
Astrocytes Undergo Metabolic Reprogramming in the Multiple Sclerosis Animal Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and EAE Induction
2.2. Astrocyte Isolation
2.3. Gene Expression Analysis by RNA-Sequencing
2.4. Gene Expression Analysis by qRT-PCR
2.5. GFAP Immunofluorescence and 3-Dimensional Reconstruction of Astrocytes
2.6. Statistical Analysis
3. Results
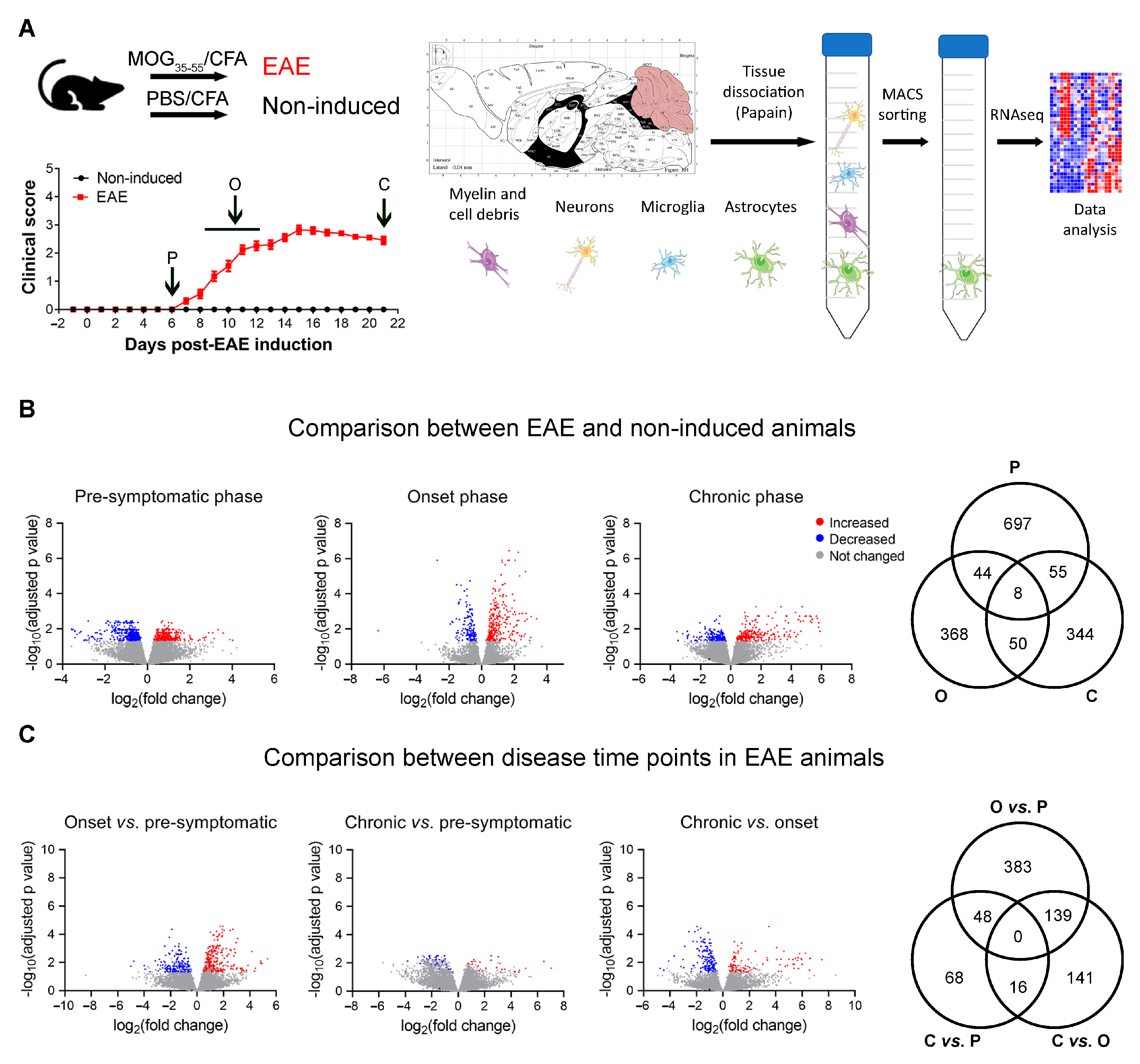
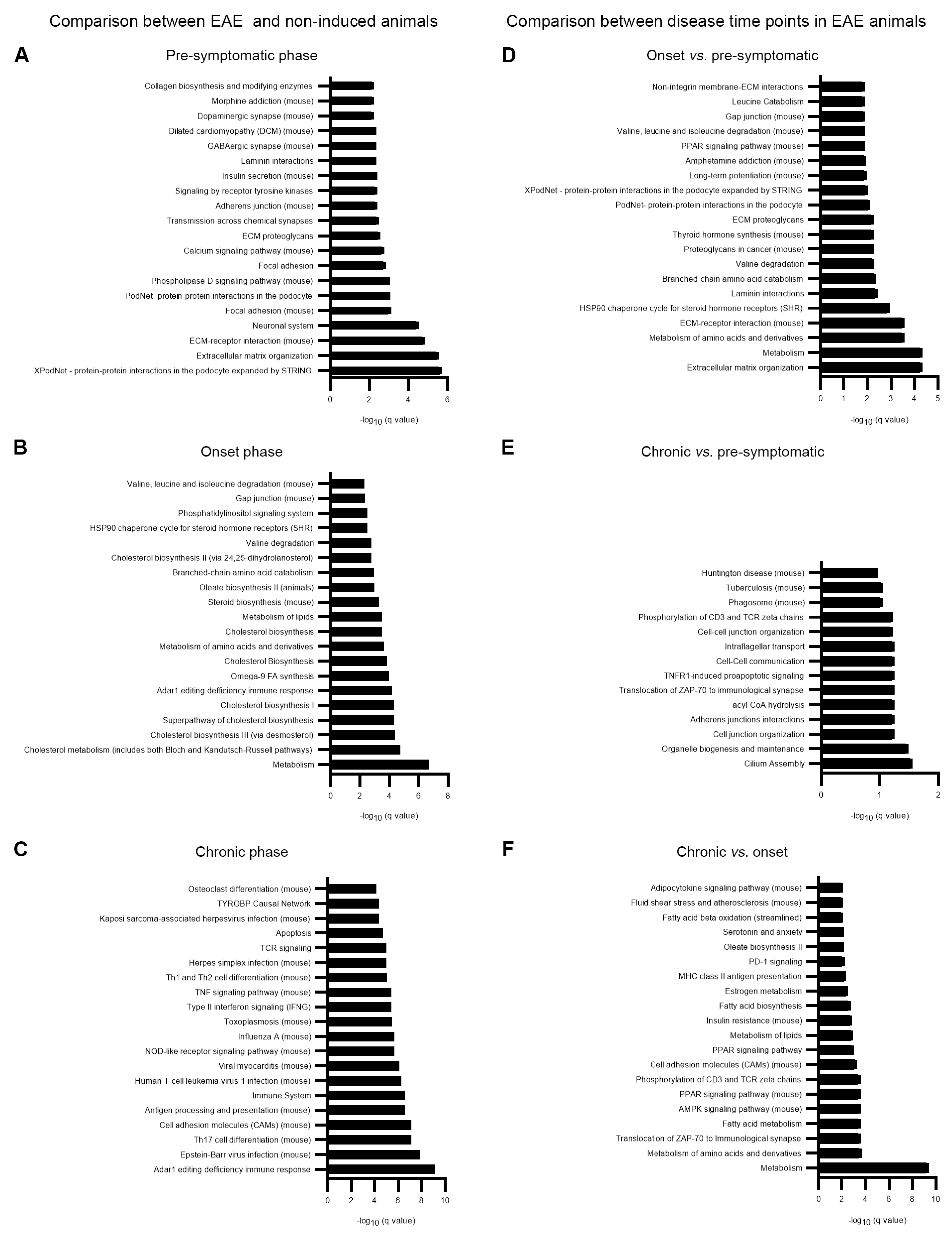
3.1. Altered Astrocytic Gene Expression in EAE Animals
3.2. Astrocytic Metabolic Reprogramming at the Onset Phase of Disease
3.3. Increased Astrocytic Length and Complexity in EAE Animals near Lesion Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lassmann, H. Multiple Sclerosis Pathology. Cold Spring Harb. Perspect. Med. 2018, 8, a028936. [Google Scholar] [CrossRef] [PubMed]
- Freedman, M.S. Multiple sclerosis: Is there a safe time to discontinue therapy in MS? Nat. Rev. Neurol. 2016, 13, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H.; van Horssen, J.; Mahad, D. Progressive multiple sclerosis: Pathology and pathogenesis. Nat. Rev. Neurol. 2012, 8, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Sochocka, M.; Diniz, B.S.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Marques, F.; Mesquita, S.D.; Sousa, J.C.; Coppola, G.; Gao, F.; Geschwind, D.H.; Columba-Cabezas, S.; Aloisi, F.; Degn, M.; Cerqueira, J.J.; et al. Lipocalin 2 is present in the EAE brain and is modulated by natalizumab. Front. Cell. Neurosci. 2012, 6, 33. [Google Scholar] [CrossRef]
- Slavin, A.; Ewing, C.; Liu, J.; Ichikawa, M.; Slavin, J.; Bernard, C.C. Induction of a multiple sclerosis-like disease in mice with an immunodominant epitope of myelin oligodendrocyte glycoprotein. Autoimmunity 1998, 28, 109–120. [Google Scholar] [CrossRef]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Ann. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef]
- Batoulis, H.; Recks, M.S.; Addicks, K.; Kuerten, S. Experimental autoimmune encephalomyelitis—Achievements and prospective advances. APMIS 2011, 119, 819–830. [Google Scholar] [CrossRef]
