
A New Understanding of Long Non-Coding RNA in Hepatocellular Carcinoma—From m6A Modification to Blood Biomarkers

Abstract
:1. Introduction
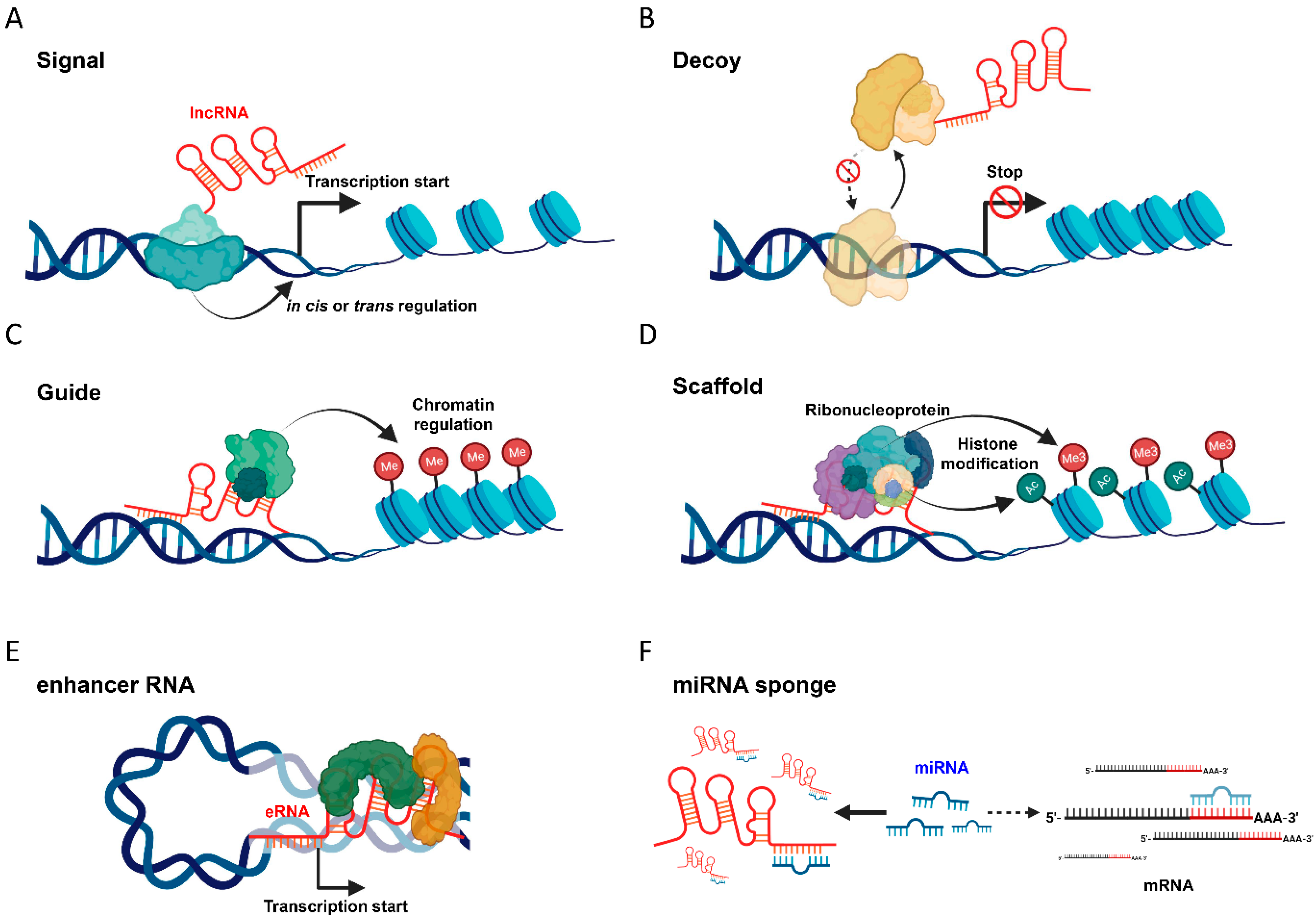
2. Understanding lncRNA: Its Definition and Mechanism of Action
2.1. Signal
2.2. Decoy
2.3. Guide
2.4. Scaffold
2.5. Enhancer RNA
2.6. MiRNA Sponge
3. Pivotal Role of lncRNA in Hepatocarcinogenesis
3.1. HOTAIR (HOX Transcript Antisense RNA)
3.2. NEAT1 (Nuclear Enriched Abundant Transcript 1)
3.3. HULC (Highly Upregulated in Liver Cancer)
3.4. MALAT1 (Metastasis Associated Lung Adenocarcinoma Transcript 1)
4. LncRNAs as Key Modulators of the HCC Tumor Microenvironment
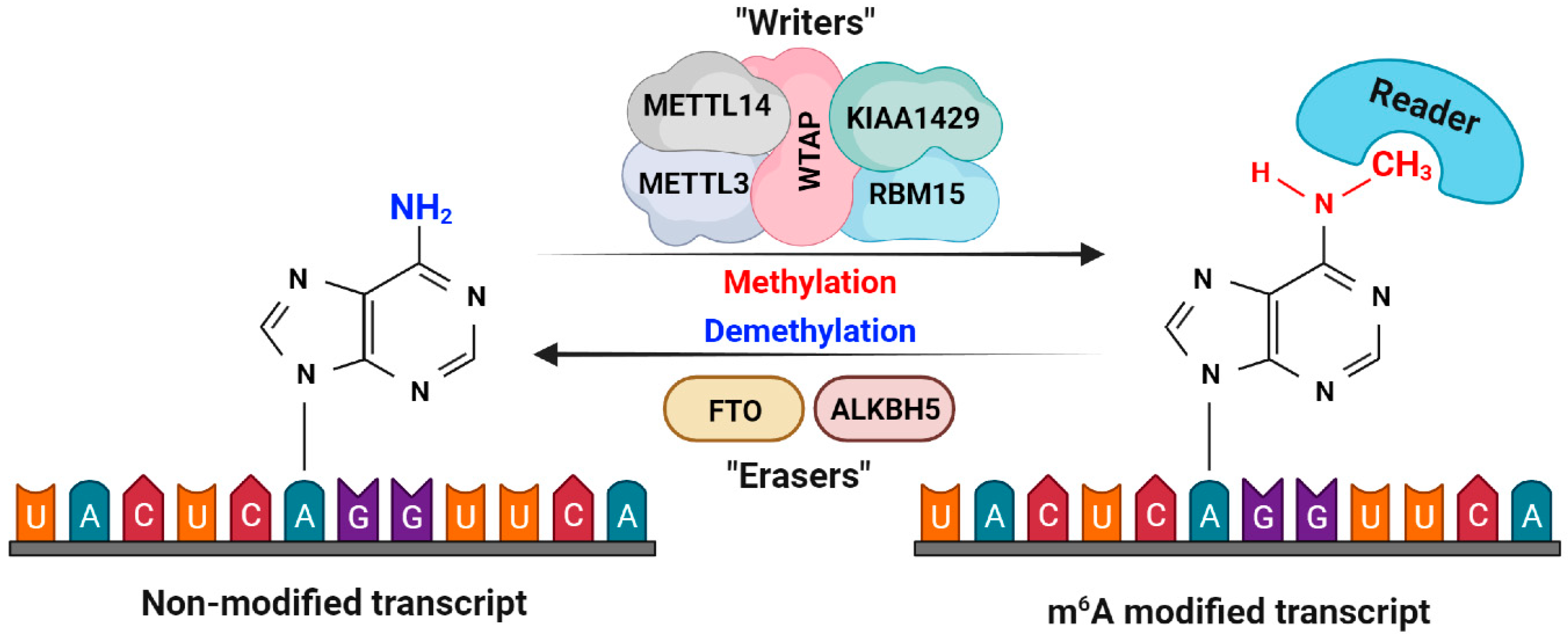
5. Implications of m6A Modification on lncRNA in HCC
6. LncRNAs as Serum Biomarkers in Liver Cancer
7. Summary and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eddy, S.R. Non–coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef]
- Zhu, Z.; Ma, Y.; Li, Y.; Li, P.; Cheng, Z.; Li, H.; Zhang, L.; Tang, Z. The comprehensive detection of miRNA, lncRNA, and circRNA in regulation of mouse melanocyte and skin development. Biol. Res. 2020, 53, 4. [Google Scholar] [CrossRef] [PubMed]
- Bahn, J.H.; Zhang, Q.; Li, F.; Chan, T.M.; Lin, X.; Kim, Y.; Wong, D.T.; Xiao, X. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin. Chem. 2015, 61, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Penny, G.D.; Kay, G.F.; Sheardown, S.A.; Rastan, S.; Brockdorff, N. Requirement for Xist in X chromosome inactivation. Nature 1996, 379, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Luo, L.; Zhen, Y.; Peng, D.; Wei, C.; Zhang, X.; Liu, X.; Han, L.; Zhang, Z. The role of N6-methyladenosine-modified non-coding RNAs in the pathological process of human cancer. Cell Death Discov. 2022, 8, 325. [Google Scholar] [CrossRef]
- Yang, C.; Hu, Y.; Zhou, B.; Bao, Y.; Li, Z.; Gong, C.; Yang, H.; Wang, S.; Xiao, Y. The role of m6A modification in physiology and disease. Cell Death Dis. 2020, 11, 960. [Google Scholar] [CrossRef]
- Sivasudhan, E.; Blake, N.; Lu, Z.L.; Meng, J.; Rong, R. Dynamics of m6A RNA Methylome on the Hallmarks of Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2021, 9, 642443. [Google Scholar] [CrossRef]
- Han, P.; Chang, C.P. Long non-coding RNA and chromatin remodeling. RNA Biol. 2015, 12, 1094–1098. [Google Scholar] [CrossRef]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, D.; Hao, Y.; Wu, S.; Luo, J.; Xue, Y.; Wang, D.; Li, G.; Liu, L.; Shao, C.; et al. LncRNA CCTT-mediated RNA-DNA and RNA-protein interactions facilitate the recruitment of CENP-C to centromeric DNA during kinetochore assembly. Mol. Cell 2022, 82, 4018–4032.e4019. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Sqazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Hemls, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.A.G.; Adachi, S.; Nong, Q.D.; Adhitama, N.; Matsuura, T.; Natsume, T.; Wada, T.; Kato, Y.; Watanabe, H. Sense-overlapping lncRNA as a decoy of translational repressor protein for dimorphic gene expression. PLoS Genet. 2021, 17, e1009683. [Google Scholar] [CrossRef]
- Hung, T.; Wang, Y.; Lin, M.F.; Koegel, A.K.; Kotake, Y.; Grant, G.D.; Horlings, H.M.; Shah, N.; Umbricht, C.; Wang, P.; et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat. Genet. 2011, 43, 621–629. [Google Scholar] [CrossRef]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal 2010, 3, ra8. [Google Scholar] [CrossRef]
- Arab, K.; Park, Y.J.; Lindroth, A.M.; Schafer, A.; Oakes, C.; Weichenhan, D.; Lukanova, A.; Lundin, E.; Risch, A.; Meister, M.; et al. Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol. Cell 2014, 55, 604–614. [Google Scholar] [CrossRef]
- Jiang, L.; Shao, C.; Wu, Q.J.; Chen, G.; Zhou, J.; Yang, B.; Li, H.; Gou, L.T.; Zhang, Y.; Wang, Y.; et al. NEAT1 scaffolds RNA-binding proteins and the Microprocessor to globally enhance pri-miRNA processing. Nat. Struct. Mol. Biol. 2017, 24, 816–824. [Google Scholar] [CrossRef]
- Kim, T.K.; Hemberg, M.; Gray, J.M. Enhancer RNAs: A class of long noncoding RNAs synthesized at enhancers. Cold Spring Harb. Perspect. Biol. 2015, 7, a018622. [Google Scholar] [CrossRef]
- Miao, Y.; Ajami, N.E.; Huang, T.S.; Lin, F.M.; Lou, C.H.; Wang, Y.T.; Li, S.; Kang, J.; Munkacsi, H.; Maurya, M.R.; et al. Enhancer-associated long non-coding RNA LEENE regulates endothelial nitric oxide synthase and endothelial function. Nat. Commun. 2018, 9, 292. [Google Scholar] [CrossRef]
- Lou, M.M.; Tang, X.Q.; Wang, G.M.; He, J.; Luo, F.; Guan, M.F.; Wang, F.; Zou, H.; Wang, J.Y.; Zhang, Q.; et al. Long noncoding RNA BS-DRL1 modulates the DNA damage response and genome stability by interacting with HMGB1 in neurons. Nat. Commun. 2021, 12, 4075. [Google Scholar] [CrossRef]
- Flores-Concha, M.; Onate, A.A. Long Non-coding RNAs in the Regulation of the Immune Response and Trained Immunity. Front. Genet. 2020, 11, 718. [Google Scholar] [CrossRef]
- Sellitto, A.; Pecoraro, G.; Giurato, G.; Nassa, G.; Rizzo, F.; Saggese, P.; Martinez, C.A.; Scafoglio, C.; Tarallo, R. Regulation of Metabolic Reprogramming by Long Non-Coding RNAs in Cancer. Cancers 2021, 13, 3485. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Q.; Liu, B.; Mei, L.; Ma, S.; Wang, S.; Wang, R.; Zhang, Y.; Niu, C.; Xiong, Z.; et al. The long noncoding RNA Synage regulates synapse stability and neuronal function in the cerebellum. Cell Death Differ. 2021, 28, 2634–2650. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Wang, J.; Zhang, L.; Li, J.; Jin, Y. LncRNA HOTAIR Promotes Cancer Stem-Like Cells Properties by Sponging miR-34a to Activate the JAK2/STAT3 Pathway in Pancreatic Ductal Adenocarcinoma. Onco Targets Ther. 2021, 14, 1883–1893. [Google Scholar] [CrossRef]
