
The Chemical Inhibitors of Endocytosis: From Mechanisms to Potential Clinical Applications
 ,
,  , , ,
, , ,  ,
,  ,
,  and
and
Abstract
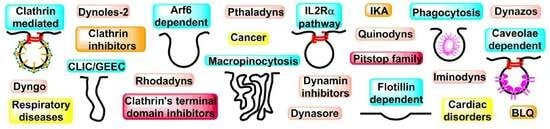
:
1. Introduction
2. Mechanisms of Endocytosis
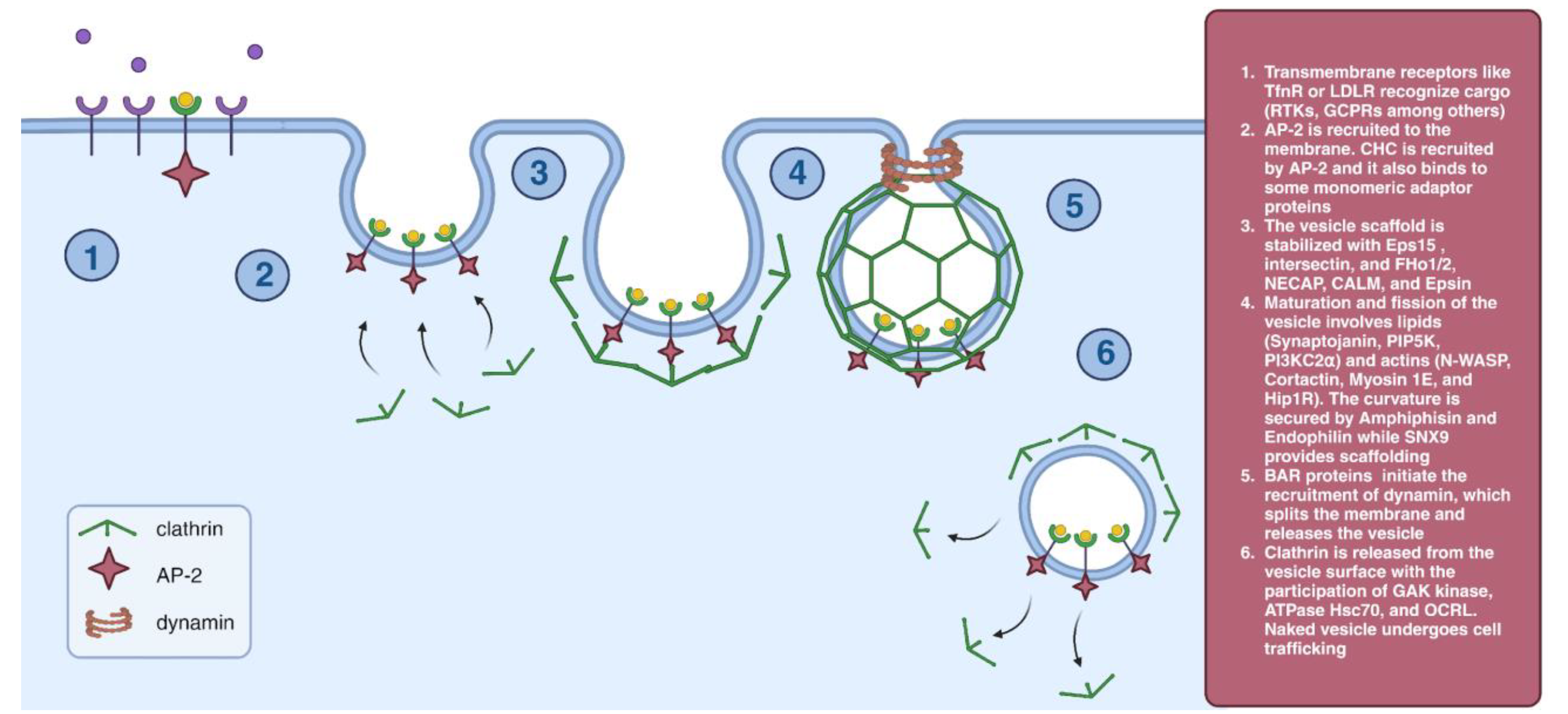
2.1. Clathrin-Mediated Endocytosis (CME)
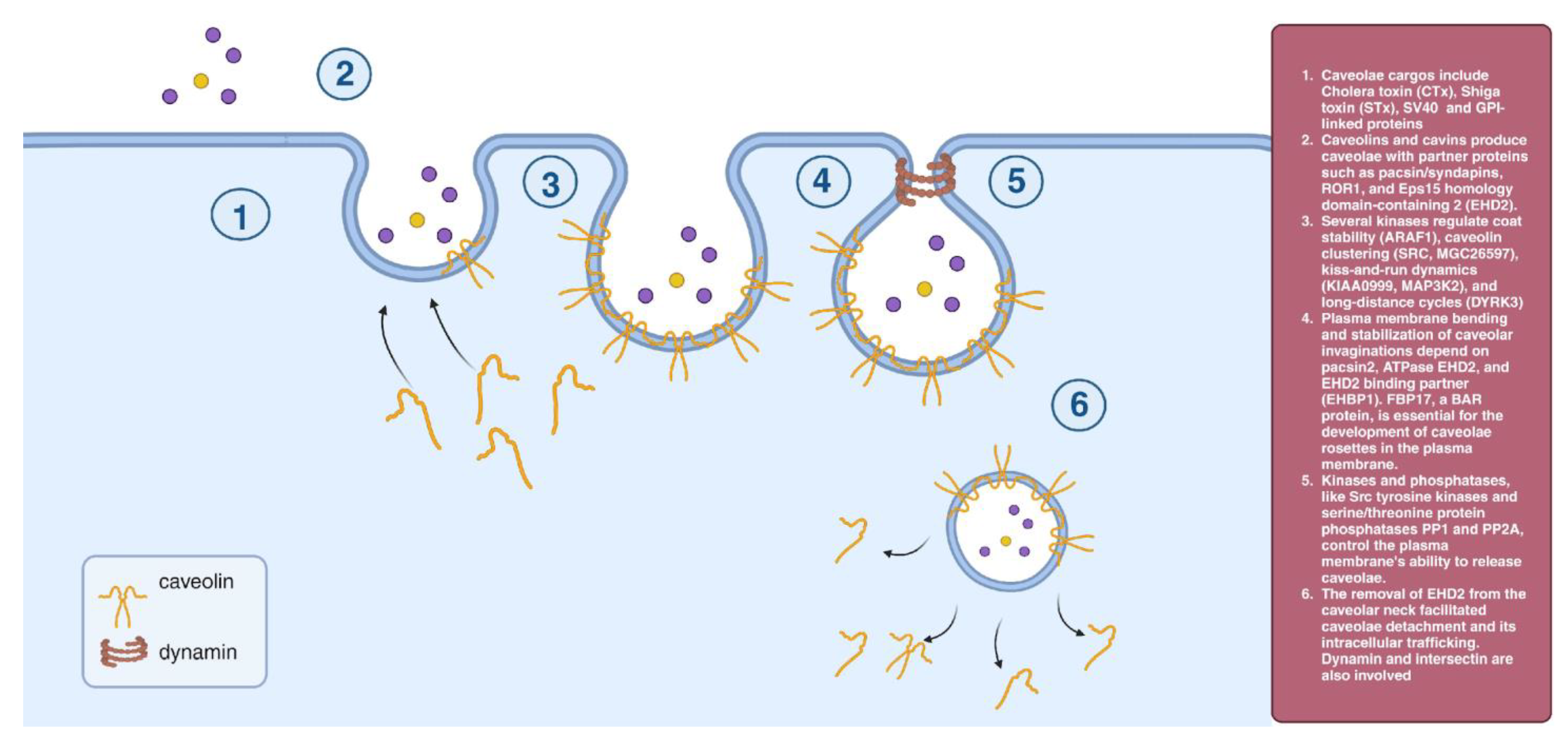
2.2. Caveolae-Dependent Endocytosis
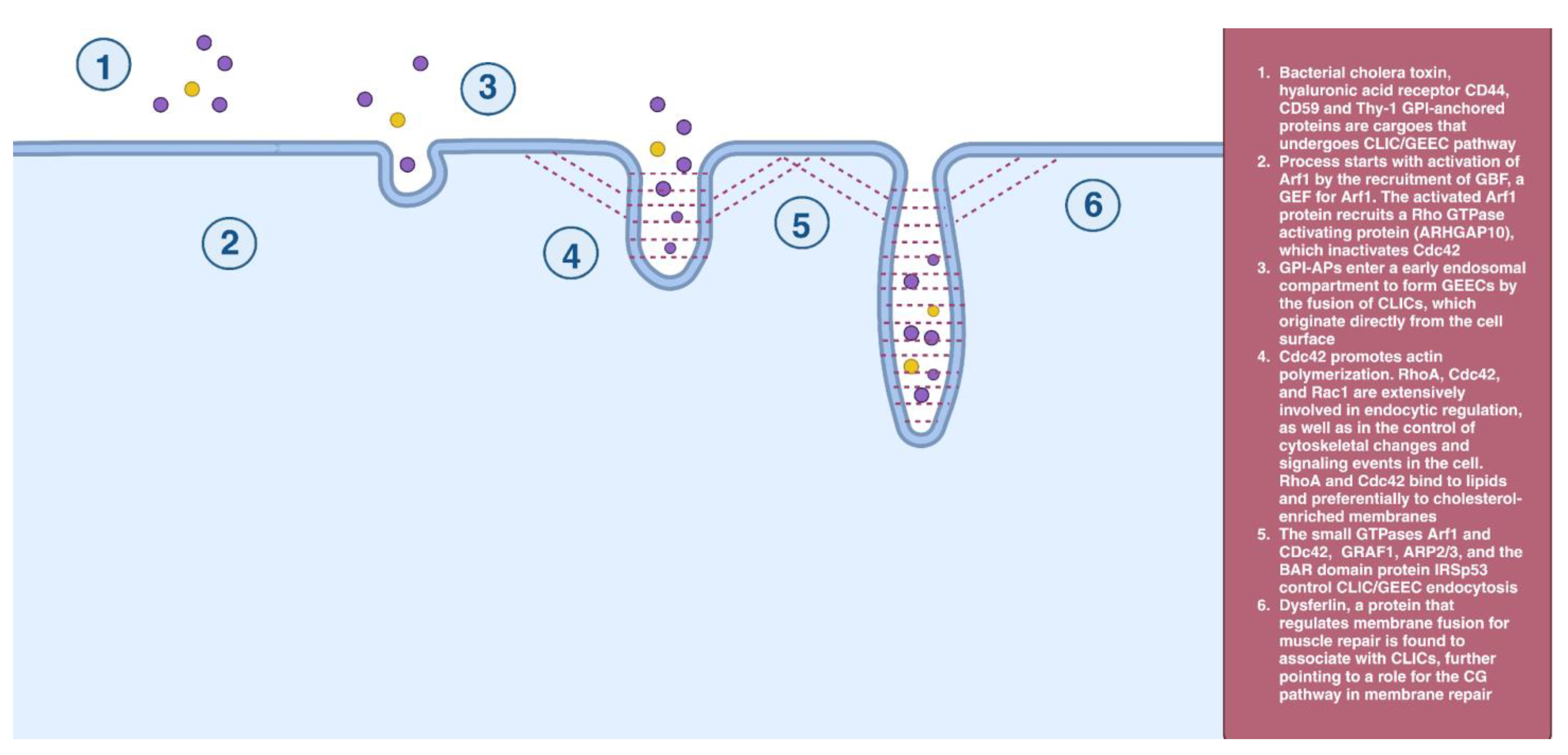
2.3. CLIC/GEEC Endocytosis
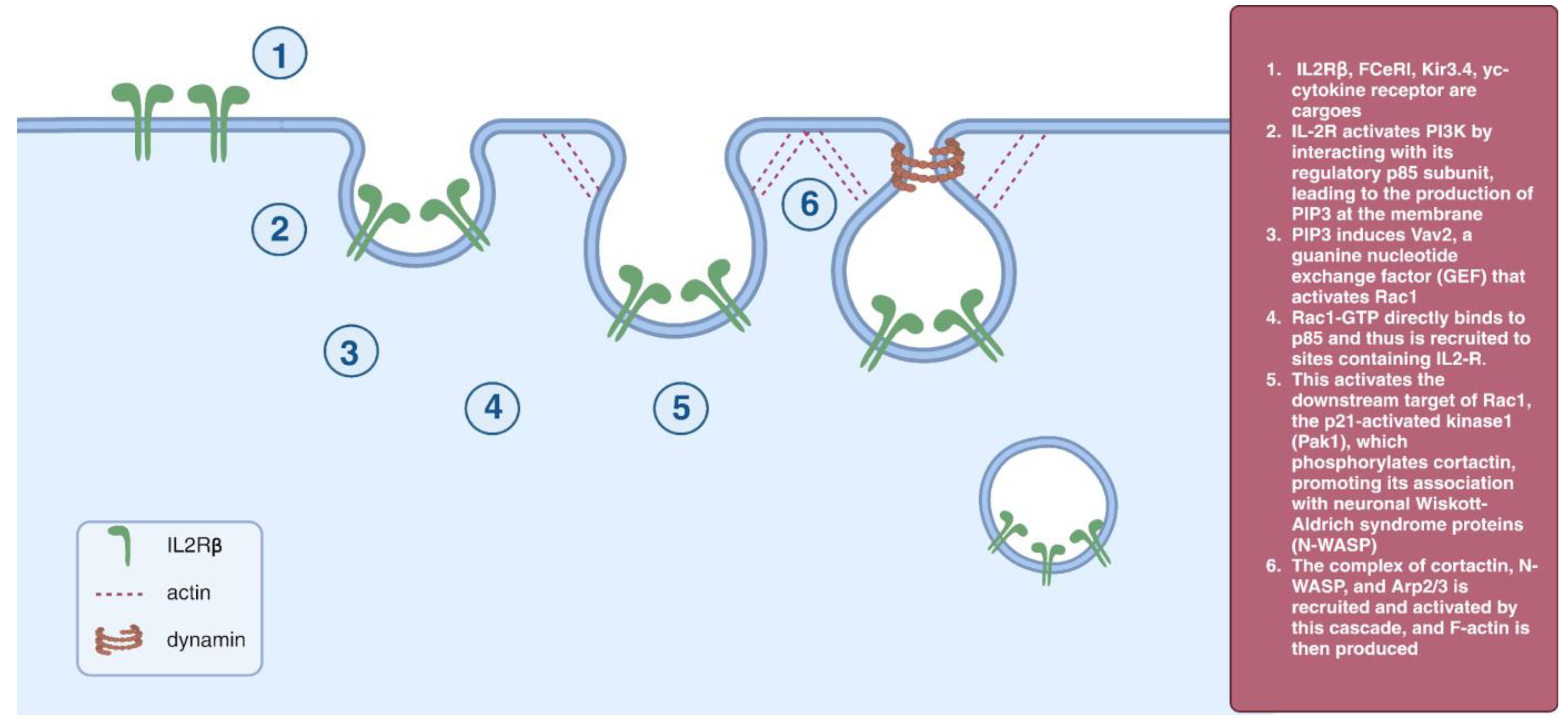
2.4. IL2Rβ Pathway
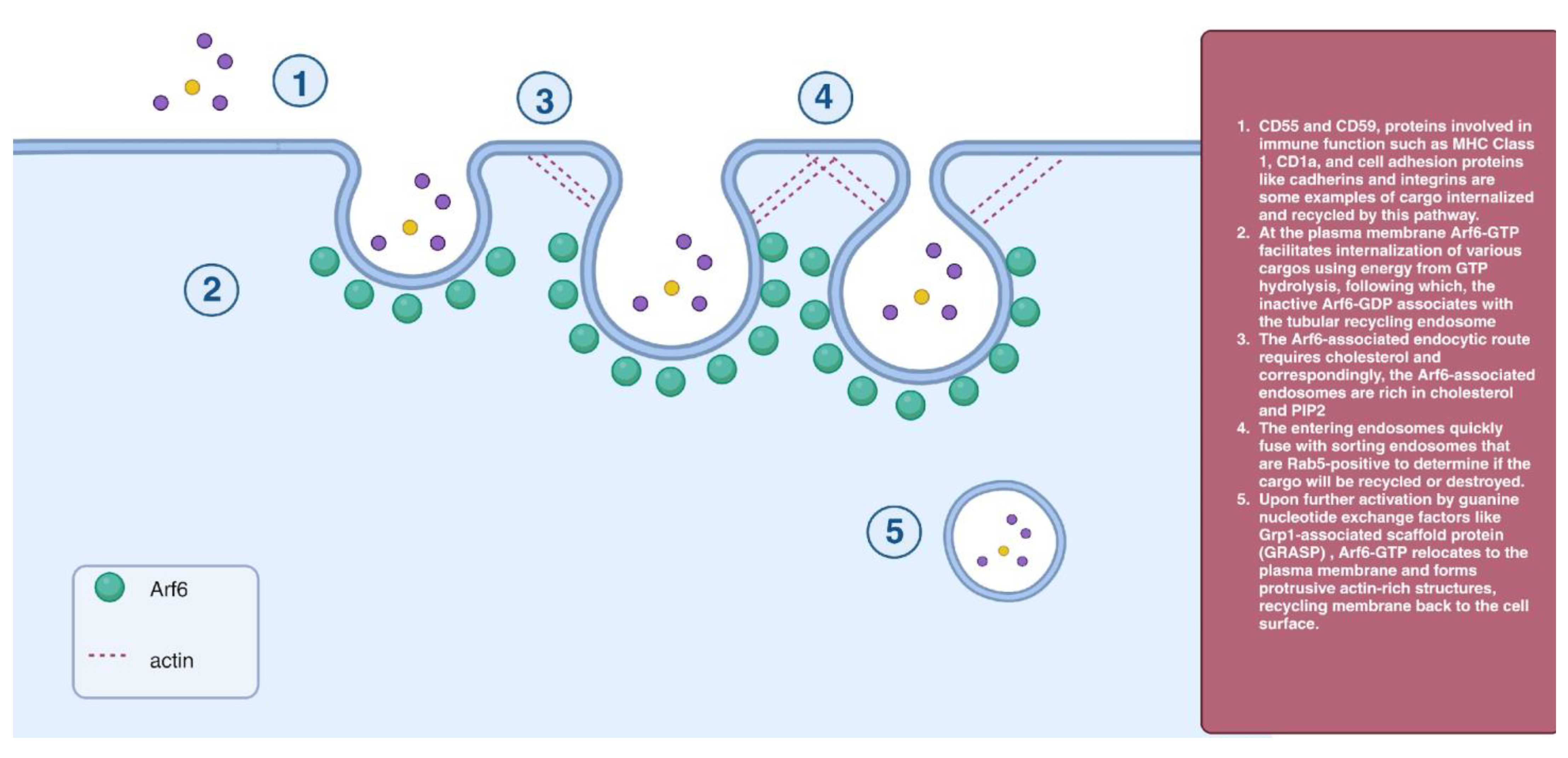
2.5. Arf6-Dependent Endocytosis
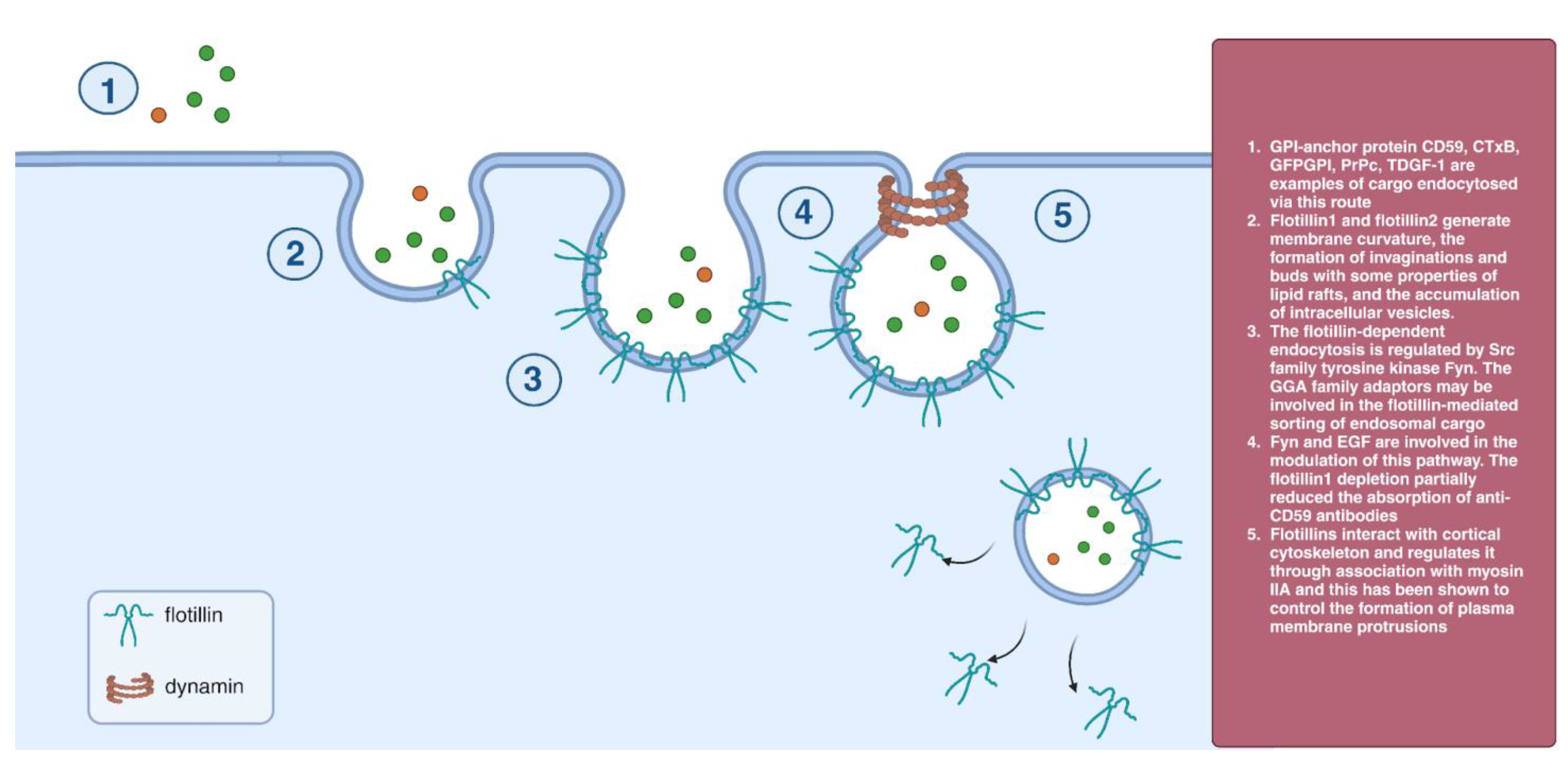
2.6. Flotillin-Dependent Endocytosis
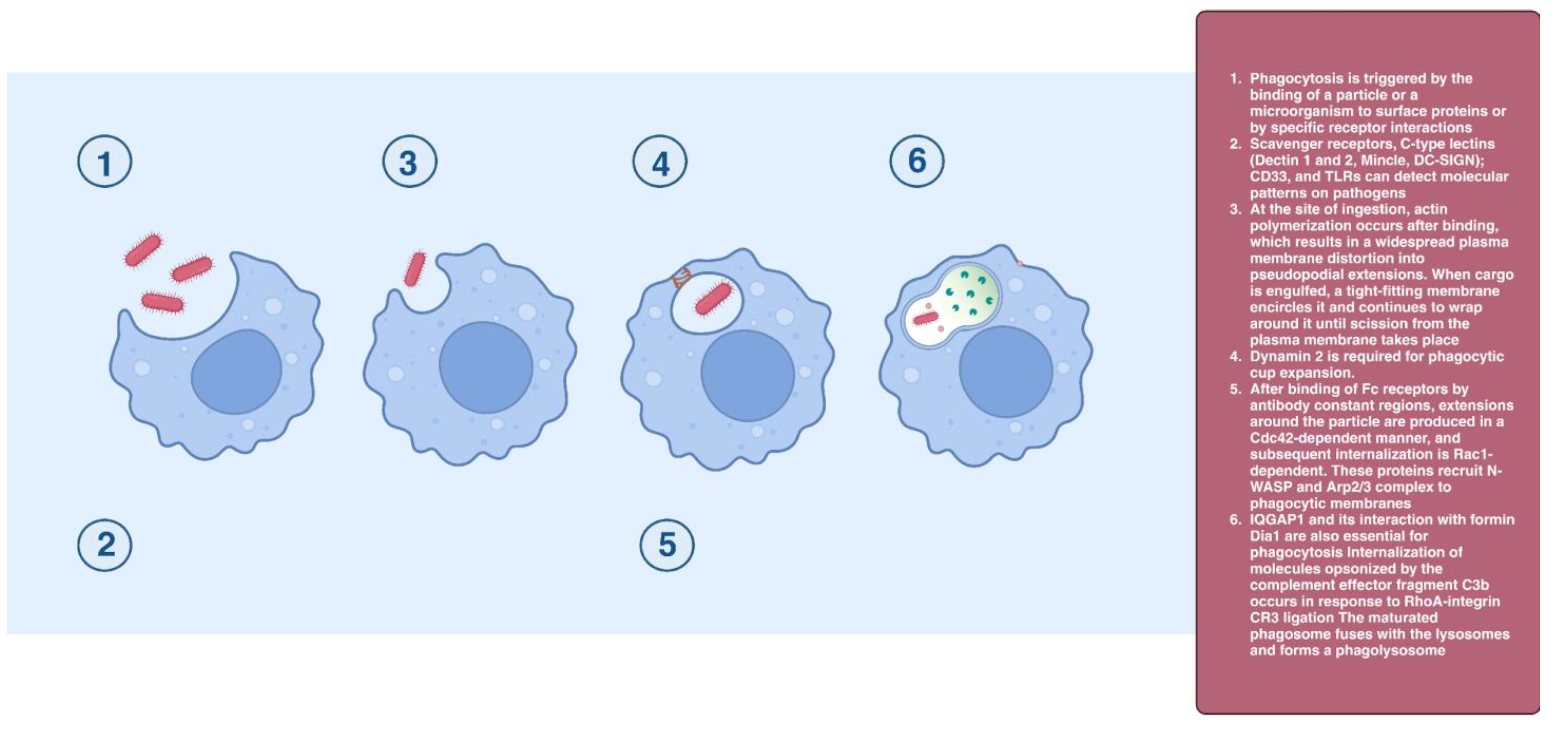
2.7. Phagocytosis
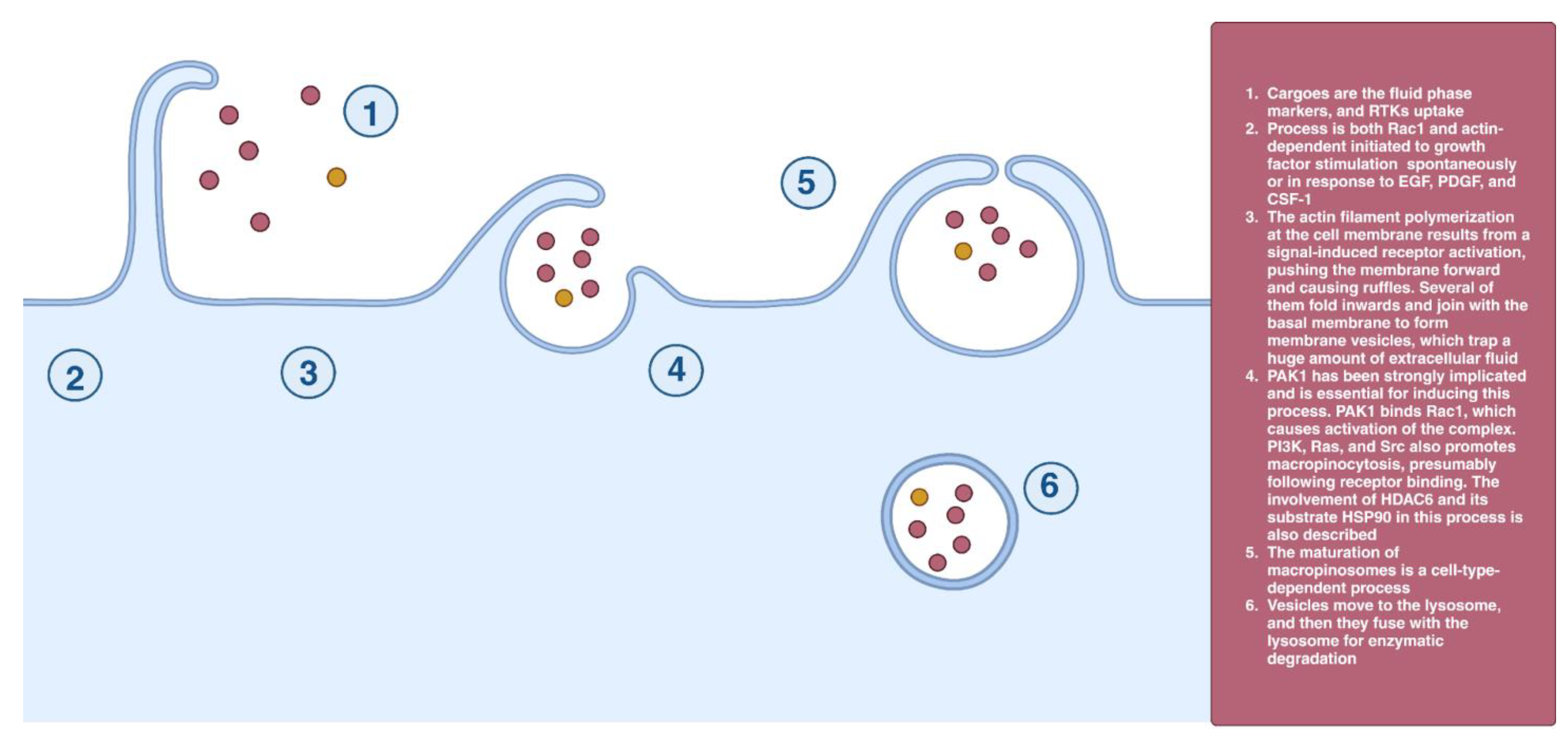
2.8. Macropinocytosis
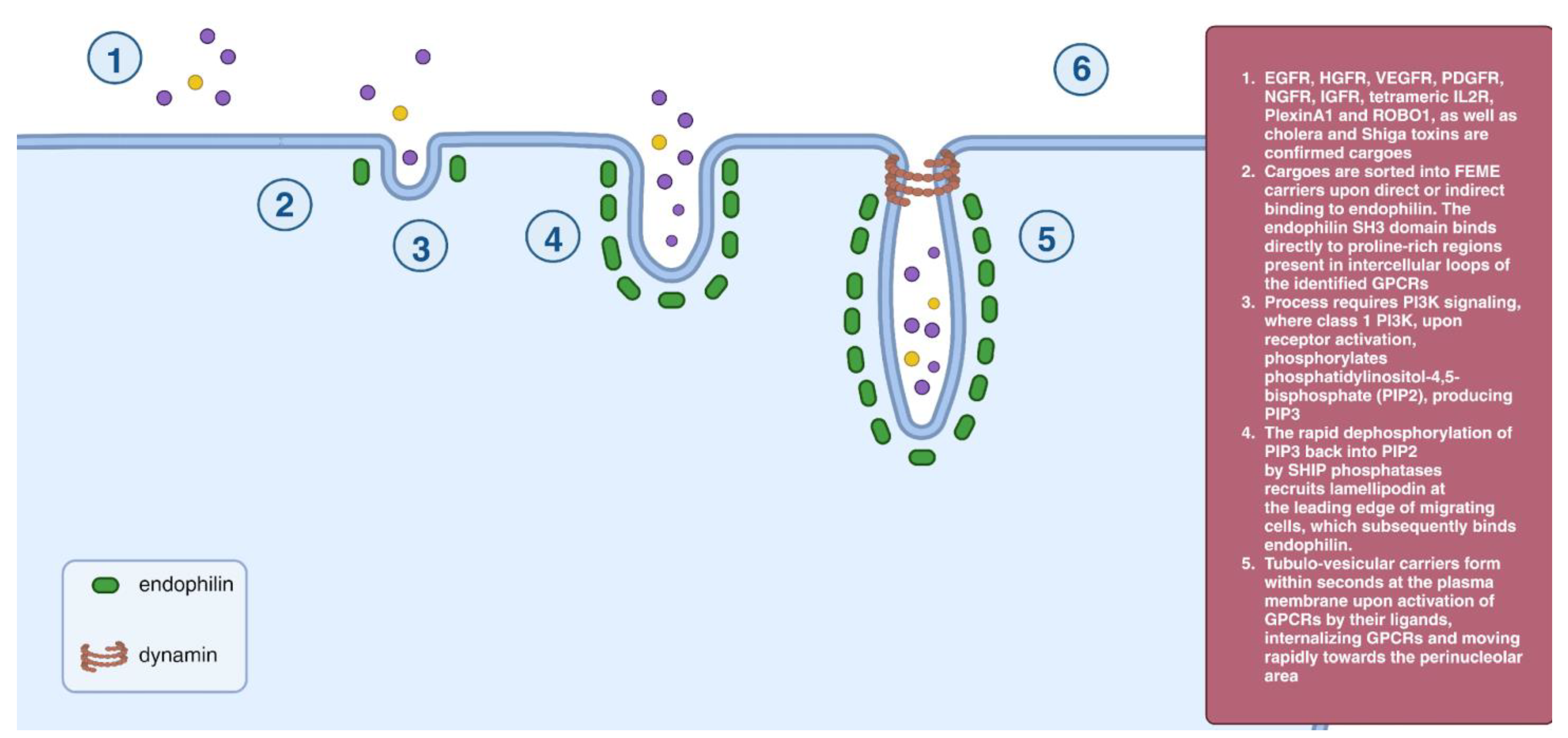
2.9. Fast Endophilin-Mediated Endocytosis (FEME)
3. Endocytosis and Pathologies
3.1. Cardiac Disorders
3.2. Lipid Disorders and Atherosclerosis
3.3. Respiratory Diseases
3.4. Cancer
4. Endocytosis of Viruses
5. Endocytosis of Cell-Penetrating Peptides
6. Endocytosis of Nanoparticles
7. Endocytosis Inhibitors: Mode of Action
7.1. Non-Specific Endocytosis Inhibitors
7.1.1. Potassium Depletion
7.1.2. Hypertonic Medium
7.1.3. Cytosol Acidification
7.1.4. Temperature Decrease
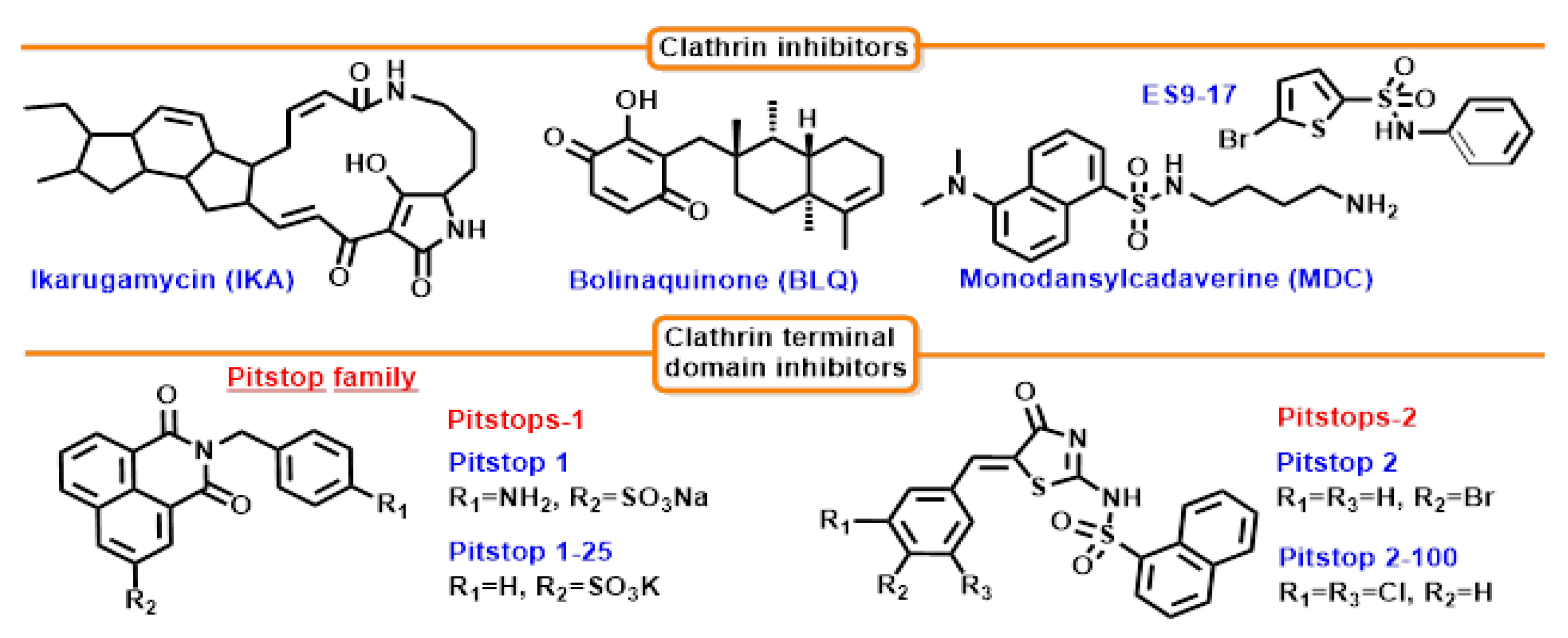
7.2. Clathrin Inhibition
7.2.1. Bolinaquinone (BLQ)
7.2.2. Ikarugamycin (IKA)
7.2.3. ES9-17
7.2.4. Monodansylcadaverine (MDC)
7.3. Clathrin’s Terminal Domain Inhibition
Pitstop Family
7.4. Inhibition of Dynamin
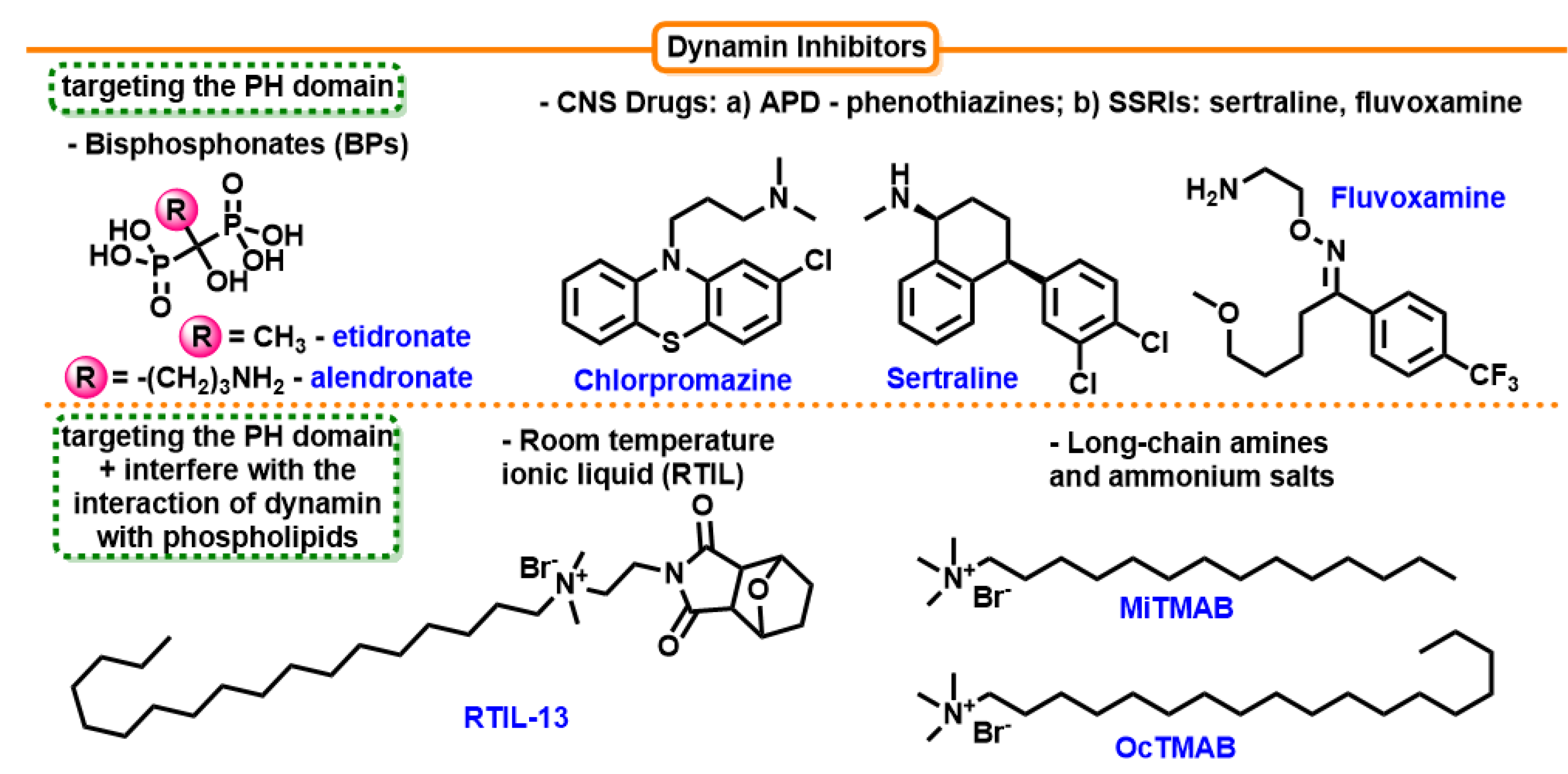
7.4.1. Long-Chain Amines and Long-Chain Ammonium Salts
7.4.2. Antipsychotic Drugs (APD)—Phenothiazines
7.4.3. Selective Serotonin Reuptake Inhibitors (SSRIs)
7.4.4. Room-temperature Ionic Liquid (RTIL)
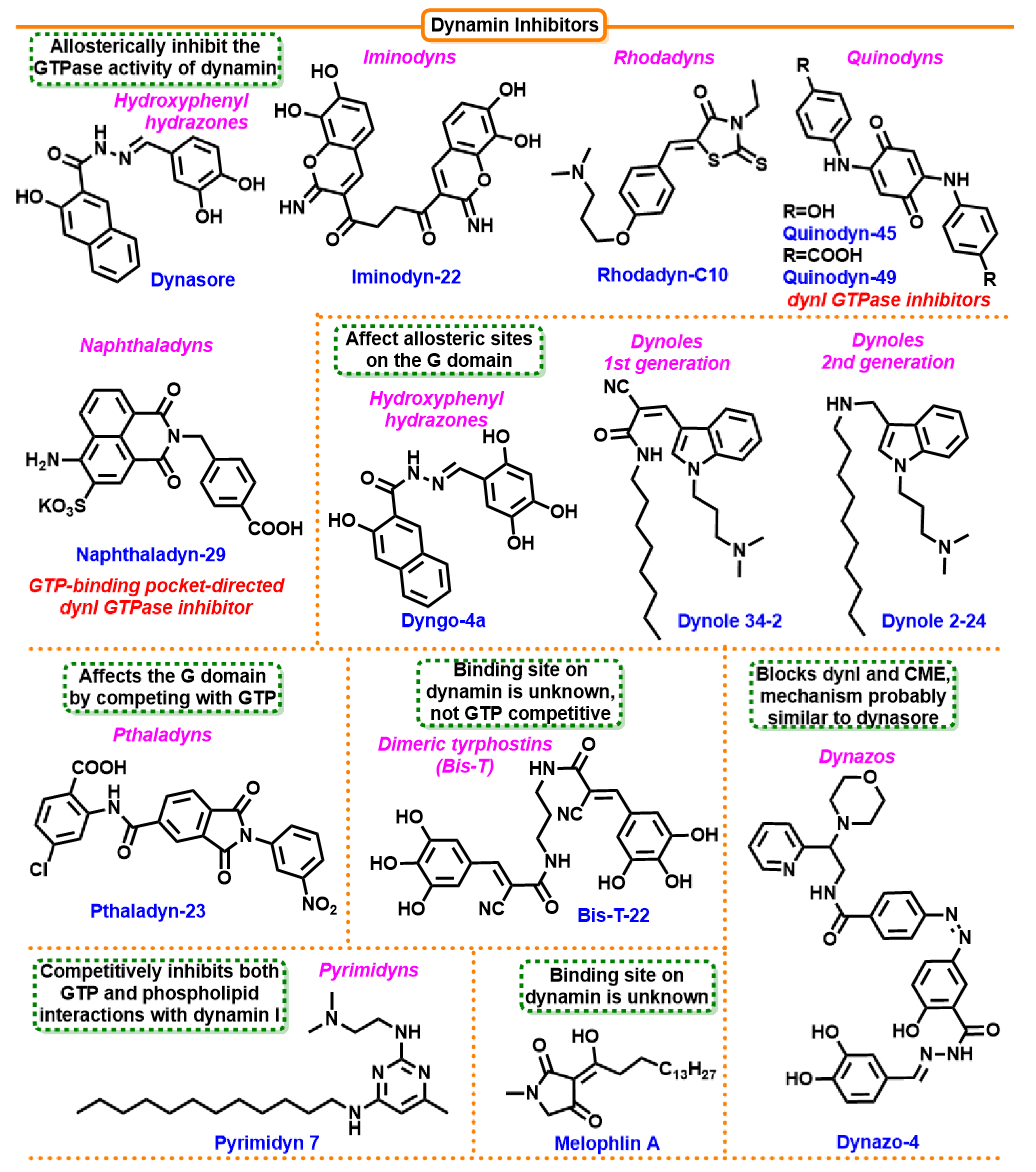
7.4.5. Dimeric Tyrphostins (Bis-T)
7.4.6. Dynasore
7.4.7. Dynoles
7.4.8. Iminodines
7.4.9. Pthaladyns
7.4.10. Rhodanines
7.4.11. Dynoles 2
7.4.12. Pyrimidines
7.4.13. Dyngo
7.4.14. Naphthaladyn Series
7.4.15. Quinones
7.4.16. Dynazos
7.5. Other Binding Sites for Inhibitors
7.5.1. Nocodazole and Paclitaxel
7.5.2. Genistein
7.5.3. Phenylarsine Oxide
8. Off-Target Activity of Endocytosis Inhibitors
9. Clinical Applications
10. Questions and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sorkin, A. Cargo recognition during clathrin-mediated endocytosis: A team effort. Curr. Opin. Cell Biol. 2004, 16, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Bitsikas, V.; Corrêa, I.R.; Nichols, B.J. Clathrin-independent pathways do not contribute significantly to endocytic flux. eLife 2014, 3, e03970. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, M.; Parton, R.G. Clathrin-independent endocytosis: New insights into caveolae and non-caveolar lipid raft carriers. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2005, 1745, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, M.; Fujita, A.; Chadda, R.; Nixon, S.J.; Kurzchalia, T.V.; Sharma, D.K.; Pagano, R.E.; Hancock, J.F.; Mayor, S.; Parton, R.G. Ultrastructural identification of uncoated caveolin-independent early endocytic vehicles. J. Cell Biol. 2005, 168, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.; Helenius, A. Virus Entry: Open Sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef]
- Schmid, E.M.; McMahon, H.T. Integrating molecular and network biology to decode endocytosis. Nature 2007, 448, 883–888. [Google Scholar] [CrossRef]
- Khan, I.; Steeg, P.S. Endocytosis: A pivotal pathway for regulating metastasis. Br. J. Cancer 2021, 124, 66–75. [Google Scholar] [CrossRef]
- Xu, Y.; Buikema, H.; van Gilst, W.H.; Henning, R.H. Caveolae and endothelial dysfunction: Filling the caves in cardiovascular disease. Eur. J. Pharmacol. 2008, 585, 256–260. [Google Scholar] [CrossRef]
- Le Lay, S.; Blouin, C.M.; Hajduch, E.; Dugail, I. Filling up adipocytes with lipids. Lessons from caveolin-1 deficiency. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2009, 1791, 514–518. [Google Scholar] [CrossRef]
- Frank, P.G.; Pavlides, S.; Lisanti, M.P. Caveolae and transcytosis in endothelial cells: Role in atherosclerosis. Cell Tissue Res. 2009, 335, 41–47. [Google Scholar] [CrossRef]
- Gosens, R.; Zaagsma, J.; Meurs, H.; Halayko, A.J. Muscarinic receptor signaling in the pathophysiology of asthma and COPD. Respir. Res. 2006, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Paramasivam, P.; Franke, C.; Stöter, M.; Höijer, A.; Bartesaghi, S.; Sabirsh, A.; Lindfors, L.; Arteta, M.Y.; Dahlén, A.; Bak, A.; et al. Endosomal escape of delivered mRNA from endosomal recycling tubules visualized at the nanoscale. J. Cell Biol. 2022, 221, e202110137. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Effah, C.Y.; Wu, L.; Yu, F.; Wei, J.; Mao, G.; Xiong, Y.; He, L. Endocytosis and intracellular RNAs imaging of nanomaterials-based fluorescence probes. Talanta 2022, 243, 123377. [Google Scholar] [CrossRef] [PubMed]
- Alkafaas, S.S.; Abdallah, A.M.; Ghosh, S.; Loutfy, S.A.; Elkafas, S.S.; Fattah, N.A.; Hessien, M. Insight into the role of clathrin-mediated endocytosis inhibitors in SARS-CoV-2 infection. Rev. Med. Virol. 2023, 33, e2403. [Google Scholar] [CrossRef] [PubMed]
- Matsui, W.; Kirchhausen, T. Stabilization of clathrin coats by the core of the clathrin-associated protein complex AP-2. Biochemistry 1990, 29, 10791–10798. [Google Scholar] [CrossRef]
- Jost, M.; Simpson, F.; Kavran, J.M.; Lemmon, M.A.; Schmid, S.L. Phosphatidylinositol-4,5-bisphosphate is required for endocytic coated vesicle formation. Curr. Biol. 1998, 8, 1399–1404. [Google Scholar] [CrossRef]
- Dawson, J.C.; Legg, J.A.; Machesky, L.M. Bar domain proteins: A role in tubulation, scission and actin assembly in clathrin-mediated endocytosis. Trends Cell Biol. 2006, 16, 493–498. [Google Scholar] [CrossRef]
- Pelkmans, L.; Zerial, M. Kinase-regulated quantal assemblies and kiss-and-run recycling of caveolae. Nature 2005, 436, 128–133. [Google Scholar] [CrossRef]
- Sargiacomo, M.; Scherer, P.E.; Tang, Z.; Kübler, E.; Song, K.S.; Sanders, M.C.; Lisanti, M.P. Oligomeric structure of caveolin: Implications for caveolae membrane organization. Proc. Natl. Acad. Sci. USA 1995, 92, 9407–9411. [Google Scholar] [CrossRef]
- Dietzen, D.J.; Hastings, W.R.; Lublin, D.M. Caveolin Is Palmitoylated on Multiple Cysteine Residues. J. Biol. Chem. 1995, 270, 6838–6842. [Google Scholar] [CrossRef]
- Monier, S.; Dietzen, D.J.; Hastings, W.R.; Lublin, D.M.; Kurzchalia, T.V. Oligomerization of VIP21-caveolin in vitro is stabilized by long chain fatty acylation or cholesterol. FEBS Lett. 1996, 388, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, K.G.; Heuser, J.E.; Donzell, W.C.; Ying, Y.-S.; Glenney, J.R.; Anderson, R.G.W. Caveolin, a protein component of caveolae membrane coats. Cell 1992, 68, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Richter, T.; Floetenmeyer, M.; Ferguson, C.; Galea, J.; Goh, J.; Lindsay, M.R.; Morgan, G.P.; Marsh, B.J.; Parton, R.G. High-Resolution 3D Quantitative Analysis of Caveolar Ultrastructure and Caveola–Cytoskeleton Interactions. Traffic 2008, 9, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Morén, B.; Shah, C.; Howes, M.T.; Schieber, N.L.; McMahon, H.T.; Parton, R.G.; Daumke, O.; Lundmark, R. EHD2 regulates caveolar dynamics via ATP-driven targeting and oligomerization. Mol. Biol. Cell 2012, 23, 1316–1329. [Google Scholar] [CrossRef]
- Matthaeus, C.; Lahmann, I.; Kunz, S.; Jonas, W.; Melo, A.A.; Lehmann, M.; Larsson, E.; Lundmark, R.; Kern, M.; Blüher, M.; et al. EHD2-mediated restriction of caveolar dynamics regulates cellular fatty acid uptake. Proc. Natl. Acad. Sci. USA 2020, 117, 7471–7481. [Google Scholar] [CrossRef]
- Gundu, C.; Arruri, V.K.; Yadav, P.; Navik, U.; Kumar, A.; Amalkar, V.S.; Vikram, A.; Gaddam, R.R. Dynamin-Independent Mechanisms of Endocytosis and Receptor Trafficking. Cells 2022, 11, 2557. [Google Scholar] [CrossRef]