- Stromnes, I.M.; Goverman, J.M. Active induction of experimental allergic encephalomyelitis. Nat. Protoc. 2006, 1, 1810–1819. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, C.S.; Farooqi, N.; O’Brien, K.; Gran, B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharmacol. 2011, 164, 1079–1106. [Google Scholar] [CrossRef] [PubMed]
- Kuerten, S.; Kostova-Bales, D.A.; Frenzel, L.P.; Tigno, J.T.; Tary-Lehmann, M.; Angelov, D.N.; Lehmann, P.V. MP4- and MOG:35-55-induced EAE in C57BL/6 mice differentially targets brain, spinal cord and cerebellum. J. Neuroimmunol. 2007, 189, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. Animal models of multiple sclerosis: The good, the bad and the bottom line. Nat. Neurosci. 2012, 15, 1074–1077. [Google Scholar] [CrossRef]
- Wiedrick, J.; Meza-Romero, R.; Gerstner, G.; Seifert, H.; Chaudhary, P.; Headrick, A.; Kent, G.; Maestas, A.; Offner, H.; Vandenbark, A.A. Sex differences in EAE reveal common and distinct cellular and molecular components. Cell Immunol. 2021, 359, 104242. [Google Scholar] [CrossRef]
- Holt, L.M.; Olsen, M.L. Novel Applications of Magnetic Cell Sorting to Analyze Cell-Type Specific Gene and Protein Expression in the Central Nervous System. PLoS ONE 2016, 11, e0150290. [Google Scholar] [CrossRef]
- Kamburov, A.; Pentchev, K.; Galicka, H.; Wierling, C.; Lehrach, H.; Herwig, R. ConsensusPathDB: Toward a more complete picture of cell biology. Nucleic Acids Res. 2011, 39, D712–D717. [Google Scholar] [CrossRef]
- Kamburov, A.; Wierling, C.; Lehrach, H.; Herwig, R. ConsensusPathDB--a database for integrating human functional interaction networks. Nucleic Acids Res. 2009, 37, D623–D628. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Longair, M.H.; Baker, D.A.; Armstrong, J.D. Simple Neurite Tracer: Open source software for reconstruction, visualization and analysis of neuronal processes. Bioinformatics 2011, 27, 2453–2454. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Tavares, G.; Martins, M.; Correia, J.S.; Sardinha, V.M.; Guerra-Gomes, S.; das Neves, S.P.; Marques, F.; Sousa, N.; Oliveira, J.F. Employing an open-source tool to assess astrocyte tridimensional structure. Brain Struct. Funct. 2017, 222, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [PubMed]