- Gao, L.; Wang, X.; Guo, S.; Xiao, L.; Liang, C.; Wang, Z.; Li, Y.; Liu, Y.; Yao, R.; Liu, Y.; et al. LncRNA HOTAIR functions as a competing endogenous RNA to upregulate SIRT1 by sponging miR-34a in diabetic cardiomyopathy. J. Cell Physiol. 2019, 234, 4944–4958. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Hu, Y.; Tang, W.; Shen, H.; Yu, Z.; Gu, J. The long noncoding RNA HOTAIR serves as a microRNA-34a-5p sponge to reduce nucleus pulposus cell apoptosis via a NOTCH1-mediated mechanism. Gene 2019, 715, 144029. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Kats, L.; Salmena, L.; Weiss, D.; Tan, S.M.; Ala, U.; Karreth, F.; Poliseno, L.; Provero, P.; Di Cunto, F.; et al. Coding-independent regulation of the tumor suppressor PTEN by competing endogenous mRNAs. Cell 2011, 147, 344–357. [Google Scholar] [CrossRef]
- Karreth, F.A.; Tay, Y.; Perna, D.; Ala, U.; Tan, S.M.; Rust, A.G.; DeNicola, G.; Webster, K.A.; Weiss, D.; Perez-Mancera, P.A.; et al. In vivo identification of tumor- suppressive PTEN ceRNAs in an oncogenic BRAF-induced mouse model of melanoma. Cell 2011, 147, 382–395. [Google Scholar] [CrossRef]
- Gu, X.; Li, M.; Jin, Y.; Liu, D.; Wei, F. Identification and integrated analysis of differentially expressed lncRNAs and circRNAs reveal the potential ceRNA networks during PDLSC osteogenic differentiation. BMC Genet. 2017, 18, 100. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Gholipour, M.; Hussen, B.M.; Taheri, M. The Impact of Long Non-Coding RNAs in the Pathogenesis of Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 649107. [Google Scholar] [CrossRef]
- Abbastabar, M.; Sarfi, M.; Golestani, A.; Khalili, E. lncRNA involvement in hepatocellular carcinoma metastasis and prognosis. EXCLI J. 2018, 17, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, M.; Kogo, R.; Shibata, K.; Sawada, G.; Takahashi, Y.; Kurashige, J.; Akiyoshi, S.; Sasaki, S.; Iwaya, T.; Sudo, T.; et al. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol. Rep. 2013, 29, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.M.; Zhu, X.; Wang, W.M.; Lu, Y.F.; Hu, B.G.; Wang, H.; Liang, W.C.; Wang, S.S.; Ko, C.H.; Waye, M.M.; et al. Hotair mediates hepatocarcinogenesis through suppressing miRNA-218 expression and activating P14 and P16 signaling. J. Hepatol. 2015, 63, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Topel, H.; Bagirsakci, E.; Comez, D.; Bagci, G.; Cakan-Akdogan, G.; Atabey, N. lncRNA HOTAIR overexpression induced downregulation of c-Met signaling promotes hybrid epithelial/mesenchymal phenotype in hepatocellular carcinoma cells. Cell Commun. Signal 2020, 18, 110. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, L.; Wu, L.M.; Lai, M.C.; Xie, H.Y.; Zhang, F.; Zheng, S.S. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol. 2011, 18, 1243–1250. [Google Scholar] [CrossRef]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1. [Google Scholar] [CrossRef]
- Fang, S.; Shen, Y.; Chen, B.; Wu, Y.; Jia, L.; Li, Y.; Zhu, Y.; Yan, Y.; Li, M.; Chen, R.; et al. H3K27me3 induces multidrug resistance in small cell lung cancer by affecting HOXA1 DNA methylation via regulation of the lncRNA HOTAIR. Ann. Transl. Med. 2018, 6, 440. [Google Scholar] [CrossRef]
- Zhang, H.; Diab, A.; Fan, H.; Mani, S.K.; Hullinger, R.; Merle, P.; Andrisani, O. PLK1 and HOTAIR Accelerate Proteasomal Degradation of SUZ12 and ZNF198 during Hepatitis B Virus-Induced Liver Carcinogenesis. Cancer Res. 2015, 75, 2363–2374. [Google Scholar] [CrossRef]
- Zhang, H.; Xing, Z.; Mani, S.K.; Bancel, B.; Durantel, D.; Zoulim, F.; Tran, E.J.; Merle, P.; Andrisani, O. RNA helicase DEAD box protein 5 regulates Polycomb repressive complex 2/Hox transcript antisense intergenic RNA function in hepatitis B virus infection and hepatocarcinogenesis. Hepatology 2016, 64, 1033–1048. [Google Scholar] [CrossRef]
- Clemson, C.M.; Hutchinson, J.N.; Sara, S.A.; Ensminger, A.W.; Fox, A.H.; Chess, A.; Lawrence, J.B. An architectural role for a nuclear noncoding RNA: NEAT1 RNA is essential for the structure of paraspeckles. Mol. Cell 2009, 33, 717–726. [Google Scholar] [CrossRef]
- Liu, Z.; Chang, Q.; Yang, F.; Liu, B.; Yao, H.W.; Bai, Z.G.; Pu, C.S.; Ma, X.M.; Yang, Y.; Wang, T.T.; et al. Long non-coding RNA NEAT1 overexpression is associated with unfavorable prognosis in patients with hepatocellular carcinoma after hepatectomy: A Chinese population-based study. Eur. J. Surg. Oncol. 2017, 43, 1697–1703. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.; Li, Z.; Wei, D.; Chen, H.; Yang, C. LncRNA, NEAT1 is a prognosis biomarker and regulates cancer progression via epithelial-mesenchymal transition in clear cell renal cell carcinoma. Cancer Biomark. 2017, 19, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, H.; Yang, P.; He, Z.Y. Prognostic role of long noncoding RNA NEAT1 in various carcinomas: A meta-analysis. Onco Targets Ther. 2017, 10, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cao, Z.; Ding, Y.; Li, Z.; Xiang, X.; Lai, R.; Sheng, Z.; Liu, Y.; Cai, W.; Hu, R.; et al. Long Non-coding RNA NEAT1 Alleviates Acute-on-Chronic Liver Failure Through Blocking TRAF6 Mediated Inflammatory Response. Front. Physiol. 2019, 10, 1503. [Google Scholar] [CrossRef] [PubMed]
- Bu, F.T.; Wang, A.; Zhu, Y.; You, H.M.; Zhang, Y.F.; Meng, X.M.; Huang, C.; Li, J. LncRNA NEAT1: Shedding light on mechanisms and opportunities in liver diseases. Liver Int. 2020, 40, 2612–2626. [Google Scholar] [CrossRef]
- Mang, Y.; Li, L.; Ran, J.; Zhang, S.; Liu, J.; Li, L.; Chen, Y.; Liu, J.; Gao, Y.; Ren, G. Long noncoding RNA NEAT1 promotes cell proliferation and invasion by regulating hnRNP A2 expression in hepatocellular carcinoma cells. Onco Targets Ther. 2017, 10, 1003–1016. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, Y.; Liu, Y.; Fang, L.; Li, L.; Sun, J.; Pan, Z.; Xin, W.; Huang, P. HIF-2alpha activated lncRNA NEAT1 promotes hepatocellular carcinoma cell invasion and metastasis by affecting the epithelial-mesenchymal transition. J. Cell Biochem. 2018, 119, 3247–3256. [Google Scholar] [CrossRef]
- Fang, L.; Sun, J.; Pan, Z.; Song, Y.; Zhong, L.; Zhang, Y.; Liu, Y.; Zheng, X.; Huang, P. Long non-coding RNA NEAT1 promotes hepatocellular carcinoma cell proliferation through the regulation of miR-129-5p-VCP-IkappaB. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G150–G156. [Google Scholar] [CrossRef]