- Kalia, M.; Kumari, S.; Chadda, R.; Hill, M.M.; Parton, R.G.; Mayor, S. Arf6-independent GPI-anchored Protein-enriched Early Endosomal Compartments Fuse with Sorting Endosomes via a Rab5/Phosphatidylinositol-3′-Kinase–dependent Machinery. Mol. Biol. Cell 2006, 17, 3689–3704. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006, 16, 522–529. [Google Scholar] [CrossRef]
- Lundmark, R.; Doherty, G.J.; Howes, M.T.; Cortese, K.; Vallis, Y.; Parton, R.G.; McMahon, H.T. The GTPase-Activating Protein GRAF1 Regulates the CLIC/GEEC Endocytic Pathway. Curr. Biol. 2008, 18, 1802–1808. [Google Scholar] [CrossRef]
- Lamaze, C.; Dujeancourt, A.; Baba, T.; Lo, C.G.; Benmerah, A.; Dautry-Varsat, A. Interleukin 2 Receptors and Detergent-Resistant Membrane Domains Define a Clathrin-Independent Endocytic Pathway. Mol. Cell 2001, 7, 661–671. [Google Scholar] [CrossRef]
- Grassart, A.; Dujeancourt, A.; Lazarow, P.B.; Dautry-Varsat, A.; Sauvonnet, N. Clathrin-independent endocytosis used by the IL-2 receptor is regulated by Rac1, Pak1 and Pak2. EMBO Rep. 2008, 9, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, J.K.; Sedgwick, A.E.; D’Souza-Schorey, C. ARF6-mediated endocytic recycling impacts cell movement, cell division and lipid homeostasis. Semin. Cell Dev. Biol. 2011, 22, 39–47. [Google Scholar] [CrossRef]
- Karim, Z.A.; Choi, W.; Whiteheart, S.W. Primary Platelet Signaling Cascades and Integrin-mediated Signaling Control ADP-ribosylation Factor (Arf) 6-GTP Levels during Platelet Activation and Aggregation. J. Biol. Chem. 2008, 283, 11995–12003. [Google Scholar] [CrossRef]
- Garner, M.J.; Hayward, R.D.; Koronakis, V. The Salmonella pathogenicity island 1 secretion system directs cellular cholesterol redistribution during mammalian cell entry and intracellular trafficking. Cell Microbiol. 2002, 4, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Cain, R.J.; Hayward, R.D.; Koronakis, V. The target cell plasma membrane is a critical interface for Salmonella cell entry effector-host interplay. Mol. Microbiol. 2004, 54, 887–904. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, C.E.; Galán, J.E. Structural mimicry in bacterial virulence. Nature 2001, 412, 701–705. [Google Scholar] [CrossRef]
- Minshall, R.D.; Tiruppathi, C.; Vogel, S.M.; Niles, W.D.; Gilchrist, A.; Hamm, H.E.; Malik, A.B. Endothelial Cell-Surface Gp60 Activates Vesicle Formation and Trafficking via Gi-Coupled Src Kinase Signaling Pathway. J. Cell Biol. 2000, 150, 1057–1070. [Google Scholar] [CrossRef]
- Naslavsky, N.; Weigert, R.; Donaldson, J.G. Convergence of Non-clathrin- and Clathrin-derived Endosomes Involves Arf6 Inactivation and Changes in Phosphoinositides. Mol. Biol. Cell 2003, 14, 417–431. [Google Scholar] [CrossRef]
- Glebov, O.O.; Bright, N.A.; Nichols, B.J. Flotillin-1 defines a clathrin-independent endocytic pathway in mammalian cells. Nat. Cell Biol. 2006, 8, 46–54. [Google Scholar] [CrossRef]
- Payne, C.K.; Jones, S.A.; Chen, C.; Zhuang, X. Internalization and Trafficking of Cell Surface Proteoglycans and Proteoglycan-Binding Ligands. Traffic 2007, 8, 389–401. [Google Scholar] [CrossRef]
- Riento, K.; Frick, M.; Schafer, I.; Nichols, B.J. Endocytosis of flotillin-1 and flotillin-2 is regulated by Fyn kinase. J. Cell Sci. 2009, 122, 912–918. [Google Scholar] [CrossRef] [PubMed]
- John, B.A.; Meister, M.; Banning, A.; Tikkanen, R. Flotillins bind to the dileucine sorting motif of β-site amyloid precursor protein-cleaving enzyme 1 and influence its endosomal sorting. FEBS J. 2014, 281, 2074–2087. [Google Scholar] [CrossRef] [PubMed]
- Marion, S.; Mazzolini, J.; Herit, F.; Bourdoncle, P.; Kambou-Pene, N.; Hailfinger, S.; Sachse, M.; Ruland, J.; Benmerah, A.; Echard, A.; et al. The NF-κB Signaling Protein Bcl10 Regulates Actin Dynamics by Controlling AP1 and OCRL-Bearing Vesicles. Dev. Cell 2012, 23, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Masters, T.A.; Pontes, B.; Viasnoff, V.; Li, Y.; Gauthier, N.C. Plasma membrane tension orchestrates membrane trafficking, cytoskeletal remodeling, and biochemical signaling during phagocytosis. Proc. Natl. Acad. Sci. USA 2013, 110, 11875–11880. [Google Scholar] [CrossRef] [PubMed]
- Massol, P.; Montcourrier, P.; Guillemot, J.-C.; Chavrier, P. Fc receptor-mediated phagocytosis requires CDC42 and Rac1. EMBO J. 1998, 17, 6219–6229. [Google Scholar] [CrossRef] [PubMed]
- Racoosin, E.L.; Swanson, J.A. Macropinosome maturation and fusion with tubular lysosomes in macrophages. J. Cell Biol. 1993, 121, 1011–1020. [Google Scholar] [CrossRef]
- Amyere, M.; Payrastre, B.; Krause, U.; Van Der Smissen, P.; Veithen, A.; Courtoy, P.J. Constitutive Macropinocytosis in Oncogene-transformed Fibroblasts Depends on Sequential Permanent Activation of Phosphoinositide 3-Kinase and Phospholipase C. Mol. Biol. Cell 2000, 11, 3453–3467. [Google Scholar] [CrossRef]
- Hewlett, L.; Prescott, A.; Watts, C. The coated pit and macropinocytic pathways serve distinct endosome populations. J. Cell Biol. 1994, 124, 689–703. [Google Scholar] [CrossRef]
- Dharmawardhane, S.; Schürmann, A.; Sells, M.A.; Chernoff, J.; Schmid, S.L.; Bokoch, G.M. Regulation of Macropinocytosis by p21-activated Kinase-1. Mol. Biol. Cell 2000, 11, 3341–3352. [Google Scholar] [CrossRef]
- Knaus, U.G.; Wang, Y.; Reilly, A.M.; Warnock, D.; Jackson, J.H. Structural Requirements for PAK Activation by Rac GTPases. J. Biol. Chem. 1998, 273, 21512–21518. [Google Scholar] [CrossRef]
- Gao, Y.; Hubbert, C.C.; Lu, J.; Lee, Y.-S.; Lee, J.-Y.; Yao, T.-P. Histone Deacetylase 6 Regulates Growth Factor-Induced Actin Remodeling and Endocytosis. Mol. Cell Biol. 2007, 27, 8637–8647. [Google Scholar] [CrossRef]
- Grimmer, S.; van Deurs, B.; Sandvig, K. Membrane ruffling and macropinocytosis in A431 cells require cholesterol. J. Cell Sci. 2002, 115, 2953–2962. [Google Scholar] [CrossRef]
- Casamento, A.; Boucrot, E. Molecular mechanism of Fast Endophilin-Mediated Endocytosis. Biochem. J. 2020, 477, 2327–2345. [Google Scholar] [CrossRef] [PubMed]
- Boucrot, E.; Ferreira, A.P.; Almeida-Souza, L.; Debard, S.; Vallis, Y.; Howard, G.; Bertot, L.; Sauvonnet, N.; McMahon, H.T. Endophilin marks and controls a clathrin-independent endocytic pathway. Nature 2015, 517, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Renard, H.F.; Simunovic, M.; Lemière, J.; Boucrot, E.; Garcia-Castillo, M.D.; Arumugam, S.; Chambon, V.; Lamaze, C.; Wunder, C.; Kenworthy, A.K.; et al. Endophilin-A2 functions in membrane scission in clathrin-independent endocytosis. Nature 2015, 517, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Le Lay, S.; Kurzchalia, T.V. Getting rid of caveolins: Phenotypes of caveolin-deficient animals. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2005, 1746, 322–333. [Google Scholar] [CrossRef]
- Okamoto, T.; Schlegel, A.; Scherer, P.E.; Lisanti, M.P. Caveolins, a Family of Scaffolding Proteins for Organizing ‘Preassembled Signaling Complexes’ at the Plasma Membrane. J. Biol. Chem. 1998, 273, 5419–5422. [Google Scholar] [CrossRef] [PubMed]
- Roth, D.M.; Patel, H.H. Role of Caveolae in Cardiac Protection. Pediatr. Cardiol. 2011, 32, 329–333. [Google Scholar] [CrossRef]
- Albrecht, E.W.; Stegeman, C.A.; Heeringa, P.; Henning, R.H.; van Goor, H. Protective role of endothelial nitric oxide synthase. J. Pathol. 2003, 199, 8–17. [Google Scholar] [CrossRef]
- Minshall, R.D.; Sessa, W.C.; Stan, R.V.; Anderson, R.G.W.; Malik, A.B. Caveolin regulation of endothelial function. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 285, L1179–L1183. [Google Scholar] [CrossRef]
- Sessa, W.C. eNOS at a glance. J. Cell Sci. 2004, 117, 2427–2429. [Google Scholar] [CrossRef] [PubMed]
- Sessa, W.C. Regulation of endothelial derived nitric oxide in health and disease. Mem. Inst. Oswaldo Cruz. 2005, 100 (Suppl. S1), 15–18. [Google Scholar] [CrossRef] [PubMed]
- Gratton, J.P.; Lin, M.I.; Yu, J.; Weiss, E.D.; Jiang, Z.L.; Fairchild, T.A.; Iwakiri, Y.; Groszmann, R.; Claffey, K.P.; Cheng, Y.C.; et al. Selective inhibition of tumor microvascular permeability by cavtratin blocks tumor progression in mice. Cancer Cell 2003, 4, 31–39. [Google Scholar] [CrossRef]