- Kantzer, C.G.; Boutin, C.; Herzig, I.D.; Wittwer, C.; Reiss, S.; Tiveron, M.C.; Drewes, J.; Rockel, T.D.; Ohlig, S.; Ninkovic, J.; et al. Anti-ACSA-2 defines a novel monoclonal antibody for prospective isolation of living neonatal and adult astrocytes. Glia 2017, 65, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Batiuk, M.Y.; de Vin, F.; Duque, S.I.; Li, C.; Saito, T.; Saido, T.; Fiers, M.; Belgard, T.G.; Holt, M.G. An immunoaffinity-based method for isolating ultrapure adult astrocytes based on ATP1B2 targeting by the ACSA-2 antibody. J. Biol. Chem. 2017, 292, 8874–8891. [Google Scholar] [CrossRef]
- Hofstetter, H.H.; Shive, C.L.; Forsthuber, T.G. Pertussis toxin modulates the immune response to neuroantigens injected in incomplete Freund’s adjuvant: Induction of Th1 cells and experimental autoimmune encephalomyelitis in the presence of high frequencies of Th2 cells. J. Immunol. 2002, 169, 117–125. [Google Scholar] [CrossRef]
- Turley, D.M.; Miller, S.D. Peripheral tolerance induction using ethylenecarbodiimide-fixed APCs uses both direct and indirect mechanisms of antigen presentation for prevention of experimental autoimmune encephalomyelitis. J. Immunol. 2007, 178, 2212–2220. [Google Scholar] [CrossRef]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Allen, N.J.; Barres, B.A. Neuroscience: Glia—More than just brain glue. Nature 2009, 457, 675–677. [Google Scholar] [CrossRef]
- Wekerle, H.; Kurschus, F.C. Animal models of multiple sclerosis. Drug Discov. Today Dis. Models 2006, 3, 359–367. [Google Scholar] [CrossRef]
- Kuerten, S.; Lehmann, P.V. The immune pathogenesis of experimental autoimmune encephalomyelitis: Lessons learned for multiple sclerosis? J. Interferon Cytokine Res. 2011, 31, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Galvan-Pena, S.; O’Neill, L.A. Metabolic reprograming in macrophage polarization. Front. Immunol. 2014, 5, 420. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Gutierrez-Vazquez, C.; Rothhammer, V.; Mayo, L.; Wheeler, M.A.; Tjon, E.C.; Zandee, S.E.J.; Blain, M.; de Lima, K.A.; Takenaka, M.C.; et al. Metabolic Control of Astrocyte Pathogenic Activity via cPLA2-MAVS. Cell 2019, 179, 1483–1498.e1422. [Google Scholar] [CrossRef]