- Wang, Z.; Zou, Q.; Song, M.; Chen, J. NEAT1 promotes cell proliferation and invasion in hepatocellular carcinoma by negative regulating miR-613 expression. Biomed. Pharmacother. 2017, 94, 612–618. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, M.; Cui, X.; O’Connell, D.; Yang, Y. Long noncoding RNA NEAT1 promotes ferroptosis by modulating the miR-362-3p/MIOX axis as a ceRNA. Cell Death Differ. 2022, 29, 1850–1863. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Ru, Y.; Chen, X.J.; Guo, W.Z.; Gao, S.G.; Qi, Y.J.; Chen, P.; Feng, X.S.; Zhang, S.J. NEAT1_2-SFPQ axis mediates cisplatin resistance in liver cancer cells in vitro. Onco Targets Ther. 2018, 11, 5695–5702. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xia, X. Long noncoding RNA NEAT1 suppresses sorafenib sensitivity of hepatocellular carcinoma cells via regulating miR-335-c-Met. J. Cell Physiol. 2019, 234, 14999–15009. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Wu, H.; Ni, P.; Gu, Z.; Qiao, Y.; Chen, N.; Sun, F.; Fan, Q. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucleic Acids Res. 2010, 38, 5366–5383. [Google Scholar] [CrossRef]
- Li, D.; Liu, X.; Zhou, J.; Hu, J.; Zhang, D.; Liu, J.; Qiao, Y.; Zhan, Q. Long noncoding RNA HULC modulates the phosphorylation of YB-1 through serving as a scaffold of extracellular signal-regulated kinase and YB-1 to enhance hepatocarcinogenesis. Hepatology 2017, 65, 1612–1627. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Huang, D.; Yang, F.; Tian, M.; Wang, Y.; Shen, D.; Wang, Q.; Chen, Q.; Zhang, L. Long Noncoding RNA Highly Upregulated in Liver Cancer Regulates the Tumor Necrosis Factor-alpha-Induced Apoptosis in Human Vascular Endothelial Cells. DNA Cell Biol. 2016, 35, 296–300. [Google Scholar] [CrossRef]
- Du, Y.; Kong, G.; You, X.; Zhang, S.; Zhang, T.; Gao, Y.; Ye, L.; Zhang, X. Elevation of highly up-regulated in liver cancer (HULC) by hepatitis B virus X protein promotes hepatoma cell proliferation via down-regulating p18. J. Biol. Chem. 2012, 287, 26302–26311. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Ma, H.; Zhou, D. Plasma HULC as a promising novel biomarker for the detection of hepatocellular carcinoma. Biomed. Res. Int. 2013, 2013, 136106. [Google Scholar] [CrossRef]
- Lu, J.; Guo, J.; Liu, J.; Mao, X.; Xu, K. Long Non-coding RNA MALAT1: A Key Player in Liver Diseases. Front. Med. 2021, 8, 734643. [Google Scholar] [CrossRef]
- Hou, Z.; Xu, X.; Zhou, L.; Fu, X.; Tao, S.; Zhou, J.; Tan, D.; Liu, S. The long non-coding RNA MALAT1 promotes the migration and invasion of hepatocellular carcinoma by sponging miR-204 and releasing SIRT1. Tumour Biol. 2017, 39, 1010428317718135. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wu, H.J.; Zhang, H.F.; Fang, S.Q.; Zeng, R. miR-143-3p inhibits proliferation and invasion of hepatocellular carcinoma cells by regulating its target gene FGF1. Clin. Transl. Oncol. 2021, 23, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Liu, J.; Huang, D. MiR-200a inhibits cell proliferation and EMT by down-regulating the ASPH expression levels and affecting ERK and PI3K/Akt pathways in human hepatoma cells. Am. J. Transl. Res. 2018, 10, 1117–1130. [Google Scholar] [PubMed]
- Cui, R.J.; Fan, J.L.; Lin, Y.C.; Pan, Y.J.; Liu, C.; Wan, J.H.; Wang, W.; Jiang, Z.Y.; Zheng, X.L.; Tang, J.B.; et al. miR-124-3p availability is antagonized by LncRNA-MALAT1 for Slug-induced tumor metastasis in hepatocellular carcinoma. Cancer Med. 2019, 8, 6358–6369. [Google Scholar] [CrossRef]
- Liu, D.; Zhu, Y.; Pang, J.; Weng, X.; Feng, X.; Guo, Y. Knockdown of long non-coding RNA MALAT1 inhibits growth and motility of human hepatoma cells via modulation of miR-195. J. Cell Biochem. 2018, 119, 1368–1380. [Google Scholar] [CrossRef]
- Chen, S.; Wang, G.; Tao, K.; Cai, K.; Wu, K.; Ye, L.; Bai, J.; Yin, Y.; Wang, J.; Shuai, X.; et al. Long noncoding RNA metastasis-associated lung adenocarcinoma transcript 1 cooperates with enhancer of zeste homolog 2 to promote hepatocellular carcinoma development by modulating the microRNA-22/Snail family transcriptional repressor 1 axis. Cancer Sci. 2020, 111, 1582–1595. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Yu, Y.; He, J.; Wang, Z.; Xu, Y.; Wang, C.; Zhang, H.; Zhang, R.; Zhang, J.; et al. LncRNA HULC enhances epithelial-mesenchymal transition to promote tumorigenesis and metastasis of hepatocellular carcinoma via the miR-200a-3p/ZEB1 signaling pathway. Oncotarget 2016, 7, 42431–42446. [Google Scholar] [CrossRef]
- Xin, X.; Wu, M.; Meng, Q.; Wang, C.; Lu, Y.; Yang, Y.; Li, X.; Zheng, Q.; Pu, H.; Gui, X.; et al. Long noncoding RNA HULC accelerates liver cancer by inhibiting PTEN via autophagy cooperation to miR15a. Mol. Cancer 2018, 17, 94. [Google Scholar] [CrossRef]
- Guo, Y.; Bai, M.; Lin, L.; Huang, J.; An, Y.; Liang, L.; Liu, Y.; Huang, W. LncRNA DLEU2 aggravates the progression of hepatocellular carcinoma through binding to EZH2. Biomed. Pharmacother. 2019, 118, 109272. [Google Scholar] [CrossRef]
- Li, B.; Li, A.; You, Z.; Xu, J.; Zhu, S. Epigenetic silencing of CDKN1A and CDKN2B by SNHG1 promotes the cell cycle, migration and epithelial-mesenchymal transition progression of hepatocellular carcinoma. Cell Death Dis. 2020, 11, 823. [Google Scholar] [CrossRef]
- Pan, J.; Hu, Y.; Yuan, C.; Wu, Y.; Zhu, X. lncRNA NEAT1 promotes the proliferation and metastasis of hepatocellular carcinoma by regulating the FOXP3/PKM2 axis. Front. Oncol. 2022, 12, 928022. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Deng, J.; Zhou, J.; Wang, Z.; Yuan, X.-F.; Pan, S.; Zhang, H.-b. Long non-coding RNA TUG1 promotes cell progression in hepatocellular carcinoma via regulating miR-216b-5p/DLX2 axis. Cancer Cell Int. 2020, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ke, J.; Guo, Q.; Barnabo Nampoukime, K.P.; Yang, P.; Ma, K. Long non-coding RNA CRNDE promotes the proliferation, migration and invasion of hepatocellular carcinoma cells through miR-217/MAPK1 axis. J. Cell Mol. Med. 2018, 22, 5862–5876. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, R.; Niu, Y.; Mo, H.; Wang, H.; Lu, Y.; Wang, L.; Sun, L.; Wang, Y.; Tu, K.; et al. HIF-1alpha-activated long non-coding RNA KDM4A-AS1 promotes hepatocellular carcinoma progression via the miR-411-5p/KPNA2/AKT pathway. Cell Death Dis. 2021, 12, 1152. [Google Scholar] [CrossRef]