- Young, L.H.; Ikeda, Y.; Lefer, A.M. Caveolin-1 peptide exerts cardioprotective effects in myocardial ischemia-reperfusion via nitric oxide mechanism. Am. J. Physiol.-Heart Circ. Physiol. 2001, 280, H2489–H2495. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Kuroda, R.; Sakoda, T.; Katayama, M.; Inoue, N.; Suematsu, M.; Kawashima, S.; Yokoyama, M. Inhibition of Endothelial Nitric Oxide Synthase Activity by Protein Kinase C. Hypertension 1995, 25, 180–185. [Google Scholar] [CrossRef]
- Zulueta, J.J.; Yu, F.S.; Hertig, I.A.; Thannickal, V.J.; Hassoun, P.M. Release of hydrogen peroxide in response to hypoxia-reoxygenation: Role of an NAD(P)H oxidase-like enzyme in endothelial cell plasma membrane. Am. J. Respir. Cell Mol. Biol. 1995, 12, 41–49. [Google Scholar] [CrossRef]
- Cohen, A.W.; Park, D.S.; Woodman, S.E.; Williams, T.M.; Chandra, M.; Shirani, J.; Pereira de Souza, A.; Kitsis, R.N.; Russell, R.G.; Weiss, L.M.; et al. Caveolin-1 null mice develop cardiac hypertrophy with hyperactivation of p42/44 MAP kinase in cardiac fibroblasts. Am. J. Physiol.-Cell Physiol. 2003, 284, C457–C474. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Liu, Y.; Stan, R.V.; Fan, L.; Gu, Y.; Dalton, N.; Chu, P.H.; Peterson, K.; Ross, J., Jr.; Chien, K.R. Defects in caveolin-1 cause dilated cardiomyopathy and pulmonary hypertension in knockout mice. Proc. Natl. Acad. Sci. USA 2002, 99, 11375–11380. [Google Scholar] [CrossRef]
- Jones, A.; Hradek, G.; Hornick, C.; Renaud, G.; Windler, E.; Havel, R. Uptake and processing of remnants of chylomicrons and very low density lipoproteins by rat liver. J. Lipid Res. 1984, 25, 1151–1158. [Google Scholar] [CrossRef]
- Lambert, G.; Sjouke, B.; Choque, B.; Kastelein, J.J.P.; Hovingh, G.K. The PCSK9 decade. J. Lipid Res. 2012, 53, 2515–2524. [Google Scholar] [CrossRef]
- Zelcer, N.; Hong, C.; Boyadjian, R.; Tontonoz, P. LXR Regulates Cholesterol Uptake Through Idol-Dependent Ubiquitination of the LDL Receptor. Science (1979) 2009, 325, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Bartuzi, P.; Billadeau, D.D.; Favier, R.; Rong, S.; Dekker, D.; Fedoseienko, A.; Fieten, H.; Wijers, M.; Levels, J.H.; Huijkman, N.; et al. CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL. Nat. Commun. 2016, 7, 10961. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Lacolley, P.; Regnault, V.; Nicoletti, A.; Li, Z.; Michel, J.-B. The vascular smooth muscle cell in arterial pathology: A cell that can take on multiple roles. Cardiovasc. Res. 2012, 95, 194–204. [Google Scholar] [CrossRef]
- Khalil, M.F.; Wagner, W.D.; Goldberg, I.J. Molecular Interactions Leading to Lipoprotein Retention and the Initiation of Atherosclerosis. Arter. Thromb. Vasc. Biol. 2004, 24, 2211–2218. [Google Scholar] [CrossRef]
- Dubland, J.A.; Francis, G.A. So Much Cholesterol. Curr. Opin. Lipidol. 2016, 27, 155–161. [Google Scholar] [CrossRef]
- Allahverdian, S.; Chehroudi, A.C.; McManus, B.M.; Abraham, T.; Francis, G.A. Contribution of Intimal Smooth Muscle Cells to Cholesterol Accumulation and Macrophage-Like Cells in Human Atherosclerosis. Circulation 2014, 129, 1551–1559. [Google Scholar] [CrossRef]
- Feil, S.; Fehrenbacher, B.; Lukowski, R.; Essmann, F.; Schulze-Osthoff, K.; Schaller, M.; Feil, R. Transdifferentiation of Vascular Smooth Muscle Cells to Macrophage-Like Cells During Atherogenesis. Circ. Res. 2014, 115, 662–667. [Google Scholar] [CrossRef]
- Lao, K.H.; Zeng, L.; Xu, Q. Endothelial and smooth muscle cell transformation in atherosclerosis. Curr. Opin. Lipidol. 2015, 26, 449–456. [Google Scholar] [CrossRef]
- Andreeva, E.R.; Pugach, I.M.; Orekhov, A.N. Subendothelial smooth muscle cells of human aorta express macrophage antigen in situ and in vitro. Atherosclerosis 1997, 135, 19–27. [Google Scholar] [CrossRef]
- Rong, J.X.; Shapiro, M.; Trogan, E.; Fisher, E.A. Transdifferentiation of mouse aortic smooth muscle cells to a macrophage-like state after cholesterol loading. Proc. Natl. Acad. Sci. USA 2003, 100, 13531–13536. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, F.; Volonté, D.; Liu, J.; Capozza, F.; Frank, P.G.; Zhu, L.; Pestell, R.G.; Lisanti, M.P. Caveolin-1 Expression Negatively Regulates Cell Cycle Progression by Inducing G0 /G1 Arrest via a p53/p21 WAF1/Cip1 -dependent Mechanism. Mol. Biol. Cell 2001, 12, 2229–2244. [Google Scholar] [CrossRef] [PubMed]
- Nemenoff, R.A.; Horita, H.; Ostriker, A.C.; Furgeson, S.B.; Simpson, P.A.; VanPutten, V.; Crossno, J.; Offermanns, S.; Weiser-Evans, M.C. SDF-1α Induction in Mature Smooth Muscle Cells by Inactivation of PTEN Is a Critical Mediator of Exacerbated Injury-Induced Neointima Formation. Arter. Thromb. Vasc. Biol. 2011, 31, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Bauriedel, G. Role of smooth muscle cell death in advanced coronary primary lesions: Implications for plaque instability. Cardiovasc. Res. 1999, 41, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Frontini, M.J.; O’Neil, C.; Sawyez, C.; Chan, B.M.C.; Huff, M.W.; Pickering, J.G. Lipid Incorporation Inhibits Src-Dependent Assembly of Fibronectin and Type I Collagen by Vascular Smooth Muscle Cells. Circ. Res. 2009, 104, 832–841. [Google Scholar] [CrossRef]
- Krasteva, G.; Pfeil, U.; Drab, M.; Kummer, W.; König, P. Caveolin-1 and -2 in airway epithelium: Expression and in situ association as detected by FRET-CLSM. Respir. Res. 2006, 7, 108. [Google Scholar] [CrossRef]
- Drab, M.; Verkade, P.; Elger, M.; Kasper, M.; Lohn, M.; Lauterbach, B.; Menne, J.; Lindschau, C.; Mende, F.; Luft, F.C.; et al. Loss of Caveolae, Vascular Dysfunction, and Pulmonary Defects in Caveolin-1 Gene-Disrupted Mice. Science (1979) 2001, 293, 2449–2452. [Google Scholar] [CrossRef]
- Razani, B.; Engelman, J.A.; Wang, X.B.; Schubert, W.; Zhang, X.L.; Marks, C.B.; Macaluso, F.; Russell, R.G.; Li, M.; Pestell, R.G.; et al. Caveolin-1 Null Mice Are Viable but Show Evidence of Hyperproliferative and Vascular Abnormalities. J. Biol. Chem. 2001, 276, 38121–38138. [Google Scholar] [CrossRef]
- Razani, B.; Wang, X.B.; Engelman, J.A.; Battista, M.; Lagaud, G.; Zhang, X.L.; Kneitz, B.; Hou, H., Jr.; Christ, G.J.; Edelmann, W.; et al. Caveolin-2-Deficient Mice Show Evidence of Severe Pulmonary Dysfunction without Disruption of Caveolae. Mol. Cell Biol. 2002, 22, 2329–2344. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhang, Y.; Kim, H.P.; Zhou, Z.; Feghali-Bostwick, C.A.; Liu, F.; Ifedigbo, E.; Xu, X.; Oury, T.D.; Kaminski, N.; et al. Caveolin-1: A critical regulator of lung fibrosis in idiopathic pulmonary fibrosis. J. Exp. Med. 2006, 203, 2895–2906. [Google Scholar] [CrossRef]
- Racine, C.; Bélanger, M.; Hirabayashi, H.; Boucher, M.; Chakir, J.; Couet, J. Reduction of Caveolin 1 Gene Expression in Lung Carcinoma Cell Lines. Biochem. Biophys. Res. Commun. 1999, 255, 580–586. [Google Scholar] [CrossRef]
- Achcar, R.O.; Demura, Y.; Rai, P.R.; Taraseviciene-Stewart, L.; Kasper, M.; Voelkel, N.F.; Cool, C.D. Loss of Caveolin and Heme Oxygenase Expression in Severe Pulmonary Hypertension. Chest 2006, 129, 696–705. [Google Scholar] [CrossRef]
- Opazo, A.M.; Seow, C.Y.; Paré, P.D. Peripheral Airway Smooth Muscle Mechanics in Obstructive Airways Disease. Am. J. Respir. Crit. Care Med. 2000, 161, 910–917. [Google Scholar] [CrossRef]
- Bai, T.R. Abnormalities in Airway Smooth Muscle in Fatal Asthma. Am. Rev. Respir. Dis. 1990, 141, 552–557. [Google Scholar] [CrossRef]
- Bramley, A.; Thomson, R.; Roberts, C.; Schellenberg, R. Hypothesis: Excessive bronchoconstriction in asthma is due to decreased airway elastance. Eur. Respir. J. 1994, 7, 337–341. [Google Scholar] [CrossRef]
- de Jongste, J.C.; Mons, H.; Bonta, I.L.; Kerrebijn, K.F. In vitro responses of airways from an asthmatic patient. Eur. J. Respir. Dis. 1987, 71, 23–29. [Google Scholar]
