
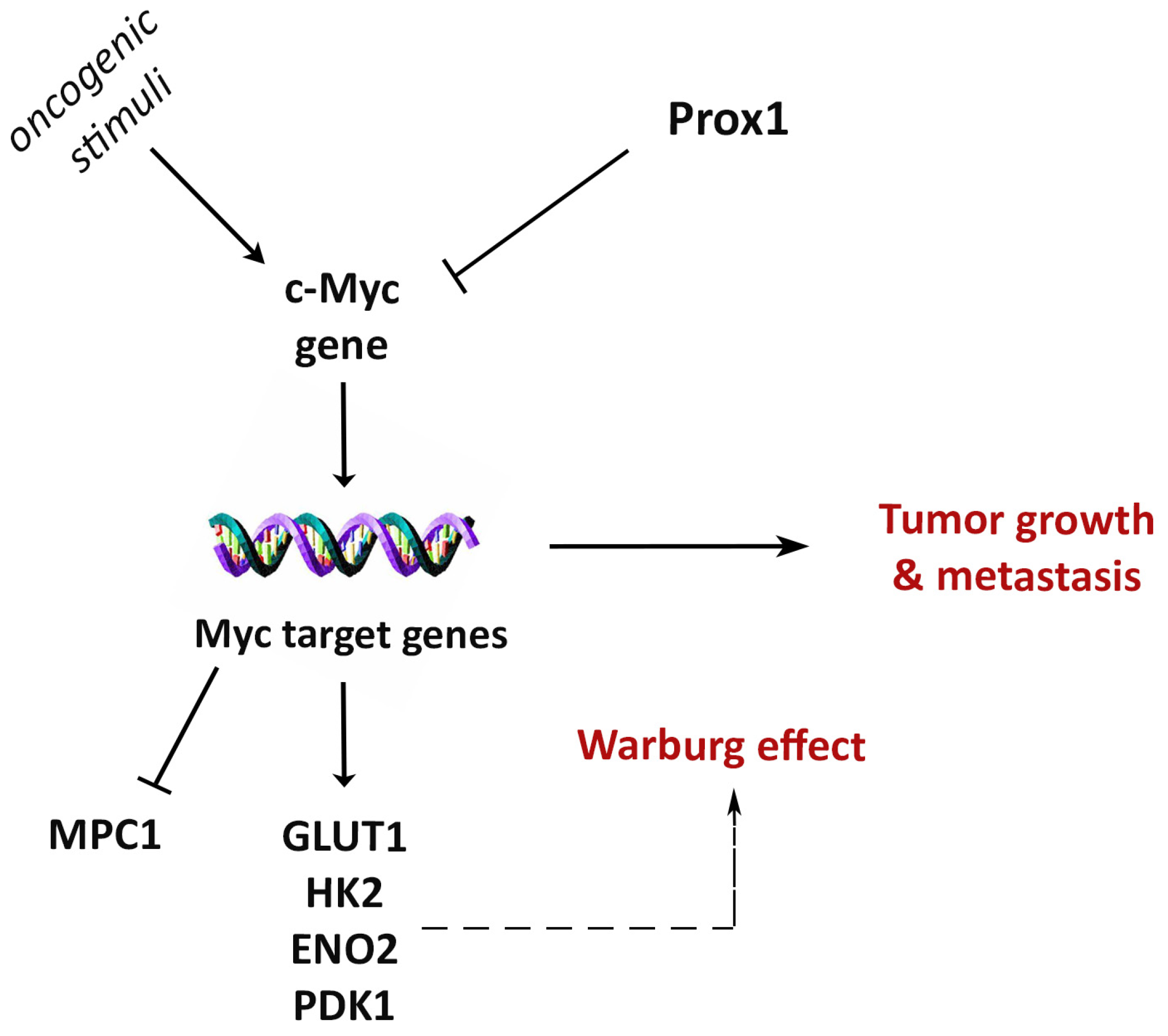
Prox1 Suppresses the Proliferation of Breast Cancer Cells via Direct Inhibition of c-Myc Gene Expression
 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Reagents
2.3. Biological Resources
2.4. Transfection and Infection Methods
2.5. Transwell Chamber Assay
2.6. RNA Extraction and Real Time RT-qPCR Analysis
2.7. Western Blot Analysis
2.8. Fluorescence-Activated Cell Sorting (FACS) Analysis
2.9. Immunostainings
2.10. Allotransplantation Experiments (Xenografts)
2.10.1. Orthotopic Xenografts for Breast Cancer
2.10.2. Heterotopic Xenografts for Breast Cancer
2.11. Wound Healing Assay
2.12. Glucose and Lactate Assays
2.13. Cross-Linking Chromatin Immunoprecipitation
2.14. Statistical Analysis and Experimental Design
2.15. Web Sites/Data Base Referencing
3. Results
3.1. Prox1 Is Significantly Reduced in Breast Cancer and Its Expression Is Correlated to Favorable Prognosis
3.2. Prox1 Inhibits Breast Cancer Cell Proliferation without inducing Cell Death
3.3. Prox1 Over-Expression Represses Breast Cancer Cells’ Capacity for Migration and Invasion
3.4. Prox1 Negatively Affects Glucose Uptake and Lactate Secretion by Breast Cancer Cells
3.5. Prox1 Inhibits Expression of Genes That Promote the Warburg Effect in Breast Cancer Cells
3.6. Prox1 Directly Suppresses c-Myc Gene Expression
3.7. c-Myc Over-Expression Rescues the Anti-Proliferative Effect of Prox1 on Breast Tumor Cells
3.8. Prox1 Suppresses Tumor Growth In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wigle, J.T.; Chowdhury, K.; Gruss, P.; Oliver, G. Prox1 function is crucial for mouse lens-fibre elongation. Nat. Genet. 1999, 21, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Risebro, C.A.; Searles, R.G.; Melville, A.A.; Ehler, E.; Jina, N.; Shah, S.; Pallas, J.; Hubank, M.; Dillard, M.; Harvey, N.L.; et al. Prox1 maintains muscle structure and growth in the developing heart. Development 2009, 136, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltezioti, V.; Kouroupi, G.; Oikonomaki, M.; Mantouvalou, E.; Stergiopoulos, A.; Charonis, A.; Rohrer, H.; Matsas, R.; Politis, P.K. Prox1 regulates the notch1-mediated inhibition of neurogenesis. PLoS Biol. 2010, 8, e1000565. [Google Scholar] [CrossRef] [Green Version]
- Sosa-Pineda, B.; Wigle, J.T.; Oliver, G. Hepatocyte migration during liver development requires Prox1. Nat. Genet. 2000, 25, 254–255. [Google Scholar] [CrossRef]
- Kaltezioti, V.; Foskolou, I.P.; Lavigne, M.D.; Ninou, E.; Tsampoula, M.; Fousteri, M.; Margarity, M.; Politis, P.K. Prox1 inhibits neurite outgrowth during central nervous system development. Cell. Mol. Life Sci. CMLS 2021, 78, 3443–3465. [Google Scholar] [CrossRef]
- Wong, B.W.; Wang, X.; Zecchin, A.; Thienpont, B.; Cornelissen, I.; Kalucka, J.; Garcia-Caballero, M.; Missiaen, R.; Huang, H.; Bruning, U.; et al. The role of fatty acid beta-oxidation in lymphangiogenesis. Nature 2017, 542, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Westmoreland, J.J.; Kilic, G.; Sartain, C.; Sirma, S.; Blain, J.; Rehg, J.; Harvey, N.; Sosa-Pineda, B. Pancreas-specific deletion of Prox1 affects development and disrupts homeostasis of the exocrine pancreas. Gastroenterology 2012, 142, 999–1009.e6. [Google Scholar] [CrossRef] [Green Version]