- Mayo, L.; Trauger, S.A.; Blain, M.; Nadeau, M.; Patel, B.; Alvarez, J.I.; Mascanfroni, I.D.; Yeste, A.; Kivisakk, P.; Kallas, K.; et al. Regulation of astrocyte activation by glycolipids drives chronic CNS inflammation. Nat. Med. 2014, 20, 1147–1156. [Google Scholar] [CrossRef]
- Mueller, A.M.; Pedre, X.; Stempfl, T.; Kleiter, I.; Couillard-Despres, S.; Aigner, L.; Giegerich, G.; Steinbrecher, A. Novel role for SLPI in MOG-induced EAE revealed by spinal cord expression analysis. J. Neuroinflamm. 2008, 5, 20. [Google Scholar] [CrossRef]
- Itoh, N.; Itoh, Y.; Tassoni, A.; Ren, E.; Kaito, M.; Ohno, A.; Ao, Y.; Farkhondeh, V.; Johnsonbaugh, H.; Burda, J.; et al. Cell-specific and region-specific transcriptomics in the multiple sclerosis model: Focus on astrocytes. Proc. Natl. Acad. Sci. USA 2018, 115, E302–E309. [Google Scholar] [CrossRef]
- Tassoni, A.; Farkhondeh, V.; Itoh, Y.; Itoh, N.; Sofroniew, M.V.; Voskuhl, R.R. The astrocyte transcriptome in EAE optic neuritis shows complement activation and reveals a sex difference in astrocytic C3 expression. Sci. Rep. 2019, 9, 10010. [Google Scholar] [CrossRef]
- Sevastou, I.; Pryce, G.; Baker, D.; Selwood, D.L. Characterisation of Transcriptional Changes in the Spinal Cord of the Progressive Experimental Autoimmune Encephalomyelitis Biozzi ABH Mouse Model by RNA Sequencing. PLoS ONE 2016, 11, e0157754. [Google Scholar] [CrossRef]
- Khadem, M.D.; Tabandeh, M.R.; Haschemi, A.; Kheirollah, A.; Shahriari, A. Dimethyl itaconate reprograms neurotoxic to neuroprotective primary astrocytes through the regulation of NLRP3 inflammasome and NRF2/HO-1 pathways. Mol. Cell. Neurosci. 2022, 122, 103758. [Google Scholar] [CrossRef]
- Robb, J.L.; Hammad, N.A.; Weightman Potter, P.G.; Chilton, J.K.; Beall, C.; Ellacott, K.L.J. The metabolic response to inflammation in astrocytes is regulated by nuclear factor-kappa B signaling. Glia 2020, 68, 2246–2263. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.V.; McKeown, S.R. A histological, histochemical and biochemical study of the macroscopically normal white matter in multiple sclerosis. J. Neurol. Sci. 1979, 41, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Graumann, U.; Reynolds, R.; Steck, A.J.; Schaeren-Wiemers, N. Molecular changes in normal appearing white matter in multiple sclerosis are characteristic of neuroprotective mechanisms against hypoxic insult. Brain Pathol. 2003, 13, 554–573. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Time Point | Interaction | |||||||
|---|---|---|---|---|---|---|---|---|---|
| F Value | p Value | Effect Size | F Value | p Value | Effect Size | F Value | p Value | Effect Size | |
| Pfkm | F(1,22) = 0.0212 | 0.8854 | F(2,22) = 4.858 | 0.0179 | η2 = 0.297 | F(2,22) = 0.4698 | 0.6312 | ||
| Aldoc | F(1,22) = 0.3990 | 0.5341 | F(2,22) = 2.832 | 0.0805 | F(2,22) = 4.161 | 0.0293 | η2 = 0.229 | ||
| Idh3g | F(1,22) = 6.639 | 0.0172 | η2 = 0.089 | F(2,22) = 17.140 | <0.0001 | η2 = 0.462 | F(2,22) = 5.664 | 0.0104 | η2 = 0.153 |
| Sdha | F(1,22) = 0.9211 | 0.3476 | F(2,22) = 5.358 | 0.0127 | η2 = 0.309 | F(2,22) = 0.5214 | 0.6009 | ||
| Fbln5 | F(1,22) = 21.370 | 0.0001 | η2 = 0.214 | F(2,22) = 12.670 | 0.0002 | η2 = 0.253 | F(2,22) = 15.670 | <0.0001 | η2 = 0.313 |
| Amigo2 | F(1,22) = 15.070 | 0.0008 | η2 = 0.209 | F(2,22) = 10.150 | 0.0008 | η2 = 0.281 | F(2,22) = 7.406 | 0.0035 | η2 = 0.205 |
| Fkbp5 | F(1,22) = 34.860 | <0.0001 | η2 = 0.326 | F(2,22) = 9.893 | 0.0009 | η2 = 0.185 | F(2,22) = 15.170 | <0.0001 | η2 = 0.284 |
| Disease | Time Point | Interaction | |||||||
|---|---|---|---|---|---|---|---|---|---|
| F Value | p Value | Effect Size | F Value | p Value | Effect Size | F Value | p Value | Effect Size | |
| Total length NAWM in EAE vs. non-induced | F(1,21) = 1.158 | 0.2941 | F(2,21) = 0.174 | 0.8415 | F(2,21) = 0.637 | 0.5389 | |||
| Total length lesion in EAE vs. non-induced | F(1,14) = 9.548 | 0.0080 | ηp2 = 0.405 | F(1,14) = 0.243 | 0.6297 | F(1,14) = 0.149 | 0.7050 | ||
| White matter region | Time point | Interaction | |||||||
| Total length lesion in EAE vs. NAWM in EAE | F(1,8) = 5.214 | 0.0518 | ηp2 = 0.395 | F(1,8) = 0.182 | 0.6809 | F(1,8) = 0.821 | 0.3915 | ||
| Interaction | ||||||||
|---|---|---|---|---|---|---|---|---|
| F Value | p Value | Effect Size | F Value | p Value | Effect Size | |||