- Gosens, R.; Stelmack, G.L.; Dueck, G.; McNeill, K.D.; Yamasaki, A.; Gerthoffer, W.T.; Unruh, H.; Gounni, A.S.; Zaagsma, J.; Halayko, A.J. Role of caveolin-1 in p42/p44 MAP kinase activation and proliferation of human airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2006, 291, L523–L534. [Google Scholar] [CrossRef]
- Thyberg, J.; Roy, J.; Tran, P.K.; Blomgren, K.; Dumitrescu, A.; Hedin, U. Expression of caveolae on the surface of rat arterial smooth muscle cells is dependent on the phenotypic state of the cells. Lab. Investig. 1997, 77, 93–101. [Google Scholar]
- Gabella, G. Quantitative morphological study of smooth muscle cells of the guinea-pig taenia coli. Cell Tissue Res. 1976, 170, 161–186. [Google Scholar] [CrossRef]
- Galbiati, F. Targeted downregulation of caveolin-1 is sufficient to drive cell transformation and hyperactivate the p42/44 MAP kinase cascade. EMBO J. 1998, 17, 6633–6648. [Google Scholar] [CrossRef]
- Gosens, R.; Dueck, G.; Gerthoffer, W.T.; Unruh, H.; Zaagsma, J.; Meurs, H.; Halayko, A.J. p42/p44 MAP kinase activation is localized to caveolae-free membrane domains in airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2007, 292, L1163–L1172. [Google Scholar] [CrossRef]
- Peterson, T.E.; Guicciardi, M.E.; Gulati, R.; Kleppe, L.S.; Mueske, C.S.; Mookadam, M.; Sowa, G.; Gores, G.J.; Sessa, W.C.; Simari, R.D. Caveolin-1 Can Regulate Vascular Smooth Muscle Cell Fate by Switching Platelet-Derived Growth Factor Signaling from a Proliferative to an Apoptotic Pathway. Arter. Thromb. Vasc. Biol. 2003, 23, 1521–1527. [Google Scholar] [CrossRef]
- Volonte, D.; Zhang, K.; Lisanti, M.P.; Galbiati, F. Expression of Caveolin-1 Induces Premature Cellular Senescence in Primary Cultures of Murine Fibroblasts. Mol. Biol. Cell 2002, 13, 2502–2517. [Google Scholar] [CrossRef]
- Banushi, B.; Joseph, S.R.; Lum, B.; Lee, J.J.; Simpson, F. Endocytosis in cancer and cancer therapy. Nat. Rev. Cancer 2023, 23, 450–473. [Google Scholar] [CrossRef]
- Floyd, S.; De Camilli, P. Endocytosis proteins and cancer: A potential link? Trends Cell Biol. 1998, 8, 299–301. [Google Scholar] [CrossRef]
- Di Fiore, P.P.; Polo, S.; Hofmann, K. When ubiquitin meets ubiquitin receptors: A signalling connection. Nat. Rev. Mol. Cell Biol. 2003, 4, 491–497. [Google Scholar] [CrossRef]
- Pickart, C.M. Ubiquitin in chains. Trends Biochem. Sci. 2000, 25, 544–548. [Google Scholar] [CrossRef]
- Haglund, K.; Sigismund, S.; Polo, S.; Szymkiewicz, I.; Di Fiore, P.P.; Dikic, I. Multiple monoubiquitination of RTKs is sufficient for their endocytosis and degradation. Nat. Cell Biol. 2003, 5, 461–466. [Google Scholar] [CrossRef]
- Dikic, I.; Szymkiewicz, I.; Soubeyran, P. Cbl signaling networks in the regulation of cell function. Cell Mol. Life Sci. 2003, 60, 1805–1827. [Google Scholar] [CrossRef]
- Tice, D.A.; Biscardi, J.S.; Nickles, A.L.; Parsons, S.J. Mechanism of biological synergy between cellular Src and epidermal growth factor receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 1415–1420. [Google Scholar] [CrossRef]
- Summy, J.M.; Gallick, G.E. Src family kinases in tumor progression and metastasis. Cancer Metastasis Rev. 2003, 22, 337–358. [Google Scholar] [CrossRef]
- Chew, H.Y.; De Lima, P.O.; Gonzalez Cruz, J.L.; Banushi, B.; Echejoh, G.; Hu, L.; Joseph, S.R.; Lum, B.; Rae, J.; O'Donnell, J.S.; et al. Endocytosis Inhibition in Humans to Improve Responses to ADCC-Mediating Antibodies. Cell 2020, 180, 895–914.e27. [Google Scholar] [CrossRef]
- Widlansky, M.E.; Gokce, N.; Keaney, J.F.; Vita, J.A. The clinical implications of endothelial dysfunction. J. Am. Coll. Cardiol. 2003, 42, 1149–1160. [Google Scholar] [CrossRef]
- Chen, W.J.; Goldstein, J.L.; Brown, M.S. NPXY, a sequence often found in cytoplasmic tails, is required for coated pit-mediated internalization of the low density lipoprotein receptor. J. Biol. Chem. 1990, 265, 3116–3123. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Popescu, L.M. Caveolar nanospaces in smooth muscle cells. J. Cell Mol. Med. 2006, 10, 519–528. [Google Scholar] [CrossRef]
- Kuo, K.-H.; Herrera, A.M.; Seow, C.Y. Ultrastructure of airway smooth muscle. Respir. Physiol. Neurobiol. 2003, 137, 197–208. [Google Scholar] [CrossRef]
- Gratton, J.-P.; Fontana, J.; O’Connor, D.S.; Garcı, G.; McCabe, T.J.; Sessa, W.C. Reconstitution of an Endothelial Nitric-oxide Synthase (eNOS), hsp90, and Caveolin-1 Complex in Vitro. J. Biol. Chem. 2000, 275, 22268–22272. [Google Scholar] [CrossRef]
- Ju, H.; Zou, R.; Venema, V.J.; Venema, R.C. Direct Interaction of Endothelial Nitric-oxide Synthase and Caveolin-1 Inhibits Synthase Activity. J. Biol. Chem. 1997, 272, 18522–18525. [Google Scholar] [CrossRef]
- Lozano, E.; Betson, M.; Braga, V.M.M. Tumor progression: Small GTPases and loss of cell-cell adhesion. BioEssays 2003, 25, 452–463. [Google Scholar] [CrossRef]
- Ugolini, S.; Mondor, I.; Sattentau, Q.J. HIV-1 attachment: Another look. Trends Microbiol. 1999, 7, 144–149. [Google Scholar] [CrossRef]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. CHEMOKINE RECEPTORS AS HIV-1 CORECEPTORS: Roles in Viral Entry, Tropism, and Disease. Annu. Rev. Immunol. 1999, 17, 657–700. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Berson, J.F.; Cook, D.G.; Doms, R.W. Characterization of human immunodeficiency virus type 1 gp120 binding to liposomes containing galactosylceramide. J. Virol. 1994, 68, 5890–5898. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.; Helenius, A. Virus Entry into Animal Cells. Adv. Virus Res. 1989, 36, 107–151. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M.; Boll, W.; Van Oijen, A.; Hariharan, R.; Chandran, K.; Nibert, M.L.; Kirchhausen, T. Endocytosis by Random Initiation and Stabilization of Clathrin-Coated Pits. Cell 2004, 118, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Hessien, M.; Donia, T.; Tabll, A.A.; Adly, E.; Abdelhafez, T.H.; Attia, A.; Alkafaas, S.S.; Kuna, L.; Glasnovic, M.; Cosic, V.; et al. Mechanistic-Based Classification of Endocytosis-Related Inhibitors: Does It Aid in Assigning Drugs against SARS-CoV-2? Viruses 2023, 15, 1040. [Google Scholar] [CrossRef]
- Chandran, K.; Sullivan, N.J.; Felbor, U.; Whelan, S.P.; Cunningham, J.M. Endosomal Proteolysis of the Ebola Virus Glycoprotein Is Necessary for Infection. Science (1979) 2005, 308, 1643–1645. [Google Scholar] [CrossRef]
- Ebert, D.H.; Deussing, J.; Peters, C.; Dermody, T.S. Cathepsin L and Cathepsin B Mediate Reovirus Disassembly in Murine Fibroblast Cells. J. Biol. Chem. 2002, 277, 24609–24617. [Google Scholar] [CrossRef]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef]
- Stehle, T.; Gamblin, S.J.; Yan, Y.; Harrison, S.C. The structure of simian virus 40 refined at 3.1 å resolution. Structure 1996, 4, 165–182. [Google Scholar] [CrossRef]
- Stehle, T. High-resolution structure of a polyomavirus VP1-oligosaccharide complex: Implications for assembly and receptor binding. EMBO J. 1997, 16, 5139–5148. [Google Scholar] [CrossRef]
- Anderson, H.A.; Chen, Y.; Norkin, L.C. Bound simian virus 40 translocates to caveolin-enriched membrane domains, and its entry is inhibited by drugs that selectively disrupt caveolae. Mol. Biol. Cell 1996, 7, 1825–1834. [Google Scholar] [CrossRef]
- Pelkmans, L.; Kartenbeck, J.; Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nat. Cell Biol. 2001, 3, 473–483. [Google Scholar] [CrossRef]
- Tsai, B. Gangliosides are receptors for murine polyoma virus and SV40. EMBO J. 2003, 22, 4346–4355. [Google Scholar] [CrossRef]
- Stang, E.; Kartenbeck, J.; Parton, R.G. Major histocompatibility complex class I molecules mediate association of SV40 with caveolae. Mol. Biol. Cell 1997, 8, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Pelkmans, L.; Pu, D.; Helenius, A. Local Actin Polymerization and Dynamin Recruitment in SV40-Induced Internalization of Caveolae. Science (1979) 2002, 296, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Pelkmans, L.; Fava, E.; Grabner, H.; Hannus, M.; Habermann, B.; Krausz, E.; Zerial, M. Genome-wide analysis of human kinases in clathrin- and caveolae/raft-mediated endocytosis. Nature 2005, 436, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Damke, H.; Baba, T.; van der Bliek, A.M.; Schmid, S.L. Clathrin-independent pinocytosis is induced in cells overexpressing a temperature-sensitive mutant of dynamin. J. Cell Biol. 1995, 131, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ruseska, I.; Zimmer, A. Internalization mechanisms of cell-penetrating peptides. Beilstein J. Nanotechnol. 2020, 11, 101–123. [Google Scholar] [CrossRef]