- Dyer, M.A.; Livesey, F.J.; Cepko, C.L.; Oliver, G. Prox1 function controls progenitor cell proliferation and horizontal cell genesis in the mammalian retina. Nat. Genet. 2003, 34, 53–58. [Google Scholar] [CrossRef]
- Wigle, J.T.; Oliver, G. Prox1 function is required for the development of the murine lymphatic system. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Harvey, N.L.; Srinivasan, R.S.; Dillard, M.E.; Johnson, N.C.; Witte, M.H.; Boyd, K.; Sleeman, M.W.; Oliver, G. Lymphatic vascular defects promoted by Prox1 haploinsufficiency cause adult-onset obesity. Nat. Genet. 2005, 37, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Elsir, T.; Smits, A.; Lindstrom, M.S.; Nister, M. Transcription factor PROX1: Its role in development and cancer. Cancer Metastasis Rev. 2012, 31, 793–805. [Google Scholar] [CrossRef]
- Foskolou, I.P.; Stellas, D.; Rozani, I.; Lavigne, M.D.; Politis, P.K. Prox1 suppresses the proliferation of neuroblastoma cells via a dual action in p27-Kip1 and Cdc25A. Oncogene 2013, 32, 947–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudarzi, K.M.; Espinoza, J.A.; Guo, M.; Bartek, J.; Nister, M.; Lindstrom, M.S.; Hagerstrand, D. Reduced Expression of PROX1 Transitions Glioblastoma Cells into a Mesenchymal Gene Expression Subtype. Cancer Res. 2018, 78, 5901–5916. [Google Scholar] [CrossRef] [Green Version]
- Hogstrom, J.; Heino, S.; Kallio, P.; Lahde, M.; Leppanen, V.M.; Balboa, D.; Wiener, Z.; Alitalo, K. Transcription Factor PROX1 Suppresses Notch Pathway Activation via the Nucleosome Remodeling and Deacetylase Complex in Colorectal Cancer Stem-like Cells. Cancer Res. 2018, 78, 5820–5832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saglietti, C.; La Rosa, S.; Sykiotis, G.P.; Letovanec, I.; Bulliard, J.L.; Piana, S.; Mermod, M.; Petrova, T.; Uccella, S.; Sessa, F.; et al. Expression of Prox1 in Medullary Thyroid Carcinoma Is Associated with Chromogranin A and Calcitonin Expression and with Ki67 Proliferative Index, but Not with Prognosis. Endocr. Pathol. 2019, 30, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Saukkonen, K.; Hagstrom, J.; Mustonen, H.; Juuti, A.; Nordling, S.; Kallio, P.; Alitalo, K.; Seppanen, H.; Haglund, C. PROX1 and beta-catenin are prognostic markers in pancreatic ductal adenocarcinoma. BMC Cancer 2016, 16, 472. [Google Scholar] [CrossRef] [Green Version]
- Dudas, J.; Mansuroglu, T.; Moriconi, F.; Haller, F.; Wilting, J.; Lorf, T.; Fuzesi, L.; Ramadori, G. Altered regulation of Prox1-gene-expression in liver tumors. BMC Cancer 2008, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, T.A.; Wang, Z.; Wu, X.; Kernstine, K.H.; Riggs, A.D.; Pfeifer, G.P. DNA methylation biomarkers for lung cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2012, 33, 287–296. [Google Scholar] [CrossRef]