- Wang, H.Z.; Liu, L.; Xu, Y.; Zhang, G.Y.; Wang, Y.Y. LncRNA UCA1 Affects the Cell Proliferation, Migration, Invasion and Apoptosis of Hepatic Carcinoma Cells by Targeting MicroRNA-193a-3p. Cancer Manag. Res. 2020, 12, 10897–10907. [Google Scholar] [CrossRef]
- Ma, J.; Li, T.; Han, X.; Yuan, H. Knockdown of LncRNA ANRIL suppresses cell proliferation, metastasis, and invasion via regulating miR-122-5p expression in hepatocellular carcinoma. J. Cancer Res. Clin. Oncol. 2018, 144, 205–214. [Google Scholar] [CrossRef]
- Huang, D.; Bi, C.; Zhao, Q.; Ding, X.; Bian, C.; Wang, H.; Wang, T.; Liu, H. Knockdown long non-coding RNA ANRIL inhibits proliferation, migration and invasion of HepG2 cells by down-regulation of miR-191. BMC Cancer 2018, 18, 919. [Google Scholar] [CrossRef]
- Li, Y.; Chen, G.; Yan, Y.; Fan, Q. CASC15 promotes epithelial to mesenchymal transition and facilitates malignancy of hepatocellular carcinoma cells by increasing TWIST1 gene expression via miR-33a-5p sponging. Eur. J. Pharmacol. 2019, 860, 172589. [Google Scholar] [CrossRef]
- Duan, R.; Li, C.; Wang, F.; Han, F.; Zhu, L. The Long Noncoding RNA ZFAS1 Potentiates the Development of Hepatocellular Carcinoma via the microRNA-624/MDK/ERK/JNK/P38 Signaling Pathway. Onco Targets Ther. 2020, 13, 4431–4444. [Google Scholar] [CrossRef]
- Dou, C.; Sun, L.; Jin, X.; Han, M.; Zhang, B.; Jiang, X.; LV, J.; Li, T. Long non-coding RNA CARLo-5 promotes tumor progression in hepatocellular carcinoma via suppressing miR-200b expression. Oncotarget 2017, 8, 70172–70182. [Google Scholar] [CrossRef]
- Xu, J.H.; Chang, W.H.; Fu, H.W.; Shu, W.Q.; Yuan, T.; Chen, P. Upregulated long non-coding RNA LOC90784 promotes cell proliferation and invasion and is associated with poor clinical features in HCC. Biochem. Biophys. Res. Commun. 2017, 490, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Han, T.; Liu, K.; Lei, C.; Wnag, Z.; Shi, G. LncRNA H19 promotes the development of hepatitis B related hepatocellular carcinoma through regulating microRNA-22 via EMT pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5392–5401. [Google Scholar] [PubMed]
- LV, J.; Yu, Y.; Li, S.; Luo, L.; Wang, Q. Aflatoxin B1 Promotes Cell Growth and Invasion in Hepatocellular Carcinoma HepG2 Cells through H19 and E2F1. Asian Pac. J. Cancer Prev. 2014, 15, 2565–2570. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, J.; Chen, W.; Fan, J.; Hylemon, P.B.; Zhou, H. Long Noncoding RNA H19: A Novel Oncogene in Liver Cancer. Noncoding RNA 2023, 9, 19. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Wu, G.; Yang, Y.; Tang, Y.; Zhang, W.; Wang, K.; Liu, Y.; Wang, X.; Li, T. Long noncoding RNA PCAT-14 induces proliferation and invasion by hepatocellular carcinoma cells by inducing methylation of miR-372. Oncotarget 2017, 8, 34429–34441. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Wang, W.; Liu, F.; Wang, W.; Su, C.; Zhu, H.; Liao, Z.; Zhang, B.; Chen, X. ALKBH5-mediated m6A modification of lincRNA LINC02551 enhances the stability of DDX24 to promote hepatocellular carcinoma growth and metastasis. Cell Death Dis. 2022, 13, 926. [Google Scholar] [CrossRef]
- Tao, H.; Zhang, Y.; Yan, T.; Li, J.; Liu, J.; Xiong, Y.; Zhu, J.; Huang, Z.; Wang, P.; Liang, H.; et al. Identification of an EMT-related lncRNA signature and LINC01116 as an immune-related oncogene in hepatocellular carcinoma. Aging 2022, 14, 1473–1491. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, D.; Li, Y.; Yan, S.; Dang, H.; Yue, H.; Ling, J.; Chen, F.; Zhao, Y.; Gou, L.; et al. Long noncoding RNA CCAT2 promotes hepatocellular carcinoma proliferation and metastasis through up-regulation of NDRG1. Exp. Cell Res. 2019, 379, 19–29. [Google Scholar] [CrossRef]
- Su, R.; Zhang, H.; Zhang, L.; Khan, A.R.; Zhang, X.; Wang, R.; Shao, C.; Wei, X.; Xu, X. Systemic analysis identifying PVT1/DUSP13 axis for microvascular invasion in hepatocellular carcinoma. Cancer Med. 2023, 12, 8937–8955. [Google Scholar] [CrossRef]
- Luo, Z.; Cao, P. Long noncoding RNA PVT1 promotes hepatoblastoma cell proliferation through activating STAT3. Cancer Manag. Res. 2019, 11, 8517–8527. [Google Scholar] [CrossRef]
- Wang, X.L.; Shi, M.; Xiang, T.; Bu, Y.Z. Long noncoding RNA DGCR5 represses hepatocellular carcinoma progression by inactivating Wnt signaling pathway. J. Cell Biochem. 2019, 120, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, F.; Qu, L.; Wang, Z.; Liu, X. MEG3 promotes liver cancer by activating PI3K/AKT pathway through regulating AP1G1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1459–1467. [Google Scholar]
- Wei, Q.; Liu, G.; Huang, Z.; Huang, Y.; Huang, L.; Huang, Z.; Wu, X.; Wei, H.; Pu, J. LncRNA MEG3 Inhibits Tumor Progression by Modulating Macrophage Phenotypic Polarization via miR-145-5p/DAB2 Axis in Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2023, 10, 1019–1035. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Pang, R.; Li, M.; Chen, B.; Huang, J.; Zhu, Y. m6A-Induced LncRNA MEG3 Suppresses the Proliferation, Migration and Invasion of Hepatocellular Carcinoma Cell Through miR-544b/BTG2 Signaling. Onco Targets Ther. 2021, 14, 3745–3755. [Google Scholar] [CrossRef]
- Qian, G.; Jin, X.; Zhang, L. LncRNA FENDRR Upregulation Promotes Hepatic Carcinoma Cells Apoptosis by Targeting miR-362-5p Via NPR3 and p38-MAPK Pathway. Cancer Biother. Radiopharm. 2020, 35, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhao, H.; Feng, X.; Li, H.; Qiu, C.; Yi, X.; Tang, H.; Zhang, J. Long Non-coding RNA FENDRR Acts as a miR-423-5p Sponge to Suppress the Treg-Mediated Immune Escape of Hepatocellular Carcinoma Cells. Mol. Ther. Nucleic Acids 2019, 17, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ke, S.; Li, M.; Lin, C.; Liu, X.; Pan, Q. Downregulation of LncRNA GAS5 promotes liver cancer proliferation and drug resistance by decreasing PTEN expression. Mol. Genet. Genom. 2020, 295, 251–260. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.Y.; Zhao, W.; Gao, Z.K.; Shen, B.; Xu, H.; Cui, Y.F. Long non-coding RNA GAS5 overexpression inhibits M2-like polarization of tumour-associated macrophages in SMCC-7721 cells by promoting PTEN expression. Int. J. Exp. Pathol. 2020, 101, 215–222. [Google Scholar] [CrossRef]