- Patel, K.P.; Coyne, C.B.; Bergelson, J.M. Dynamin- and Lipid Raft-Dependent Entry of Decay-Accelerating Factor (DAF)-Binding and Non-DAF-Binding Coxsackieviruses into Nonpolarized Cells. J. Virol. 2009, 83, 11064–11077. [Google Scholar] [CrossRef]
- Pae, J.; Säälik, P.; Liivamägi, L.; Lubenets, D.; Arukuusk, P.; Langel, Ü.; Pooga, M. Translocation of cell-penetrating peptides across the plasma membrane is controlled by cholesterol and microenvironment created by membranous proteins. J. Control. Release 2014, 192, 103–113. [Google Scholar] [CrossRef]
- Fittipaldi, A.; Ferrari, A.; Zoppé, M.; Arcangeli, C.; Pellegrini, V.; Beltram, F.; Giacca, M. Cell Membrane Lipid Rafts Mediate Caveolar Endocytosis of HIV-1 Tat Fusion Proteins. J. Biol. Chem. 2003, 278, 34141–34149. [Google Scholar] [CrossRef]
- Pujals, S.; Giralt, E. Proline-rich, amphipathic cell-penetrating peptides. Adv. Drug Deliv. Rev. 2008, 60, 473–484. [Google Scholar] [CrossRef]
- Veiman, K.L.; Mäger, I.; Ezzat, K.; Margus, H.; Lehto, T.; Langel, K.; Kurrikoff, K.; Arukuusk, P.; Suhorutšenko, J.; Padari, K.; et al. PepFect14 Peptide Vector for Efficient Gene Delivery in Cell Cultures. Mol. Pharm. 2013, 10, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Zheng, W.; Li, A.; Mu, Y.; Shi, M.; Li, T.; Zou, H.; Shao, H.; Qin, A.; Ye, J. A novel CAV derived cell-penetrating peptide efficiently delivers exogenous molecules through caveolae-mediated endocytosis. Vet. Res. 2018, 49, 16. [Google Scholar] [CrossRef] [PubMed]
- Nakase, I.; Niwa, M.; Takeuchi, T.; Sonomura, K.; Kawabata, N.; Koike, Y.; Takehashi, M.; Tanaka, S.; Ueda, K.; Simpson, J.C.; et al. Cellular Uptake of Arginine-Rich Peptides: Roles for Macropinocytosis and Actin Rearrangement. Mol. Ther. 2004, 10, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.M.; Wadia, J.S.; Dowdy, S.F. Cationic TAT peptide transduction domain enters cells by macropinocytosis. J. Control. Release 2005, 102, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Wadia, J.S.; Stan, R.V.; Dowdy, S.F. Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosis. Nat. Med. 2004, 10, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Nakase, I.; Hirose, H.; Tanaka, G.; Tadokoro, A.; Kobayashi, S.; Takeuchi, T.; Futaki, S. Cell-surface Accumulation of Flock House Virus-derived Peptide Leads to Efficient Internalization via Macropinocytosis. Mol. Ther. 2009, 17, 1868–1876. [Google Scholar] [CrossRef]
- Tanaka, G.; Nakase, I.; Fukuda, Y.; Masuda, R.; Oishi, S.; Shimura, K.; Kawaguchi, Y.; Takatani-Nakase, T.; Langel, U.; Gräslund, A.; et al. CXCR4 Stimulates Macropinocytosis: Implications for Cellular Uptake of Arginine-Rich Cell-Penetrating Peptides and HIV. Chem. Biol. 2012, 19, 1437–1446. [Google Scholar] [CrossRef]
- Marino, J.; Wigdahl, B.; Nonnemacher, M.R. Extracellular HIV-1 Tat Mediates Increased Glutamate in the CNS Leading to Onset of Senescence and Progression of HAND. Front. Aging Neurosci. 2020, 12, 168. [Google Scholar] [CrossRef]
- Dickens, A.M.; Yoo, S.W.; Chin, A.C.; Xu, J.; Johnson, T.P.; Trout, A.L.; Hauser, K.F.; Haughey, N.J. Chronic low-level expression of HIV-1 Tat promotes a neurodegenerative phenotype with aging. Sci. Rep. 2017, 7, 7748. [Google Scholar] [CrossRef]
- Roszczenko, P.; Szewczyk, O.K.; Czarnomysy, R.; Bielawski, K.; Bielawska, A. Biosynthesized Gold, Silver, Palladium, Platinum, Copper, and Other Transition Metal Nanoparticles. Pharmaceutics 2022, 14, 2286. [Google Scholar] [CrossRef] [PubMed]
- Canton, I.; Battaglia, G. Endocytosis at the nanoscale. Chem. Soc. Rev. 2012, 41, 2718. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liao, J.; Shao, X.; Li, Q.; Lin, Y. The Effect of shape on Cellular Uptake of Gold Nanoparticles in the forms of Stars, Rods, and Triangles. Sci. Rep. 2017, 7, 3827. [Google Scholar] [CrossRef]
- Akinc, A.; Battaglia, G. Exploiting Endocytosis for Nanomedicines. Cold Spring Harb. Perspect. Biol. 2013, 5, a016980. [Google Scholar] [CrossRef] [PubMed]
- Jambhrunkar, S.; Qu, Z.; Popat, A.; Yang, J.; Noonan, O.; Acauan, L.; Ahmad Nor, Y.; Yu, C.; Karmakar, S. Effect of Surface Functionality of Silica Nanoparticles on Cellular Uptake and Cytotoxicity. Mol. Pharm. 2014, 11, 3642–3655. [Google Scholar] [CrossRef]
- Varma, S.D.S.P.D. Cellular Uptake Pathways of Nanoparticles: Process of Endocytosis and Factors Affecting their Fate. Curr. Pharm. Biotechnol. 2022, 23, 679–706. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, M.S.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem. Soc. Rev. 2021, 50, 5397–5434. [Google Scholar] [CrossRef]
- Larkin, J.M.; Brown, M.S.; Goldstein, J.L.; Anderson, R.G.W. Depletion of intracellular potassium arrests coated pit formation and receptor-mediated endocytosis in fibroblasts. Cell 1983, 33, 273–285. [Google Scholar] [CrossRef]
- Daukas, G.; Zigmond, S.H. Inhibition of receptor-mediated but not fluid-phase endocytosis in polymorphonuclear leukocytes. J. Cell Biol. 1985, 101, 1673–1679. [Google Scholar] [CrossRef]
- Cosson, P.; de Curtis, I.; Pouysségur, J.; Griffiths, G.; Davoust, J. Low cytoplasmic pH inhibits endocytosis and transport from the trans-Golgi network to the cell surface. J. Cell Biol. 1989, 108, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, H.; Kishimoto, Y.; Lee, Y.C. Temperature effect on endocytosis and exocytosis by rabbit alveolar macrophages. J. Biol. Chem. 1989, 264, 15445–15450. [Google Scholar] [CrossRef]
- Roth, T.F.; Porter, K.R. Yolk protein uptake in the oocyte of the mosquito Aedes aegypti L. J. Cell Biol. 1964, 20, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Pearse, B.M.F. Coated vesicles from pig brain: Purification and biochemical characterization. J. Mol. Biol. 1975, 97, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, M.; Roux, A. Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef]
- Mettlen, M.; Chen, P.-H.; Srinivasan, S.; Danuser, G.; Schmid, S.L. Regulation of Clathrin-Mediated Endocytosis. Annu. Rev. Biochem. 2018, 87, 871–896. [Google Scholar] [CrossRef]
- Kirchhausen, T. Clathrin. Annu. Rev. Biochem. 2000, 69, 699–727. [Google Scholar] [CrossRef]
- Kirchhausen, T.; Owen, D.; Harrison, S.C. Molecular Structure, Function, and Dynamics of Clathrin-Mediated Membrane Traffic. Cold Spring Harb. Perspect. Biol. 2014, 6, a016725. [Google Scholar] [CrossRef]
- Kirchhausen, T.; Toyoda, T. Immunoelectron microscopic evidence for the extended conformation of light chains in clathrin trimers. J. Biol. Chem. 1993, 268, 10268–10273. [Google Scholar] [CrossRef]
- Margarucci, L.; Monti, M.C.; Fontanella, B.; Riccio, R.; Casapullo, A. Chemical proteomics reveals bolinaquinone as a clathrin-mediated endocytosis inhibitor. Mol. BioSyst. 2011, 7, 480–485. [Google Scholar] [CrossRef]
- Elkin, S.R.; Oswald, N.W.; Reed, D.K.; Mettlen, M.; MacMillan, J.B.; Schmid, S.L. Ikarugamycin: A Natural Product Inhibitor of Clathrin-Mediated Endocytosis. Traffic 2016, 17, 1139–1149. [Google Scholar] [CrossRef]
- Dejonghe, W.; Sharma, I.; Denoo, B.; De Munck, S.; Lu, Q.; Mishev, K.; Bulut, H.; Mylle, E.; De Rycke, R.; Vasileva, M.; et al. Disruption of endocytosis through chemical inhibition of clathrin heavy chain function. Nat. Chem. Biol. 2019, 15, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.I. Pharmacological Inhibition of Endocytic Pathways: Is It Specific Enough to Be Useful? Exocytosis Endocytosis 2008, 440, 15–33. [Google Scholar] [CrossRef]