- Rodrigues, M.; de Oliveira Rodini, C.; de Aquino Xavier, F.C.; Paiva, K.B.; Severino, P.; Moyses, R.A.; Lopez, R.M.; DeCicco, R.; Rocha, L.A.; Carvalho, M.B.; et al. PROX1 gene is differentially expressed in oral cancer and reduces cellular proliferation. Medicine 2014, 93, e192. [Google Scholar] [CrossRef]
- Shimoda, M.; Takahashi, M.; Yoshimoto, T.; Kono, T.; Ikai, I.; Kubo, H. A homeobox protein, prox1, is involved in the differentiation, proliferation, and prognosis in hepatocellular carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 6005–6011. [Google Scholar] [CrossRef] [Green Version]
- Benevenuto de Andrade, B.A.; Ramirez-Amador, V.; Anaya-Saavedra, G.; Martinez-Mata, G.; Fonseca, F.P.; Graner, E.; Paes de Almeida, O. Expression of PROX-1 in oral Kaposi’s sarcoma spindle cells. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2014, 43, 132–136. [Google Scholar] [CrossRef]
- Choi, D.; Ramu, S.; Park, E.; Jung, E.; Yang, S.; Jung, W.; Choi, I.; Lee, S.; Kim, K.E.; Seong, Y.J.; et al. Aberrant Activation of Notch Signaling Inhibits PROX1 Activity to Enhance the Malignant Behavior of Thyroid Cancer Cells. Cancer Res. 2016, 76, 582–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, T.V.; Nykanen, A.; Norrmen, C.; Ivanov, K.I.; Andersson, L.C.; Haglund, C.; Puolakkainen, P.; Wempe, F.; von Melchner, H.; Gradwohl, G.; et al. Transcription factor PROX1 induces colon cancer progression by promoting the transition from benign to highly dysplastic phenotype. Cancer Cell 2008, 13, 407–419. [Google Scholar] [CrossRef]
- Versmold, B.; Felsberg, J.; Mikeska, T.; Ehrentraut, D.; Kohler, J.; Hampl, J.A.; Rohn, G.; Niederacher, D.; Betz, B.; Hellmich, M.; et al. Epigenetic silencing of the candidate tumor suppressor gene PROX1 in sporadic breast cancer. Int. J. Cancer 2007, 121, 547–554. [Google Scholar] [CrossRef]
- Morton, L.M.; Sampson, J.N.; Armstrong, G.T.; Chen, T.H.; Hudson, M.M.; Karlins, E.; Dagnall, C.L.; Li, S.A.; Wilson, C.L.; Srivastava, D.K.; et al. Genome-Wide Association Study to Identify Susceptibility Loci That Modify Radiation-Related Risk for Breast Cancer After Childhood Cancer. J. Natl. Cancer Inst. 2017, 109, djx058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheridan, J.M.; Ritchie, M.E.; Best, S.A.; Jiang, K.; Beck, T.J.; Vaillant, F.; Liu, K.; Dickins, R.A.; Smyth, G.K.; Lindeman, G.J.; et al. A pooled shRNA screen for regulators of primary mammary stem and progenitor cells identifies roles for Asap1 and Prox1. BMC Cancer 2015, 15, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Vaupel, P.; Schmidberger, H.; Mayer, A. The Warburg effect: Essential part of metabolic reprogramming and central contributor to cancer progression. Int. J. Radiat. Biol. 2019, 95, 912–919. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Bounds, P.L.; Dang, C.V. Otto Warburg’s contributions to current concepts of cancer metabolism. Nat. Rev. Cancer 2011, 11, 325–337. [Google Scholar] [CrossRef]