- Dai, Y.Z.; Liu, Y.D.; Li, J.; Chen, M.T.; Huang, M.; Wang, F.; Yang, Q.S.; Yuan, J.H.; Sun, S.H. METTL16 promotes hepatocellular carcinoma progression through downregulating RAB11B-AS1 in an m6A-dependent manner. Cell Mol. Biol. Lett. 2022, 27, 41. [Google Scholar] [CrossRef]
- Park, E.G.; Pyo, S.J.; Cui, Y.; Yoon, S.H.; Nam, J.W. Tumor immune microenvironment lncRNAs. Brief. Bioinform. 2022, 23, bbab504. [Google Scholar] [CrossRef]
- Jiang, P.; Li, X. Regulatory Mechanism of lncRNAs in M1/M2 Macrophages Polarization in the Diseases of Different Etiology. Front. Immunol. 2022, 13, 835932. [Google Scholar] [CrossRef]
- Hou, Z.H.; Xu, X.W.; Fu, X.Y.; Zhou, L.D.; Liu, S.P.; Tan, D.M. Long non-coding RNA MALAT1 promotes angiogenesis and immunosuppressive properties of HCC cells by sponging miR-140. Am. J. Physiol. Cell Physiol. 2020, 318, C649–C663. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Tang, J.; Chen, Y.; Deng, L.; Ji, J.; Xie, Y.; Wang, K.; Jia, W.; Chu, W.M.; Sun, B. The long noncoding RNA lnc-EGFR stimulates T-regulatory cells differentiation thus promoting hepatocellular carcinoma immune evasion. Nat. Commun. 2017, 8, 15129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, L.; Dong, X.; Yang, X.; Zhang, X.; Liu, Z.; Zhao, X.; Wen, T. Overexpression of NNT-AS1 Activates TGF-beta Signaling to Decrease Tumor CD4 Lymphocyte Infiltration in Hepatocellular Carcinoma. Biomed. Res. Int. 2020, 2020, 8216541. [Google Scholar] [CrossRef]
- Peng, L.; Chen, Y.; Ou, Q.; Wang, X.; Tang, N. LncRNA MIAT correlates with immune infiltrates and drug reactions in hepatocellular carcinoma. Int. Immunopharmacol. 2020, 89, 107071. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Nie, P.; Peng, D.; He, Z.; Liu, M.; Xie, Y.; Miao, Y.; Zuo, Z.; Ren, J. m6AVar: A database of functional variants involved in m6A modification. Nucleic Acids Res. 2018, 46, D139–D145. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, H.; Liang, J.; Zhao, Q.; Xie, Y.; Ren, J.; Zuo, Z. RMVar: An updated database of functional variants involved in RNA modifications. Nucleic Acids Res. 2021, 49, D1405–D1412. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m6A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef]
- Schwartz, S.; Mumbach, M.R.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D.; et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5′ sites. Cell Rep. 2014, 8, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m6A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Xu, C.; Liu, K.; Ahmed, H.; Loppnau, P.; Schapira, M.; Min, J. Structural Basis for the Discriminative Recognition of N6-Methyladenosine RNA by the Human YT521-B Homology Domain Family of Proteins. J. Biol. Chem. 2015, 290, 24902–24913. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Zuo, X.; Chen, Z.; Gao, W.; Zhang, Y.; Wang, J.; Wang, J.; Cao, M.; Cai, J.; Wu, J.; Wang, X. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J. Hematol. Oncol. 2020, 13, 5. [Google Scholar] [CrossRef]
- Li, L.; Xie, R.; Lu, G. Identification of m6A methyltransferase-related lncRNA signature for predicting immunotherapy and prognosis in patients with hepatocellular carcinoma. Biosci. Rep. 2021, 41, BSR20210760. [Google Scholar] [CrossRef]
- Peng, L.; Pan, B.; Zhang, X.; Wang, Z.; Qiu, J.; Wang, X.; Tang, N. Lipopolysaccharide facilitates immune escape of hepatocellular carcinoma cells via m6A modification of lncRNA MIR155HG to upregulate PD-L1 expression. Cell Biol. Toxicol. 2022, 38, 1159–1173. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, N.; Zeng, J.; Wang, T.; Shen, Y.; Ma, C.; Yang, M. N6-methyladenosine-modified lncRNA ARHGAP5-AS1 stabilises CSDE1 and coordinates oncogenic RNA regulons in hepatocellular carcinoma. Clin. Transl. Med. 2022, 12, e1107. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Wang, M.; Wang, G.; Mai, Z.; Zhou, B.; Han, Y.; Zhuang, J.; Xia, W. lncRNA miR4458HG modulates hepatocellular carcinoma progression by activating m6A-dependent glycolysis and promoting the polarization of tumor-associated macrophages. Cell Mol. Life Sci. 2023, 80, 99. [Google Scholar] [CrossRef] [PubMed]
- Hammerle, M.; Gutschner, T.; Uckelmann, H.; Ozgur, S.; Fiskin, E.; Gross, M.; Skawran, B.; Geffers, R.; Longerich, T.; Breuhahn, K.; et al. Posttranscriptional destabilization of the liver-specific long noncoding RNA HULC by the IGF2 mRNA-binding protein 1 (IGF2BP1). Hepatology 2013, 58, 1703–1712. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Lin, J.; Xie, X.; Xie, Y.; Zhang, J.; Xu, D.; He, X.; Liu, F.; Xie, B. LncRNA SLC7A11-AS1 promotes the progression of hepatocellular carcinoma by mediating KLF9 ubiquitination. Neoplasma 2023, 70, 361–374. [Google Scholar] [CrossRef]
- Zhang, Q.; Wei, T.; Yan, L.; Zhu, S.; Jin, W.; Bai, Y.; Zeng, Y.; Zhang, X.; Yin, Z.; Yang, J.; et al. Hypoxia-Responsive lncRNA AC115619 Encodes a Micropeptide That Suppresses m6A Modifications and Hepatocellular Carcinoma Progression. Cancer Res. 2023, 83, 2496–2512. [Google Scholar] [CrossRef]
- Beylerli, O.; Gareev, I.; Sufianov, A.; Ilyasova, T.; Guang, Y. Long noncoding RNAs as promising biomarkers in cancer. Noncoding RNA Res. 2022, 7, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Taketa, K.; Okada, S.; Win, N.; Hlaing, N.; Win, K.-M. Evaluation of Tumor Markers for the Detection of Hepatocellular Carcinoma in Yangon General Hospital, Myanmar. Acta Med. Okayama. 2002, 56, 317–320. [Google Scholar]
- Khien, V.-V.; Mao, H.; Chinh, T.; Bang, M.; Lac, B.-V.; Hop, T.-V.; Tuan, N.-A.; Don, L.; Taketa, K.; Satomura, S. Clinical evaluation of lentil lectin-reactive alpha-fetoprotein-L3 in histology-proven hepatocellular carcinoma. Int. J. Biol. Markers 2001, 16, 105–111. [Google Scholar] [CrossRef]
- Hippo, Y.; Watanabe, K.; Watanabe, A.; Midorikawa, Y.; Yamamoto, S.; Ihara, S.; Tokita, S.; Iwanari, H.; Ito, Y.; Nakano, K.; et al. Identification of Soluble NH2-Terminal Fragment of Glypican-3 as a Serological Marker for Early-Stage Hepatocellular Carcinoma. Cancer Res. 2004, 64, 2418–2423. [Google Scholar] [CrossRef]