- Slepnev, V.I.; De Camilli, P. Accessory factors in clathrin-dependent synaptic vesicle endocytosis. Nat. Rev. Neurosci. 2000, 1, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Haar, E.T.; Musacchio, A.; Harrison, S.C.; Kirchhausen, T. Atomic Structure of Clathrin. Cell 1998, 95, 563–573. [Google Scholar] [CrossRef]
- Collette, J.R.; Chi, R.J.; Boettner, D.R.; Fernandez-Golbano, I.M.; Plemel, R.; Merz, A.J.; Geli, M.I.; Traub, L.M.; Lemmon, S.K. Clathrin Functions in the Absence of the Terminal Domain Binding Site for Adaptor-associated Clathrin-Box Motifs. Mol. Biol. Cell 2009, 20, 3401–3413. [Google Scholar] [CrossRef]
- Lemmon, S.K.; Traub, L.M. Getting in Touch with the Clathrin Terminal Domain. Traffic 2012, 13, 511–519. [Google Scholar] [CrossRef]
- Knuehl, C.; Chen, C.-Y.; Manalo, V.; Hwang, P.K.; Ota, N.; Brodsky, F.M. Novel Binding Sites on Clathrin and Adaptors Regulate Distinct Aspects of Coat Assembly. Traffic 2006, 7, 1688–1700. [Google Scholar] [CrossRef]
- Chen, Z.; Mino, R.E.; Mettlen, M.; Michaely, P.; Bhave, M.; Reed, D.K.; Schmid, S.L. Wbox2: A clathrin terminal domain–derived peptide inhibitor of clathrin-mediated endocytosis. J. Cell Biol. 2020, 219, e201908189. [Google Scholar] [CrossRef]
- Muenzner, J.; Traub, L.M.; Kelly, B.T.; Graham, S.C. Cellular and viral peptides bind multiple sites on the N-terminal domain of clathrin. Traffic 2017, 18, 44–57. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; McCluskey, A. In Silico Docking, Molecular Dynamics and Binding Energy Insights into the Bolinaquinone-Clathrin Terminal Domain Binding Site. Molecules 2014, 19, 6609–6622. [Google Scholar] [CrossRef] [PubMed]
- Ghods, A.; Gilbert, J.; Baker, J.R.; Russell, C.C.; Sakoff, J.A.; McCluskey, A. A focused library synthesis and cytotoxicity of quinones derived from the natural product bolinaquinone. R. Soc. Open Sci. 2018, 5, 171189. [Google Scholar] [CrossRef] [PubMed]
- von Kleist, L.; Stahlschmidt, W.; Bulut, H.; Gromova, K.; Puchkov, D.; Robertson, M.J.; MacGregor, K.A.; Tomilin, N.; Pechstein, A.; Chau, N.; et al. Role of the Clathrin Terminal Domain in Regulating Coated Pit Dynamics Revealed by Small Molecule Inhibition. Cell 2011, 146, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Williamson, C.D.; Cole, N.B.; Donaldson, J.G. Pitstop 2 Is a Potent Inhibitor of Clathrin-Independent Endocytosis. PLoS ONE 2012, 7, e45799. [Google Scholar] [CrossRef]
- Obar, R.A.; Collins, C.A.; Hammarback, J.A.; Shpetner, H.S.; Vallee, R.B. Molecular cloning of the microtubule-associated mechanochemical enzyme dynamin reveals homology with a new family of GTP-binding proteins. Nature 1990, 347, 256–261. [Google Scholar] [CrossRef]
- Ramachandran, R.; Schmid, S.L. The dynamin superfamily. Curr. Biol. 2018, 28, R411–R416. [Google Scholar] [CrossRef]
- Hinshaw, J.E.; Schmid, S.L. Dynamin self-assembles into rings suggesting a mechanism for coated vesicle budding. Nature 1995, 374, 190–192. [Google Scholar] [CrossRef]
- Chappie, J.S.; Acharya, S.; Leonard, M.; Schmid, S.L.; Dyda, F. G domain dimerization controls dynamin’s assembly-stimulated GTPase activity. Nature 2010, 465, 435–440. [Google Scholar] [CrossRef]
- Faelber, K.; Posor, Y.; Gao, S.; Held, M.; Roske, Y.; Schulze, D.; Haucke, V.; Noé, F.; Daumke, O. Crystal structure of nucleotide-free dynamin. Nature 2011, 477, 556–560. [Google Scholar] [CrossRef]
- Danino, D.; Moon, K.-H.; Hinshaw, J.E. Rapid constriction of lipid bilayers by the mechanochemical enzyme dynamin. J. Struct. Biol. 2004, 147, 259–267. [Google Scholar] [CrossRef]
- Hinshaw, J.E. Dynamin and Its Role in Membrane Fission. Annu. Rev. Cell Dev. Biol. 2000, 16, 483–519. [Google Scholar] [CrossRef]
- Shpetner, H.S.; Vallee, R.B. Identification of dynamin, a novel mechanochemical enzyme that mediates interactions between microtubules. Cell 1989, 59, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Praefcke, G.J.K.; McMahon, H.T. The dynamin superfamily: Universal membrane tubulation and fission molecules? Nat. Rev. Mol. Cell Biol. 2004, 5, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Garcia, F.; McNiven, M.A. Differential Distribution of Dynamin Isoforms in Mammalian Cells. Mol. Biol. Cell 1998, 9, 2595–2609. [Google Scholar] [CrossRef] [PubMed]
- Ford, M.G.J.; Jenni, S.; Nunnari, J. The crystal structure of dynamin. Nature 2011, 477, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.M.; De Camilli, P. Dynamin, a membrane-remodelling GTPase. Nat. Rev. Mol. Cell Biol. 2012, 13, 75–88. [Google Scholar] [CrossRef]
- Gasper, R.; Meyer, S.; Gotthardt, K.; Sirajuddin, M.; Wittinghofer, A. It takes two to tango: Regulation of G proteins by dimerization. Nat. Rev. Mol. Cell Biol. 2009, 10, 423–429. [Google Scholar] [CrossRef]
- Niemann, H.H. Crystal structure of a dynamin GTPase domain in both nucleotide-free and GDP-bound forms. EMBO J. 2001, 20, 5813–5821. [Google Scholar] [CrossRef]
- Bramkamp, M. Structure and function of bacterial dynamin-like proteins. Biol. Chem. 2012, 393, 1203–1214. [Google Scholar] [CrossRef]
- Ramachandran, R.; Pucadyil, T.J.; Liu, Y.W.; Acharya, S.; Leonard, M.; Lukiyanchuk, V.; Schmid, S.L. Membrane Insertion of the Pleckstrin Homology Domain Variable Loop 1 Is Critical for Dynamin-catalyzed Vesicle Scission. Mol. Biol. Cell 2009, 20, 4630–4639. [Google Scholar] [CrossRef]
- Ferguson, K. Crystal structure at 2.2 Å resolution of the pleckstrin homology domain from human dynamin. Cell 1994, 79, 199–209. [Google Scholar] [CrossRef]
- Zheng, J.; Cahill, S.M.; Lemmon, M.A.; Fushman, D.; Schlessinger, J.; Cowburn, D. Identification of the Binding Site for Acidic Phospholipids on the PH Domain of Dynamin: Implications for Stimulation of GTPase Activity. J. Mol. Biol. 1996, 255, 14–21. [Google Scholar] [CrossRef]
- Vallis, Y.; Wigge, P.; Marks, B.; Evans, P.R.; McMahon, H.T. Importance of the pleckstrin homology domain of dynamin in clathrin-mediated endocytosis. Curr. Biol. 1999, 9, 257–263. [Google Scholar] [CrossRef] [PubMed]
- McPherson, P. Regulatory Role of SH3 Domain-mediated Protein–Protein Interactions in Synaptic Vesicle Endocytosis. Cell Signal 1999, 11, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Anggono, V.; Smillie, K.J.; Graham, M.E.; Valova, V.A.; Cousin, M.A.; Robinson, P.J. Syndapin I is the phosphorylation-regulated dynamin I partner in synaptic vesicle endocytosis. Nat. Neurosci. 2006, 9, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Lundmark, R.; Carlsson, S.R. Regulated Membrane Recruitment of Dynamin-2 Mediated by Sorting Nexin 9. J. Biol. Chem. 2004, 279, 42694–42702. [Google Scholar] [CrossRef]
- Shpetner, H.S.; Herskovits, J.S.; Vallee, R.B. A Binding Site for SH3 Domains Targets Dynamin to Coated Pits. J. Biol. Chem. 1996, 271, 13–16. [Google Scholar] [CrossRef]
- Clayton, E.L.; Anggono, V.; Smillie, K.J.; Chau, N.; Robinson, P.J.; Cousin, M.A. The Phospho-Dependent Dynamin-Syndapin Interaction Triggers Activity-Dependent Bulk Endocytosis of Synaptic Vesicles. J. Neurosci. 2009, 29, 7706–7717. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, Z.; Chung, M. MUC1 intra-cellular trafficking is clathrin, dynamin, and rab5 dependent. Biochem. Biophys. Res. Commun. 2008, 376, 688–693. [Google Scholar] [CrossRef]
- Cousin, M.A.; Robinson, P.J. The dephosphins: Dephosphorylation by calcineurin triggers synaptic vesicle endocytosis. Trends Neurosci. 2001, 24, 659–665. [Google Scholar] [CrossRef]
- Henley, J.R.; Krueger, E.W.A.; Oswald, B.J.; McNiven, M.A. Dynamin-mediated Internalization of Caveolae. J. Cell Biol. 1998, 141, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Gold, E.S.; Underhill, D.M.; Morrissette, N.S.; Guo, J.; McNiven, M.A.; Aderem, A. Dynamin 2 Is Required for Phagocytosis in Macrophages. J. Exp. Med. 1999, 190, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- van Dam, E.M.; Stoorvogel, W. Dynamin-dependent Transferrin Receptor Recycling by Endosome-derived Clathrin-coated Vesicles. Mol. Biol. Cell 2002, 13, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Chircop, M. Clathrin-Mediated Endocytic Proteins are Involved in Regulating Mitotic Progression and Completion. Traffic 2012, 13, 1628–1641. [Google Scholar] [CrossRef]