- Shestov, A.A.; Liu, X.; Ser, Z.; Cluntun, A.A.; Hung, Y.P.; Huang, L.; Kim, D.; Le, A.; Yellen, G.; Albeck, J.G.; et al. Quantitative determinants of aerobic glycolysis identify flux through the enzyme GAPDH as a limiting step. eLife 2014, 3, e03342. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. The interplay between MYC and HIF in the Warburg effect. In Ernst Schering Foundation Symposium Proceedings; Springer: Berlin/Heidelberg, Germany, 2007; pp. 35–53. [Google Scholar] [CrossRef]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. [Google Scholar] [CrossRef] [Green Version]
- Robey, I.F.; Stephen, R.M.; Brown, K.S.; Baggett, B.K.; Gatenby, R.A.; Gillies, R.J. Regulation of the Warburg effect in early-passage breast cancer cells. Neoplasia 2008, 10, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Kalezic, A.; Udicki, M.; Srdic Galic, B.; Aleksic, M.; Korac, A.; Jankovic, A.; Korac, B. Tissue-Specific Warburg Effect in Breast Cancer and Cancer-Associated Adipose Tissue-Relationship between AMPK and Glycolysis. Cancers 2021, 13, 2731. [Google Scholar] [CrossRef] [PubMed]
- Doherty, J.R.; Cleveland, J.L. Targeting lactate metabolism for cancer therapeutics. J. Clin. Investig. 2013, 123, 3685–3692. [Google Scholar] [CrossRef] [PubMed]
- Stellas, D.; Szabolcs, M.; Koul, S.; Li, Z.; Polyzos, A.; Anagnostopoulos, C.; Cournia, Z.; Tamvakopoulos, C.; Klinakis, A.; Efstratiadis, A. Therapeutic effects of an anti-Myc drug on mouse pancreatic cancer. J. Natl. Cancer Inst. 2014, 106, dju320. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cao, Y.; Zhang, W.; Bergmeier, S.; Qian, Y.; Akbar, H.; Colvin, R.; Ding, J.; Tong, L.; Wu, S.; et al. A small-molecule inhibitor of glucose transporter 1 downregulates glycolysis, induces cell-cycle arrest, and inhibits cancer cell growth in vitro and in vivo. Mol. Cancer Ther. 2012, 11, 1672–1682. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.A.; Pattengale, P.K.; Leder, P. Spontaneous mammary adenocarcinomas in transgenic mice that carry and express MTV/myc fusion genes. Cell 1984, 38, 627–637. [Google Scholar] [CrossRef]
- Cregan, S.P.; MacLaurin, J.; Gendron, T.F.; Callaghan, S.M.; Park, D.S.; Parks, R.J.; Graham, F.L.; Morley, P.; Slack, R.S. Helper-dependent adenovirus vectors: Their use as a gene delivery system to neurons. Gene Ther. 2000, 7, 1200–1209. [Google Scholar] [CrossRef] [Green Version]
- Bartha, A.; Gyorffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef] [PubMed]
- Lanczky, A.; Gyorffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef] [PubMed]
- Kaltezioti, V.; Antoniou, D.; Stergiopoulos, A.; Rozani, I.; Rohrer, H.; Politis, P.K. Prox1 regulates Olig2 expression to modulate binary fate decisions in spinal cord neurons. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 15816–15831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Intlekofer, A.M.; Finley, L.W.S. Metabolic signatures of cancer cells and stem cells. Nat. Metab. 2019, 1, 177–188. [Google Scholar] [CrossRef]