- Miura, N.; Maeda, Y.; Kanbe, Y.; Yazama, H.; Takeda, Y.; Sato, R.; Tsukamoto, T.; Sato, E.; Marunoto, A.; Harada, T.; et al. Serum HumanTelomerase ReverseTranscriptase Messenger RNA as a NovelTumor Marker for Hepatocellular Carcinoma. Clin. Cancer Res. 2005, 11, 3205–3209. [Google Scholar] [CrossRef]
- Kim, H.S.; Yoon, J.H.; Baek, G.O.; Yoon, M.G.; Han, J.E.; Cho, H.J.; Kim, S.S.; Jeong, J.Y.; Cheong, J.Y.; Eun, J.W. Tumor Endothelial Cells-Associated Integrin Alpha-6 as a Promising Biomarker for Early Detection and Prognosis of Hepatocellular Carcinoma. Cancers 2023, 15, 4156. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Eun, J.W.; Lee, K.; Kim, H.S.; Yang, H.D.; Kim, S.Y.; Lee, E.K.; Kim, T.; Kang, K.; Kim, S.; et al. Barrier to autointegration factor 1, procollagen-lysine, 2-oxoglutarate 5-dioxygenase 3, and splicing factor 3b subunit 4 as early-stage cancer decision markers and drivers of hepatocellular carcinoma. Hepatology 2018, 67, 1360–1377. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zheng, Y.; Xiao, X.; Liu, C.; Lin, J.; Zheng, S.; Yang, B.; Ou, Q. A Circulating Long Noncoding RNA Panel Serves as a Diagnostic Marker for Hepatocellular Carcinoma. Dis. Markers 2020, 2020, 5417598. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Baek, G.O.; Ahn, H.R.; Sung, S.; Seo, C.W.; Cho, H.J.; Nam, S.W.; Cheong, J.Y.; Eun, J.W. Serum small extracellular vesicle-derived LINC00853 as a novel diagnostic marker for early hepatocellular carcinoma. Mol. Oncol. 2020, 14, 2646–2659. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Baek, G.O.; Son, J.A.; Ahn, H.R.; Yoon, M.K.; Cho, H.J.; Yoon, J.H.; Nam, S.W.; Cheong, J.Y.; Eun, J.W. Early detection of hepatocellular carcinoma via liquid biopsy: Panel of small extracellular vesicle-derived long noncoding RNAs identified as markers. Mol. Oncol. 2021, 15, 2715–2731. [Google Scholar] [CrossRef]
- Yuan, S.X.; Yang, F.; Yang, Y.; Tao, Q.F.; Zhang, J.; Huang, G.; Yang, Y.; Wang, R.Y.; Yang, S.; Huo, X.S.; et al. Long noncoding RNA associated with microvascular invasion in hepatocellular carcinoma promotes angiogenesis and serves as a predictor for hepatocellular carcinoma patients’ poor recurrence-free survival after hepatectomy. Hepatology 2012, 56, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Dong, J.; Li, Y.; Dong, Z.; Liu, Z.; Huang, J.; Wang, Y.; Zhen, Y.; Lu, Y. The expression level and clinical significance of lncRNA X91348 in hepatocellular carcinoma. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3067–3071. [Google Scholar] [CrossRef]
- Wu, L.; Yang, Z.; Zhang, J.; Xie, H.; Zhou, L.; Zheng, S. Long noncoding RNA HOTTIP expression predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Hepatobiliary Surg. Nutr. 2018, 7, 429–439. [Google Scholar] [CrossRef]
- Lee, Y.R.; Kim, G.; Tak, W.Y.; Jang, S.Y.; Kweon, Y.O.; Park, J.G.; Lee, H.W.; Han, Y.S.; Chun, J.M.; Park, S.Y.; et al. Circulating exosomal noncoding RNAs as prognostic biomarkers in human hepatocellular carcinoma. Int. J. Cancer 2019, 144, 1444–1452. [Google Scholar] [CrossRef]
- Lin, L.Y.; Yang, L.; Zeng, Q.; Wang, L.; Chen, M.L.; Zhao, Z.H.; Ye, G.D.; Luo, Q.C.; Lv, P.Y.; Guo, Q.W.; et al. Tumor-originated exosomal lncUEGC1 as a circulating biomarker for early-stage gastric cancer. Mol. Cancer 2018, 17, 84. [Google Scholar] [CrossRef]
- Vosough, P.; Khatami, S.H.; Hashemloo, A.; Tajbakhsh, A.; Karimi-Fard, F.; Taghvimi, S.; Taheri-Anganeh, M.; Soltani Fard, E.; Savardashtaki, A.; Movahedpour, A. Exosomal lncRNAs in gastrointestinal cancer. Clin. Chim. Acta 2023, 540, 117216. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, M.; Chen, B.; Calin, G.A. Exosomal lncRNAs as new players in cell-to-cell communication. Transl. Cancer Res. 2018, 7, S243–S252. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Shao, Y.; Zhang, X.; Zheng, T.; Miao, M.; Qin, L.; Wang, B.; Ye, G.; Xiao, B.; Guo, J. Plasma long noncoding RNA protected by exosomes as a potential stable biomarker for gastric cancer. Tumour Biol. 2015, 36, 2007–2012. [Google Scholar] [CrossRef]
- Lee, S.A.; Yoo, T.H. Therapeutic application of extracellular vesicles for various kidney diseases: A brief review. BMB Rep. 2022, 55, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Han, J.; Zhang, J.; Li, G.; Liu, H.; Cui, X.; Xu, Y.; Li, T.; Liu, J.; Wang, C. The long noncoding RNAs PVT1 and uc002mbe.2 in sera provide a new supplementary method for hepatocellular carcinoma diagnosis. Medicine 2016, 95, e4436. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.K.; Pang, C.; Yang, Y.; Duan, Q.; Zhang, J.; Liu, W.C. Serum long noncoding RNA urothelial carcinoma-associated 1: A novel biomarker for diagnosis and prognosis of hepatocellular carcinoma. J. Int. Med. Res. 2018, 46, 348–356. [Google Scholar] [CrossRef]
- Abdelmoety, A.A.; Elhassafy, M.Y.; Omar Said, R.S.; Elsheaita, A.; Mahmoud, M.M. The role of UCA1 and WRAP53 in diagnosis of hepatocellular carcinoma: A single-center case-control study. Clin. Exp. Hepatol. 2023, 9, 129–137. [Google Scholar] [CrossRef]
- Yuan, W.; Sun, Y.; Liu, L.; Zhou, B.; Wang, S.; Gu, D. Circulating LncRNAs Serve as Diagnostic Markers for Hepatocellular Carcinoma. Cell Physiol. Biochem. 2017, 44, 125–132. [Google Scholar] [CrossRef]
- Yin, Q.; Huang, X.; Yang, Q.; Lin, S.; Song, Q.; Fan, W.; Li, W.; Li, Z.; Gao, L. LncRNA model predicts liver cancer drug resistance and validate in vitro experiments. Front. Cell Dev. Biol. 2023, 11, 1174183. [Google Scholar] [CrossRef]
- Guo, D.; Li, Y.; Chen, Y.; Zhang, D.; Wang, X.; Lu, G.; Ren, M.; Lu, X.; He, S. DANCR promotes HCC progression and regulates EMT by sponging miR-27a-3p via ROCK1/LIMK1/COFILIN1 pathway. Cell Prolif. 2019, 52, e12628. [Google Scholar] [CrossRef]