- Lu, J.; Helton, T.D.; Blanpied, T.A.; Rácz, B.; Newpher, T.M.; Weinberg, R.J.; Ehlers, M.D. Postsynaptic Positioning of Endocytic Zones and AMPA Receptor Cycling by Physical Coupling of Dynamin-3 to Homer. Neuron 2007, 55, 874–889. [Google Scholar] [CrossRef]
- Hill, T.A.; Odell, L.R.; Quan, A.; Abagyan, R.; Ferguson, G.; Robinson, P.J.; McCluskey, A. Long chain amines and long chain ammonium salts as novel inhibitors of dynamin GTPase activity. Bioorg. Med. Chem. Lett. 2004, 14, 3275–3278. [Google Scholar] [CrossRef]
- Quan, A.; McGeachie, A.B.; Keating, D.J.; van Dam, E.M.; Rusak, J.; Chau, N.; Malladi, C.S.; Chen, C.; McCluskey, A.; Cousin, M.A.; et al. Myristyl Trimethyl Ammonium Bromide and Octadecyl Trimethyl Ammonium Bromide Are Surface-Active Small Molecule Dynamin Inhibitors that Block Endocytosis Mediated by Dynamin I or Dynamin II. Mol. Pharmacol. 2007, 72, 1425–1439. [Google Scholar] [CrossRef]
- Daniel, J.A.; Chau, N.; Abdel-Hamid, M.K.; Hu, L.; von Kleist, L.; Whiting, A.; Krishnan, S.; Maamary, P.; Joseph, S.R.; Simpson, F.; et al. Phenothiazine-Derived Antipsychotic Drugs Inhibit Dynamin and Clathrin-Mediated Endocytosis. Traffic 2015, 16, 635–654. [Google Scholar] [CrossRef]
- Otomo, M.; Takahashi, K.; Miyoshi, H.; Osada, K.; Nakashima, H.; Yamaguchi, N. Some Selective Serotonin Reuptake Inhibitors Inhibit Dynamin I Guanosine Triphosphatase (GTPase). Biol. Pharm. Bull 2008, 31, 1489–1495. [Google Scholar] [CrossRef]
- Takahashi, K.; Miyoshi, H.; Otomo, M.; Osada, K.; Yamaguchi, N.; Nakashima, H. Suppression of dynamin GTPase activity by sertraline leads to inhibition of dynamin-dependent endocytosis. Biochem. Biophys. Res. Commun. 2010, 391, 382–387. [Google Scholar] [CrossRef]
- Zhang, J.; Lawrance, G.A.; Chau, N.; Robinson, P.J.; McCluskey, A. From Spanish fly to room-temperature ionic liquids (RTILs): Synthesis, thermal stability and inhibition of dynamin 1 GTPase by a novel class of RTILs. New J. Chem. 2008, 32, 28–36. [Google Scholar] [CrossRef]
- Hill, T.; Odell, L.R.; Edwards, J.K.; Graham, M.E.; McGeachie, A.B.; Rusak, J.; Quan, A.; Abagyan, R.; Scott, J.L.; Robinson, P.J.; et al. Small Molecule Inhibitors of Dynamin I GTPase Activity: Development of Dimeric Tyrphostins. J. Med. Chem. 2005, 48, 7781–7788. [Google Scholar] [CrossRef] [PubMed]
- Odell, L.R.; Robertson, M.J.; Young, K.A.; McGeachie, A.B.; Quan, A.; Robinson, P.J.; McCluskey, A. Prodrugs of the Archetypal Dynamin Inhibitor Bis-T-22. ChemMedChem 2022, 17, e202200400. [Google Scholar] [CrossRef] [PubMed]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a Cell-Permeable Inhibitor of Dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef]
- Hill, T.A.; Gordon, C.P.; McGeachie, A.B.; Venn-Brown, B.; Odell, L.R.; Chau, N.; Quan, A.; Mariana, A.; Sakoff, J.A.; Chircop, M.; et al. Inhibition of Dynamin Mediated Endocytosis by the Dynoles—Synthesis and Functional Activity of a Family of Indoles. J. Med. Chem. 2009, 52, 3762–3773. [Google Scholar] [CrossRef]
- Hill, T.A.; Mariana, A.; Gordon, C.P.; Odell, L.R.; Robertson, M.J.; McGeachie, A.B.; Chau, N.; Daniel, J.A.; Gorgani, N.N.; Robinson, P.J.; et al. Iminochromene Inhibitors of Dynamins I and II GTPase Activity and Endocytosis. J. Med. Chem. 2010, 53, 4094–4102. [Google Scholar] [CrossRef]
- Odell, L.R.; Howan, D.; Gordon, C.P.; Robertson, M.J.; Chau, N.; Mariana, A.; Whiting, A.E.; Abagyan, R.; Daniel, J.A.; Gorgani, N.N.; et al. The Pthaladyns: GTP Competitive Inhibitors of Dynamin I and II GTPase Derived from Virtual Screening. J. Med. Chem. 2010, 53, 5267–5280. [Google Scholar] [CrossRef]
- Mermer, A. The Importance of Rhodanine Scaffold in Medicinal Chemistry: A Comprehensive Overview. Mini-Rev. Med. Chem. 2021, 21, 738–789. [Google Scholar] [CrossRef]
- Robertson, M.J.; Hadzic, G.; Ambrus, J.; Pomè, D.Y.; Hyde, E.; Whiting, A.; Mariana, A.; von Kleist, L.; Chau, N.; Haucke, V.; et al. The Rhodadyns, a New Class of Small Molecule Inhibitors of Dynamin GTPase Activity. ACS Med. Chem. Lett. 2012, 3, 352–356. [Google Scholar] [CrossRef]
- Gordon, C.P.; Venn-Brown, B.; Robertson, M.J.; Young, K.A.; Chau, N.; Mariana, A.; Whiting, A.; Chircop, M.; Robinson, P.J.; McCluskey, A. Development of Second-Generation Indole-Based Dynamin GTPase Inhibitors. J. Med. Chem. 2013, 56, 46–59. [Google Scholar] [CrossRef]
- McGeachie, A.B.; Odell, L.R.; Quan, A.; Daniel, J.A.; Chau, N.; Hill, T.A.; Gorgani, N.N.; Keating, D.J.; Cousin, M.A.; Van Dam, E.M.; et al. Pyrimidyn Compounds: Dual-Action Small Molecule Pyrimidine-Based Dynamin Inhibitors. ACS Chem. Biol. 2013, 8, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, A.; Daniel, J.A.; Hadzic, G.; Chau, N.; Clayton, E.L.; Mariana, A.; Whiting, A.; Gorgani, N.N.; Lloyd, J.; Quan, A.; et al. Building a Better Dynasore: The Dyngo Compounds Potently Inhibit Dynamin and Endocytosis. Traffic 2013, 14, 1272–1289. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, M.K.; Macgregor, K.A.; Odell, L.R.; Chau, N.; Mariana, A.; Whiniting, A.; Robinson, P.J.; McCluskey, A. 1,8-Naphthalimide derivatives: New leads against dynamin I GTPase activity. Org. Biomol. Chem. 2015, 13, 8016–8028. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, K.A.; Abdel-Hamid, M.K.; Odell, L.R.; Chau, N.; Whiting, A.; Robinson, P.J.; McCluskey, A. Development of quinone analogues as dynamin GTPase inhibitors. Eur. J. Med. Chem. 2014, 85, 191–206. [Google Scholar] [CrossRef]
- Camarero, N.; Trapero, A.; Pérez-Jiménez, A.; Macia, E.; Gomila-Juaneda, A.; Martín-Quirós, A.; Nevola, L.; Llobet, A.; Llebaria, A.; Hernando, J.; et al. Correction: Photoswitchable dynasore analogs to control endocytosis with light. Chem. Sci. 2020, 11, 9712. [Google Scholar] [CrossRef]
- Hamm-Alvarez, S.F.; Sonee, M.; Loran-Goss, K.; Shen, W.C. Paclitaxel and nocodazole differentially alter endocytosis in cultured cells. Pharm. Res. 1996, 13, 1647–1656. [Google Scholar] [CrossRef]
- Zhang, L.W.; Monteiro-Riviere, N.A. Mechanisms of Quantum Dot Nanoparticle Cellular Uptake. Toxicol. Sci. 2009, 110, 138–155. [Google Scholar] [CrossRef]
- Vercauteren, D.; Vandenbroucke, R.E.; Jones, A.T.; Rejman, J.; Demeester, J.; De Smedt, S.C.; Sanders, N.N.; Braeckmans, K. The Use of Inhibitors to Study Endocytic Pathways of Gene Carriers: Optimization and Pitfalls. Mol. Ther. 2010, 18, 561–569. [Google Scholar] [CrossRef]
- Gibson, A.E.; Noel, R.J.; Herlihy, J.T.; Ward, W.F. Phenylarsine oxide inhibition of endocytosis: Effects on asialofetuin internalization. Am. J. Physiol.-Cell Physiol. 1989, 257, C182–C184. [Google Scholar] [CrossRef]
- Park, R.J.; Shen, H.; Liu, L.; Liu, X.; Ferguson, S.M.; De Camilli, P. Dynamin triple knockout cells reveal off target effects of commonly used dynamin inhibitors. J. Cell Sci. 2013, 126, 5305–5312. [Google Scholar] [CrossRef]
- Basagiannis, D.; Zografou, S.; Goula, E.; Gkeka, D.; Kolettas, E.; Christoforidis, S. Chemical inhibitors of dynamin exert differential effects in vegf signaling. Cells 2021, 10, 997. [Google Scholar] [CrossRef] [PubMed]
- Persaud, A.; Cormerais, Y.; Pouyssegur, J.; Rotin, D. Dynamin inhibitors block activation of mTORC1 by amino acids independently of dynamin. J. Cell Sci. 2018, 131, jcs211755. [Google Scholar] [CrossRef]
- Preta, G.; Lotti, V.; Cronin, J.G.; Sheldon, I.M. Protective role of the dynamin inhibitor Dynasore against the cholesterol-dependent cytolysin of Trueperella pyogenes. FASEB J. 2015, 29, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Sultana, J.; Crisafulli, S.; Gabbay, F.; Lynn, E.; Shakir, S.; Trifirò, G. Challenges for Drug Repurposing in the COVID-19 Pandemic Era. Front. Pharmacol. 2020, 11, 1657. [Google Scholar] [CrossRef] [PubMed]
