. Metab. 2004, 15, 60–65. [Google Scholar] [CrossRef]
- Janssens, K.; Ten Dijke, P.; Janssens, S.; Van Hul, W. Transforming growth factor-β1 to the bone. Endocr. Rev. 2005, 26, 743–774. [Google Scholar] [CrossRef]
- Chang, B.; Liu, X. Osteon: Structure, Turnover, and Regeneration. Tissue Eng. Part B Rev. 2022, 28, 261–278. [Google Scholar] [CrossRef] [PubMed]
- An, Y.H.; Draughn, R.A. Mechanical Testing of Bone and the Bone-Implant Interface; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
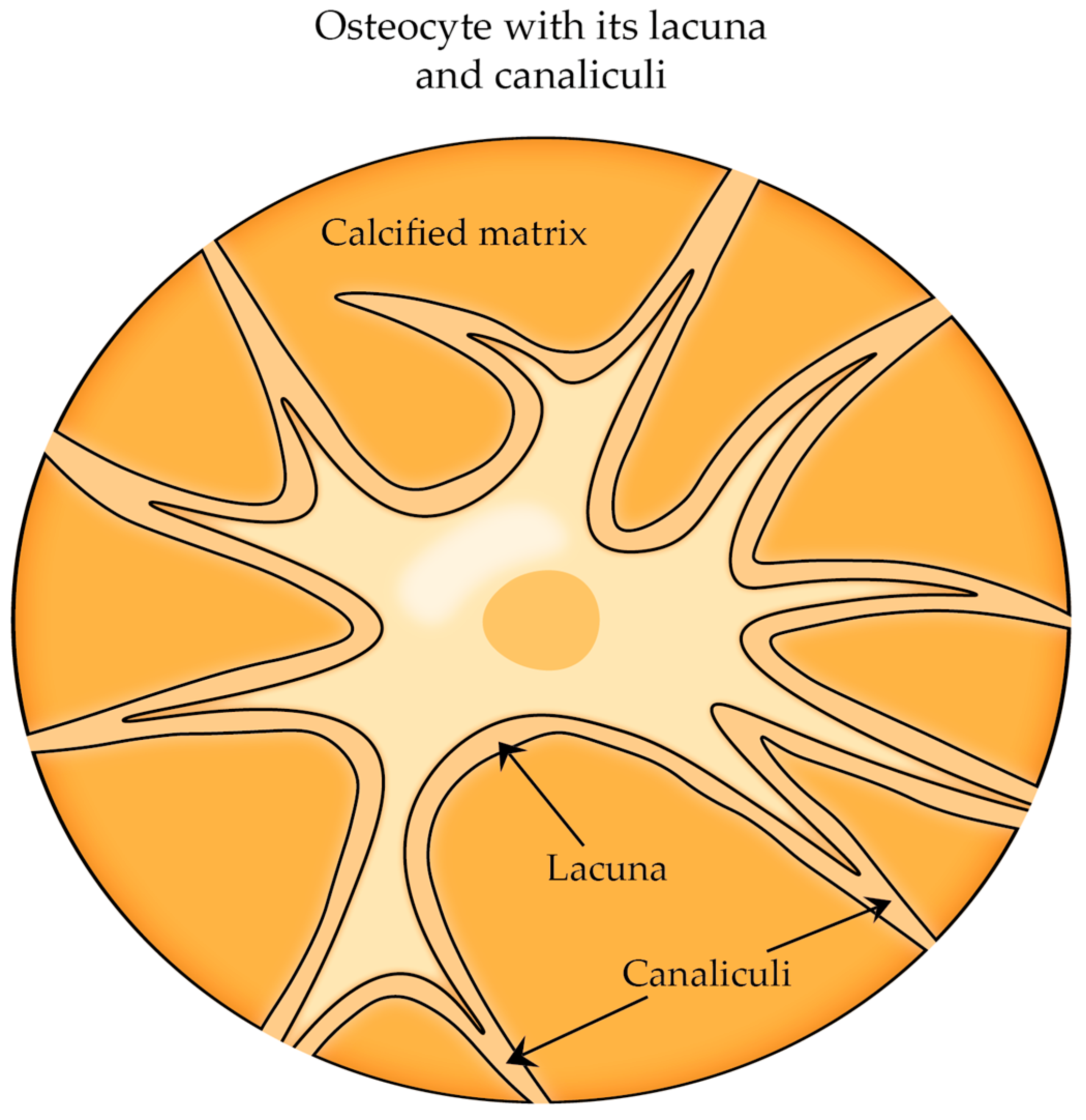
- Zhang, K.; Barragan-Adjemian, C.; Ye, L.; Kotha, S.; Dallas, M.; Lu, Y.; Zhao, S.; Harris, M.; Harris, S.E.; Feng, J.Q.; et al. E11/gp38 Selective Expression in Osteocytes: Regulation by Mechanical Strain and Role in Dendrite Elongation. Mol. Cell. Biol. 2006, 26, 4539–4552. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. Generation and function of osteocyte dendritic processes. J. Musculoskelet. Neuronal Interact. 2005, 5, 321–324. [Google Scholar]
- Milovanovic, P.; Busse, B. Inter-site Variability of the Human Osteocyte Lacunar Network: Implications for Bone Quality. Curr. Osteoporos. Rep. 2019, 17, 105–115. [Google Scholar] [CrossRef]
- Shapiro, F.; Wu, J.Y. Woven bone overview: Structural classification based on its integral role in developmental, repair and pathological bone formation throughout vertebrate groups. Eur. Cells Mater. 2019, 38, 137–167. [Google Scholar] [CrossRef] [PubMed]
- Oftadeh, R.; Perez-Viloria, M.; Villa-Camacho, J.C.; Vaziri, A.; Nazarian, A. Biomechanics and Mechanobiology of Trabecular Bone: A Review. J. Biomech. Eng. 2015, 137, 010802–01080215. [Google Scholar] [CrossRef]
- Rho, J.Y.; Kuhn-Spearing, L.; Zioupos, P. Mechanical properties and the hierarchical structure of bone. Med. Eng. Phys. 1998, 20, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.R.; Hock, J.M.; Burr, D.B. Periosteum: Biology, regulation, and response to osteoporosis therapies. Bone 2004, 35, 1003–1012. [Google Scholar] [CrossRef]
- Nahian, A.; Chauhan, P.R. Histology, Periosteum and Endosteum; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Dwek, J.R. The periosteum: What is it, where is it, and what mimics it in its absence? Skeletal Radiol. 2010, 39, 319–323. [Google Scholar] [CrossRef]
- Lucas, D. Structural organization of the bone marrow and its role in hematopoiesis. Curr. Opin. Hematol. 2021, 28, 36–42. [Google Scholar] [CrossRef]
- Breeland, G.; Sinkler, M.A.; Menezes, R.G. Embryology, Bone Ossification; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Allen, M.R.; Burr, D.B. Bone Growth, Modeling, and Remodeling. In Basic and Applied Bone Biology; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Ortega, N.; Behonick, D.J.; Werb, Z. Matrix remodeling during endochondral ossification. Trends Cell Biol. 2004, 14, 86–93. [Google Scholar] [CrossRef]
- Carter, D.R.; Van der Meulen, M.C.H.; Beaupré, G.S. Mechanical factors in bone growth and development. Bone 1996, 18, S5–S10. [Google Scholar] [CrossRef]
- Ricordeau, A.; Mellouli, N. A stochastic bone remodeling process. In Proceedings of the 2008 5th IEEE International Symposium on Biomedical Imaging: From Nano to Macro, Paris, France, 14–17 May 2008; pp. 1207–1210. [Google Scholar]