- Xu, X.; Gu, J.; Ding, X.; Ge, G.; Zang, X.; Ji, R.; Shao, M.; Mao, Z.; Zhang, Y.; Zhang, J.; et al. LINC00978 promotes the progression of hepatocellular carcinoma by regulating EZH2-mediated silencing of p21 and E-cadherin expression. Cell Death Dis. 2019, 10, 752. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Lin, Z.; Ye, H.; Cheng, D.; Zhang, G.; Zhou, J.; Huang, Z.; Wang, M.; Cai, C.; Zeng, J.; et al. Upregulation of LncDQ is Associated with Poor Prognosis and Promotes Tumor Progression via Epigenetic Regulation of the EMT Pathway in HCC. Cell Physiol. Biochem. 2018, 46, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Chen, X.; Wu, X.; Li, J.; Mei, C.; Jing, W.; Teng, L.; Tu, H.; Jiang, X.; Wang, G.; et al. Long noncoding RNA SPRY4-IT1 promotes proliferation and metastasis of hepatocellular carcinoma via mediating TNF signaling pathway. J. Cell Physiol. 2020, 235, 7849–7862. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Tang, J.; Huang, R.; Li, J.; Xu, D.; Xie, Y.; Jiang, R.; Deng, L.; Zhang, X.; et al. LINC01225 promotes occurrence and metastasis of hepatocellular carcinoma in an epidermal growth factor receptor-dependent pathway. Cell Death Dis. 2016, 7, e2130. [Google Scholar] [CrossRef]
- Wang, D.; Xing, N.; Yang, T.; Liu, J.; Zhao, H.; He, J.; Ai, Y.; Yang, J. Exosomal lncRNA H19 promotes the progression of hepatocellular carcinoma treated with Propofol via miR-520a-3p/LIMK1 axis. Cancer Med. 2020, 9, 7218–7230. [Google Scholar] [CrossRef] [PubMed]
- Motawi, T.M.K.; El-Maraghy, S.A.; Sabry, D.; Mehana, N.A. The expression of long non coding RNA genes is associated with expression with polymorphisms of HULC rs7763881 and MALAT1 rs619586 in hepatocellular carcinoma and HBV Egyptian patients. J. Cell Biochem. 2019, 120, 14645–14656. [Google Scholar] [CrossRef]
- Yao, Z.; Jia, C.; Tai, Y.; Liang, H.; Zhong, Z.; Xiong, Z.; Deng, M.; Zhang, Q. Serum exosomal long noncoding RNAs lnc-FAM72D-3 and lnc-EPC1-4 as diagnostic biomarkers for hepatocellular carcinoma. Aging 2020, 12, 11843–11863. [Google Scholar] [CrossRef]
- Lin, X.Q.; Huang, Z.M.; Chen, X.; Wu, F.; Wu, W. XIST Induced by JPX Suppresses Hepatocellular Carcinoma by Sponging miR-155-5p. Yonsei Med. J. 2018, 59, 816–826. [Google Scholar] [CrossRef]
- Wang, Y.G.; Liu, J.; Shi, M.; Chen, F.X. LncRNA DGCR5 represses the development of hepatocellular carcinoma by targeting the miR-346/KLF14 axis. J. Cell Physiol. 2018, 234, 572–580. [Google Scholar] [CrossRef]
- Pote, N.; Cauchy, F.; Albuquerque, M.; Voitot, H.; Belghiti, J.; Castera, L.; Puy, H.; Bedossa, P.; Paradis, V. Performance of PIVKA-II for early hepatocellular carcinoma diagnosis and prediction of microvascular invasion. J. Hepatol. 2015, 62, 848–854. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.D. Non-coding RNA: A new frontier in regulatory biology. Natl. Sci. Rev. 2014, 1, 190–204. [Google Scholar] [CrossRef] [PubMed]
- The, R.C.; Petrov, A.I.; Kay, S.J.E.; Kalvari, I.; Howe, K.L.; Gray, K.A.; Bruford, E.A.; Kersey, P.J.; Cochrane, G.; Finn, R.D.; et al. RNAcentral: A comprehensive database of non-coding RNA sequences. Nucleic Acids Res. 2017, 45, D128–D134. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Tastsoglou, S.; Skoufos, G.; Karavangeli, A.; Pierros, V.; Zacharopoulou, E.; Hatzigeorgiou, A.G. DIANA-LncBase v3: Indexing experimentally supported miRNA targets on non-coding transcripts. Nucleic Acids Res. 2020, 48, D101–D110. [Google Scholar] [CrossRef] [PubMed]
- Hofacker, I.L.; Stadler, P.F. Memory efficient folding algorithms for circular RNA secondary structures. Bioinformatics 2006, 22, 1172–1176. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, J.; Gao, Y.; Li, Y.; Feng, C.; Song, C.; Ning, Z.; Zhou, X.; Zhao, J.; Feng, M.; et al. LncSEA: A platform for long non-coding RNA related sets and enrichment analysis. Nucleic Acids Res. 2021, 49, D969–D980. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.; Jiang, S.; Li, Q.; Feng, C.; Du, Q.; Zou, D.; Xiao, J.; Zhang, Z.; Ma, L. LncExpDB: An expression database of human long non-coding RNAs. Nucleic Acids Res. 2021, 49, D962–D968. [Google Scholar] [CrossRef]
- Seifuddin, F.; Singh, K.; Suresh, A.; Judy, J.T.; Chen, Y.C.; Chaitankar, V.; Tunc, I.; Ruan, X.; Li, P.; Chen, Y.; et al. lncRNAKB, a knowledgebase of tissue-specific functional annotation and trait association of long noncoding RNA. Sci. Data 2020, 7, 326. [Google Scholar] [CrossRef]
- Volders, P.J.; Helsens, K.; Wang, X.; Menten, B.; Martens, L.; Gevaert, K.; Vandesompele, J.; Mestdagh, P. LNCipedia: A database for annotated human lncRNA transcript sequences and structures. Nucleic Acids Res. 2013, 41, D246–D251. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhang, W.Y.; Xin, X.H.; Du, P.F. dbEssLnc: A manually curated database of human and mouse essential lncRNA genes. Comput. Struct. Biotechnol. J. 2022, 20, 2657–2663. [Google Scholar] [CrossRef]
- Li, J.; Ma, W.; Zeng, P.; Wang, J.; Geng, B.; Yang, J.; Cui, Q. LncTar: A tool for predicting the RNA targets of long noncoding RNAs. Brief. Bioinform. 2015, 16, 806–812. [Google Scholar] [CrossRef]
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An Interactive Open Platform to Explore the Function of lncRNAs in Cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, L.; Feng, C.; Qin, Y.; Xiao, J.; Zhang, Z.; Ma, L. LncBook 2.0: Integrating human long non-coding RNAs with multi-omics annotations. Nucleic Acids Res. 2023, 51, D186–D191. [Google Scholar] [CrossRef]
- Mas-Ponte, D.; Carlevaro-Fita, J.; Palumbo, E.; Hermoso Pulido, T.; Guigo, R.; Johnson, R. LncATLAS database for subcellular localization of long noncoding RNAs. RNA 2017, 23, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Tang, Q.; He, J.; Li, L.; Yang, N.; Yu, S.; Wang, M.; Zhang, Y.; Lin, J.; Cui, T.; et al. RNAInter v4.0: RNA interactome repository with redefined confidence scoring system and improved accessibility. Nucleic Acids Res. 2022, 50, D326–D332. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef]
- Cui, L.; Ma, R.; Cai, J.; Guo, C.; Chen, Z.; Yao, L.; Wang, Y.; Fan, R.; Wang, X.; Shi, Y. RNA modifications: Importance in immune cell biology and related diseases. Signal Transduct. Target. Ther. 2022, 7, 334. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Expression | lncRNA | Known Function | Reference |
|---|---|---|---|
| Upregulated | HULC | EMT, metastasis, apoptosis | [56,67,68] |