| NAWM in EAE vs. non-induced | Disease | F(1,21) = 0.939 | 0.3435 | radius*disease | F(17,357) = 1.049 | 0.4034 | ||
| Time point | F(2,21) = 0.159 | 0.8544 | radius* time point | F(34,357) = 0.479 | 0.9946 | |||
| Radius | F(17,357) = 354.805 | <0.0001 | ηp2 = 0.944 | radius*disease* time point | F(34,357) = 0.793 | 0.7920 | ||
| disease* time point | F(2,21) = 0.762 | 0.4794 | ||||||
| Lesion in EAE vs. non-induced | Disease | F(1,14) = 9.201 | 0.0089 | radius*disease | F(15,210) = 4.685 | <0.0001 | ηp2 = 0.251 | |
| Time point | ηp2 = 0.397 | F(1,14) = 0.104 | 0.7524 | radius* time point | F(15,210) = 0.903 | 0.5613 | ||
| Radius | F(15,210) = 330.458 | <0.0001 | ηp2 = 0.959 | radius*disease* time point | F(15,210) = 1.061 | 0.3949 | ||
| disease* time point | F(1,14) = 0.490 | 0.4952 | ||||||
| Lesion in EAE vs. non-induced | White matter region | F(1,8) = 4.418 | 0.0687 | radius*region | F(17,136) = 2.991 | 0.0002 | ηp2 = 0.272 | |
| Time point | F(1,8) = 0.426 | 0.5321 | radius* time point | F(17,136) = 0.299 | 0.9969 | |||
| Radius | F(17,136) = 153.157 | <0.0001 | ηp2 = 0.950 | radius*region* time point | F(17,136) = 1.673 | 0.0550 | ||
| region* time point | F(1,8) = 0.548 | 0.4802 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
das Neves, S.P.; Sousa, J.C.; Magalhães, R.; Gao, F.; Coppola, G.; Mériaux, S.; Boumezbeur, F.; Sousa, N.; Cerqueira, J.J.; Marques, F. Astrocytes Undergo Metabolic Reprogramming in the Multiple Sclerosis Animal Model. Cells 2023, 12, 2484. https://doi.org/10.3390/cells12202484
das Neves SP, Sousa JC, Magalhães R, Gao F, Coppola G, Mériaux S, Boumezbeur F, Sousa N, Cerqueira JJ, Marques F. Astrocytes Undergo Metabolic Reprogramming in the Multiple Sclerosis Animal Model. Cells. 2023; 12(20):2484. https://doi.org/10.3390/cells12202484
Chicago/Turabian Styledas Neves, Sofia Pereira, João Carlos Sousa, Ricardo Magalhães, Fuying Gao, Giovanni Coppola, Sebatien Mériaux, Fawzi Boumezbeur, Nuno Sousa, João José Cerqueira, and Fernanda Marques. 2023. "Astrocytes Undergo Metabolic Reprogramming in the Multiple Sclerosis Animal Model" Cells 12, no. 20: 2484. https://doi.org/10.3390/cells12202484