- Raggatt, L.J.; Partridge, N.C. Cellular and Molecular Mechanisms of Bone Remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.L.; Khosla, S.; Melton, L.J. A Unitary Model for Involutional Osteoporosis: Estrogen Deficiency Causes Both Type I and Type II Osteoporosis in Postmenopausal Women and Contributes to Bone Loss in Aging Men. J. Bone Miner. Res. 1998, 13, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Russell, G.; Mueller, G.; Shipman, C.; Croucher, P. Clinical Disorders of Bone Resorption. In The Molecular Basis of Skeletogenesis: Novartis Foundation Symposium; John Wiley & Sons, Ltd.: Chichester, UK, 2001; pp. 251–271. [Google Scholar]
- Eastell, R. Treatment of Postmenopausal Osteoporosis. N. Engl. J. Med. 1998, 338, 736–746. [Google Scholar] [CrossRef]
- Prestwood, M.D.K.M.; Pilbeam, M.D.C.C.; Raisz, M.D.L.G. Treatment of osteoporosis. Annu. Rev. Med. 1995, 46, 249–256. [Google Scholar] [CrossRef]
- Charles, J.M.; Key, L.L. Developmental spectrum of children with congenital osteopetrosis. J. Pediatr. 1998, 132, 371–374. [Google Scholar] [CrossRef]
- De Rosa, G.; Testa, A.; Giacomini, D.; Carrozza, C.; Astazi, P.; Caradonna, P. Prospective study of bone loss in pre- and post-menopausal women on L-thyroxine therapy for non-toxic goitre. Clin. Endocrinol. 1997, 47, 529–535. [Google Scholar] [CrossRef]
- DEMPSTER, D.W.; COSMAN, F.; PARISIEN, M.; SHEN, V.; LINDSAY, R. Anabolic Actions of Parathyroid Hormone on Bone. Endocr. Rev. 1993, 14, 690–709. [Google Scholar] [CrossRef] [PubMed]
- Kölbl, O.; Knelles, D.; Barthel, T.; Kraus, U.; Flentje, M.; Eulert, J. Randomized trial comparing early postoperative irradiation vs. the use of nonsteroidal antiinflammatory drugs for prevention of heterotopic ossification following prosthetic total hip replacement. Int. J. Radiat. Oncol. 1997, 39, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, M. Cellular basis for failure of joint prosthesis. Biomed. Mater. Eng. 1996, 6, 165–172. [Google Scholar] [CrossRef] [PubMed]
- MEE, A.; DIXON, J.; HOYLAND, J.; DAVIES, M.; SELBY, P.; MAWER, E. Detection of canine distemper virus in 100% of Paget’s disease samples by in situ-reverse transcriptase-polymerase chain reaction. Bone 1998, 23, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Nellissery, M.J.; Padalecki, S.S.; Brkanac, Z.; Singer, F.R.; Roodman, G.D.; Unni, K.K.; Leach, R.J.; Hansen, M.F. Evidence for a Novel Osteosarcoma Tumor-Suppressor Gene in the Chromosome 18 Region Genetically Linked with Paget Disease of Bone. Am. J. Hum. Genet. 1998, 63, 817–824. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šromová, V.; Sobola, D.; Kaspar, P. A Brief Review of Bone Cell Function and Importance. Cells 2023, 12, 2576. https://doi.org/10.3390/cells12212576
Šromová V, Sobola D, Kaspar P. A Brief Review of Bone Cell Function and Importance. Cells. 2023; 12(21):2576. https://doi.org/10.3390/cells12212576
Chicago/Turabian StyleŠromová, Veronika, Dinara Sobola, and Pavel Kaspar. 2023. "A Brief Review of Bone Cell Function and Importance" Cells 12, no. 21: 2576. https://doi.org/10.3390/cells12212576