| MALAT1 | EMT, metastasis, apoptosis, cell-cycle arrest | [61,65] | |
| HOTAIR | EMT, metastasis, apoptosis, cell-cycle arrest | [34,35,36] | |
| DLEU2 | Vascular invasion, lymphatic metastasis | [69] | |
| SNHG1 | EMT, cell-cycle regulation, metastasis, apoptosis | [70] | |
| NEAT1 | Ferroptosis, metastasis, proliferation, invasion, drug resistance | [47,51,53,54,71] | |
| TUG1 | Metastasis, apoptosis | [72] | |
| CRNDE | Proliferation, migration, invasion | [73] | |
| KDM4A-AS1 | EMT, metastasis | [74] | |
| UCA1 | EMT, cell-cycle regulation, apoptosis | [75] | |
| ANRIL | Metastasis, apoptosis | [76,77] | |
| CASC15 | EMT, metastasis, suppression of apoptosis | [78] | |
| ZFAS1 | Metastasis | [79] | |
| CARLo-5 | EMT | [80] | |
| LOC90784 | Apoptosis, cell-cycle arrest | [81] | |
| H19 | EMT, metastasis, suppression of apoptosis | [82,83,84] | |
| PCAT-14 | cell-cycle arrest | [85] | |
| LINC02551 | EMT, metastasis | [86] | |
| LINC01116 | EMT, cell-cycle regulation, metastasis, immune-cell infiltration | [87] | |
| CCAT2 | Proliferation, migration, invasion | [88] | |
| PVT1 | Microvascular invasion, proliferation | [89,90] | |
| Downregulated | DGCR5 | Proliferation, migration, invasion | [91] |
| MEG3 | Inhibition of metastasis, angioinvasion and proliferation by cell-cycle regulation | [92,93,94] | |
| FENDRR | Apoptosis, Treg-mediated immune escape | [95,96] | |
| GAS5 | Suppression of proliferation, drug resistance and M2 macrophage polarization | [97,98] | |
| RAB11B-AS1 | Apoptosis | [99] |
| Expression | lncRNA | m6A Binding Partner | Reference |
|---|---|---|---|
| Upregulated | HULC | IGF2BP1 | [123] |
| LINC00958 | METTL3 | [118] | |
| LINC02362 | - | [119] | |
| SNHG20 | |||
| SNHG6 | |||
| MIR4458HG | IGF2BP2 | [122] | |
| LINC02551 | ALKBH5 | [86] | |
| MIR155HG | METTL14 | [120] | |
| SLC7A11-AS1 | METTL3 | [124] | |
| ARHGAP5-AS1 | METTL14 IGF2BP2 | [121] | |
| Downregulated | RAB11B-AS1 | METTL16 | [99] |
| MEG3 | METTL3 | [94] | |
| AC115619 | WTAP | [125] |
| Expression | lncRNA | Blood Detection | Reference |
|---|---|---|---|
| Upregulated | MVIH | Yes | [136] |
| HOTTIP | Yes | [135,138] | |
| ATB | Yes | [139] | |
| DLEU2 | Yes | [135] | |
| MALAT1 | Yes | ||
| SNHG1 | Yes | ||
| PVT1 | Yes | [145] | |
| uc002mbe.2 | Yes | ||
| LINC00853 | Yes | [134] | |
| UCA1 | Yes | [146,147] | |
| WRAP53 | Yes | [147] | |
| LINC00152 | Yes | [148] | |
| RP11-160H22.5 | Yes | ||
| XLOC014172 | Yes | ||
| DANCR | - | [150] | |
| LINC00978 | Yes | [151] | |
| LncDQ | Yes | [152] | |
| SPRY4-IT1 | - | [153] | |
| UBE2CP3 | Yes | [133] | |
| LINC01225 | Yes | [154] | |
| H19 | Yes | [155] | |
| uc003wbd | Yes | [156] | |
| CAHM | - | [149] | |
| FAM72D-3 | Yes | [157] | |
| EPC1-4 | Yes | ||
| Downregulated | JPX | - | [158] |
| XIST | - | ||
| DGCR5 | - | [159] | |
| X91348 | Yes | [137] |
| Name | Website | Description | Reference |
|---|---|---|---|
| RNACentral | https://rnacentral.org (accessed on 10 September 2023) | A public platform offering access to a wide collection of non-coding RNA sequences from various organisms and RNA types | [163] |
| LncBase | https://diana.e-ce.uth.gr/lncbasev3 (accessed on 10 September 2023) | A database cataloging 500,000 verified miRNA–lncRNA interactions across 243 cell types | [164] |
| RNAfold | http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi (accessed on 10 September 2023) | A tool used for predicting the secondary structure of RNA sequences | [165] |
| LncSEA | http://bio.liclab.net/LncSEA/index.php (accessed on 10 September 2023) | A platform for lncRNA-related sets and enrichment analysis | [166] |
| LncExpDB | https://ngdc.cncb.ac.cn/lncexpdb (accessed on 10 September 2023) | Expression database of human lncRNAs | [167] |
| lncRNAKB | https://bio.tools/lncrnakb (accessed on 10 September 2023) | A knowledgebase of tissue-specific functional annotation and trait association of lncRNA | [168] |
| LNCipedia | http://www.lncipedia.org (accessed on 10 September 2023) | A database for annotated human lncRNA transcript sequences and structures | [169] |
| dbEssLnc | https://esslnc.pufengdu.org (accessed on 10 September 2023) | A manually curated database of human and mouse essential lncRNA genes | [170] |
| LncTar | http://www.cuilab.cn/lnctar (accessed on 10 September 2023) | A tool for predicting the RNA targets of lncRNAs. | [171] |
| TANRIC | https://bioinformatics.mdanderson.org/public-software/tanric (accessed on 10 September 2023) | TANRIC webapp provides analysis of lncRNA in cancer, highlighting potential therapeutic targets and biomarkers | [172] |
| LncBook | https://ngdc.cncb.ac.cn/lncbook (accessed on 10 September 2023) | A comprehensive database of human lncRNAs, offering annotations for understanding their roles in diseases and biological contexts. | [173] |
| lncATLAS | https://lncatlas.crg.eu (accessed on 10 September 2023) | A database showing subcellular locations of GENCODE-annotated lncRNAs, using RCI values | [174] |
| RNAInter | http://www.rnainter.org (accessed on 10 September 2023) | A database with a scoring system to rate the confidence of RNA-associated interactions based on experimental evidence and tissue/cell types | [175] |
| ENCORI | https://rnasysu.com/encori (accessed on 10 September 2023) | A platform for studying RNA interactions, integrating diverse data and enabling pan-cancer analysis | [176] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eun, J.W.; Cheong, J.Y.; Jeong, J.-Y.; Kim, H.S. A New Understanding of Long Non-Coding RNA in Hepatocellular Carcinoma—From m6A Modification to Blood Biomarkers. Cells 2023, 12, 2272. https://doi.org/10.3390/cells12182272
Eun JW, Cheong JY, Jeong J-Y, Kim HS. A New Understanding of Long Non-Coding RNA in Hepatocellular Carcinoma—From m6A Modification to Blood Biomarkers. Cells. 2023; 12(18):2272. https://doi.org/10.3390/cells12182272
Chicago/Turabian StyleEun, Jung Woo, Jae Youn Cheong, Jee-Yeong Jeong, and Hyung Seok Kim. 2023. "A New Understanding of Long Non-Coding RNA in Hepatocellular Carcinoma—From m6A Modification to Blood Biomarkers" Cells 12, no. 18: 2272. https://doi.org/10.3390/cells12182272