- Study of Ruxolitinib Plus Simvastatin in the Prevention and Treatment of Respiratory Failure of COVID-19—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04348695 (accessed on 17 March 2023).
- Early Phase Human Drug Trial to Investigate Dynamin 101 (DYN101) in Patients ≥ 16 Years with Centronuclear Myopathies—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04033159 (accessed on 17 March 2023).
- Survey of Inhibitors in Plasma-Product Exposed Toddlers—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01064284 (accessed on 17 March 2023).
- Peyvandi, F.; Mannucci, P.M.; Garagiola, I.; El-Beshlawy, A.; Elalfy, M.; Ramanan, V.; Eshghi, P.; Hanagavadi, S.; Varadarajan, R.; Karimi, M.; et al. A Randomized Trial of Factor VIII and Neutralizing Antibodies in Hemophilia A. N. Engl. J. Med. 2016, 374, 2054–2064. [Google Scholar] [CrossRef] [PubMed]
- Post Exposure Prophylaxis in Healthcare Workers Exposed to COVID-19 Patients—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04437693 (accessed on 17 March 2023).
- Akhtar, S.; Das, J.K.; Ismail, T.; Wahid, M.; Saeed, W.; Bhutta, Z.A. Nutritional perspectives for the prevention and mitigation of COVID-19. Nutr. Rev. 2021, 79, 289–300. [Google Scholar] [CrossRef]
- Hydroxychloroquine Post Exposure Prophylaxis for Coronavirus Disease (COVID-19)—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04318444 (accessed on 17 March 2023).
- Hydroxychloroquine Treatment for Severe COVID-19 Pulmonary Infection (HYDRA Trial)—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04315896 (accessed on 17 March 2023).
- Hydroxychloroquine Chemoprophylaxis in Healthcare Personnel in Contact with COVID-19 Patients (PHYDRA Trial)—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04318015 (accessed on 17 March 2023).
- Post-exposure Prophylaxis/Preemptive Therapy for SARS-Coronavirus-2—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04308668 (accessed on 17 March 2023).
- Norwegian Coronavirus Disease 2019 Study—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04316377 (accessed on 17 March 2023).
- Lyngbakken MNBerdal, J.E.; Eskesen AKvale, D.; Olsen, I.C.; Rangberg, A.; Jonassen, C.M.; Omland, T.; Rosjo, H.; Dalgard, O. Norwegian Coronavirus Disease 2019 (NO COVID-19) Pragmatic Open label Study to assess early use of hydroxychloroquine sulphate in moderately severe hospitalised patients with coronavirus disease 2019: A structured summary of a study protocol for a randomised controlled trial. Trials 2020, 21, 485. [Google Scholar] [CrossRef]
- Repurposing of Chlorpromazine in COVID-19 Treatment—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04366739 (accessed on 17 March 2023).
- Chloroquine Phosphate Against Infection by the Novel Coronavirus SARS-CoV-2 (COVID-19): The HOPE Open-Label, Non Randomized Clinical Trial—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04344951 (accessed on 17 March 2023).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathology | Type of Endocytosis | Ligands and Receptors | Molecular Mechanism | References |
|---|---|---|---|---|
| Cardiac disorders | Caveolin-dependent | eNOS, Cav-1, PCK, L-arginine, L-citrulline, ERK 1/2 | After attaching to the CSD (caveolin scaffolding domain), eNOS remains dormant, which lowers NO production. The catalytic domain of eNOS, which is thought to act as the enzyme’s on/off switch, contains a putative binding domain for Cav-1. Cav-1 binding is prevented by mutagenesis in eNOS, although Cav-1’s CSD is less effective at controlling eNOS activity in vivo | [113] |
| Lipid disorders and atherosclerosis | Clathrin-mediated | RhoA, PKC, LDLR, AP-2, ARH | After binding to the LDL receptor, LDL particles are internalized by clathrin-coated pits into vesicles. Internalization of LDLR requires the presence of AP-2 protein. LDLR interacts with AP-2 indirectly through the adaptor protein ARH. After the fusion of endocytosed vesicles with early endosomes, a drop in pH induces LDL dissociation from its receptor. LDLR is recycled back to the surface of the cell, and LDLs are directed to lysosomes. | [114] |
| Respiratory diseases | Caveolin-dependent | HSP90, Gq protein, Cav-1, Cav-3, calmodulin, L-arginine, NO | Caveolins participate in Ca2+ handling in airway smooth muscle. Caveolins probably promote smooth muscle contraction by regulating Gq protein function and phosphoinositide metabolism. In addition, recruitment across the plasma membrane of RhoA and PKC is dependent on Cav-1. Negative allosteric modulation of eNOS by Cav-1 and 3 competes with positive allosteric modulation of eNOS by Ca2+/calmodulin and HSP90 complexes. Thus, in the presence of elevated levels of Ca2+/calmodulin and HSP90, the inhibitory effect of Cav-1 on NOS activity can be completely reversed. | [115,116,117,118] |
| Cancer | Clathrin-mediated | Cdc42, Cbl, b-Pix EGFR, Rac1, Src, RhoA | Cdc42 bound to GTP forms a complex mediated by the adaptor protein b-Pix and Cbl. As a consequence, Cbl cannot react with EGFR; this promotes ubiquitination and degradation of the receptor. Cdc42 belongs to the small GTPase family, which represents critical effectors of pathways and Ras-derived signaling that have a proven role in tumor cell invasiveness and metastasis. | [119] |
| Target | Compound | Condition or Disease | Mechanism | Number | Status | References |
|---|---|---|---|---|---|---|
| Blocking clathrin-dependent endocytosis | Ruxolitinib and simvastatin | Coronavirus Infection | Block the entry process used by COVID-19 | NCT04348695 | Phase 2 | [244,245] |
| Dynamin2 | DYN101 | Centronuclear Myopathy | Antisense oligonucleotide directed against dynamin2 pre-mRNA | NCT04033159 | Phase 2 | [246] |
| Blocking endocytosis in human dendritic cells | Chaperone molecule for procoagulant factor VIII (FVIII) | Hemophilia A | VWF protects FVIII from being endocytosed by human dendritic cells and subsequently presented to FVIII-specific T cells | NCT01064284 | Phase 4 | [247,248] |
| Reducing the expression of phosphatidylinositol-binding clathrin assembly protein and blocking the clathrin-mediated endocytosis | Hydroxychloroquine (HCQ) | Coronavirus Infection (COVID-19) | Inhibits the ability of viruses to escape into the host cell and start replicating | NCT04437693 | Phase 3 | [249] |
| Coronavirus Infection (COVID-19) | NCT04318444 | Phase 3 | [250,251] | |||
| COVID-19, Severe Acute Respiratory Syndrome | NCT04315896 | Phase 3 | [252] | |||
| COVID-19, Severe Acute Respiratory Syndrome | NCT04318015 | Phase 3 | [253] | |||
| Coronavirus Infection (COVID-19) | NCT04308668 | Phase 3 | [254] | |||
| Coronavirus Infection (COVID-19) | NCT04316377 | Phase 4 | [255,256] | |||
| Blocking clathrin-mediated endocytosis | Chlorpromazine (CPZ) | Coronavirus Infection (COVID-19) | CPZ affects the translocation of the clathrin and AP2 from the cell surface to intracellular endosomes | NCT04366739 | Phase 3 | [257] |
| Inhibiting endosome acidification | UNIKINON (Chloroquine phosphate) | Pneumonia, Coronavirus Infection (COVID-19) | Blocks the endosomal-mediated viral entry | NCT04344951 | Phase 2 | [258] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szewczyk-Roszczenko, O.K.; Roszczenko, P.; Shmakova, A.; Finiuk, N.; Holota, S.; Lesyk, R.; Bielawska, A.; Vassetzky, Y.; Bielawski, K. The Chemical Inhibitors of Endocytosis: From Mechanisms to Potential Clinical Applications. Cells 2023, 12, 2312. https://doi.org/10.3390/cells12182312
Szewczyk-Roszczenko OK, Roszczenko P, Shmakova A, Finiuk N, Holota S, Lesyk R, Bielawska A, Vassetzky Y, Bielawski K. The Chemical Inhibitors of Endocytosis: From Mechanisms to Potential Clinical Applications. Cells. 2023; 12(18):2312. https://doi.org/10.3390/cells12182312
Chicago/Turabian StyleSzewczyk-Roszczenko, Olga Klaudia, Piotr Roszczenko, Anna Shmakova, Nataliya Finiuk, Serhii Holota, Roman Lesyk, Anna Bielawska, Yegor Vassetzky, and Krzysztof Bielawski. 2023. "The Chemical Inhibitors of Endocytosis: From Mechanisms to Potential Clinical Applications" Cells 12, no. 18: 2312. https://doi.org/10.3390/cells12182312