- Du, J.; Yang, M.; Chen, S.; Li, D.; Chang, Z.; Dong, Z. PDK1 promotes tumor growth and metastasis in a spontaneous breast cancer model. Oncogene 2016, 35, 3314–3323. [Google Scholar] [CrossRef]
- Peng, F.; Wang, J.H.; Fan, W.J.; Meng, Y.T.; Li, M.M.; Li, T.T.; Cui, B.; Wang, H.F.; Zhao, Y.; An, F.; et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene 2018, 37, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Bensaad, K.; Tsuruta, A.; Selak, M.A.; Vidal, M.N.; Nakano, K.; Bartrons, R.; Gottlieb, E.; Vousden, K.H. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell 2006, 126, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Han, G.; Li, X.; Kan, Q.; Fan, Z.; Li, Y.; Ji, Y.; Zhao, J.; Zhang, M.; Grigalavicius, M.; et al. Mitochondrial pyruvate carrier function determines cell stemness and metabolic reprogramming in cancer cells. Oncotarget 2017, 8, 46363–46380. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; He, G.; Yan, S.; Chen, C.; Song, L.; Rosol, T.J.; Deng, X. Triple-negative breast cancer: Is there a treatment on the horizon? Oncotarget 2017, 8, 1913–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahba, H.A.; El-Hadaad, H.A. Current approaches in treatment of triple-negative breast cancer. Cancer Biol. Med. 2015, 12, 106–116. [Google Scholar]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. BCR 2011, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. MYC on the path to cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, H. Reprogramming of glucose, fatty acid and amino acid metabolism for cancer progression. Cell. Mol. Life Sci. CMLS 2016, 73, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Weng, Y.; Wang, Y.; Lin, J.; Deng, X.; Shen, B.; Zhan, Q.; Lu, X. miR-934 as a Prognostic Marker Facilitates Cell Proliferation and Migration of Pancreatic Tumor by Targeting PROX1. OncoTargets Ther. 2020, 13, 3389–3399. [Google Scholar] [CrossRef] [Green Version]
- Rust, K.; Tiwari, M.D.; Mishra, V.K.; Grawe, F.; Wodarz, A. Myc and the Tip60 chromatin remodeling complex control neuroblast maintenance and polarity in Drosophila. EMBO J. 2018, 37, e98659. [Google Scholar] [CrossRef]
- Gramolelli, S.; Cheng, J.; Martinez-Corral, I.; Vaha-Koskela, M.; Elbasani, E.; Kaivanto, E.; Rantanen, V.; Tuohinto, K.; Hautaniemi, S.; Bower, M.; et al. PROX1 is a transcriptional regulator of MMP14. Sci. Rep. 2018, 8, 9531. [Google Scholar] [CrossRef] [Green Version]
- Bencivenga, D.; Stampone, E.; Roberti, D.; Della Ragione, F.; Borriello, A. p27(Kip1), an Intrinsically Unstructured Protein with Scaffold Properties. Cells 2021, 10, 2254. [Google Scholar] [CrossRef]
- Dupuy, F.; Tabaries, S.; Andrzejewski, S.; Dong, Z.; Blagih, J.; Annis, M.G.; Omeroglu, A.; Gao, D.; Leung, S.; Amir, E.; et al. PDK1-Dependent Metabolic Reprogramming Dictates Metastatic Potential in Breast Cancer. Cell Metab. 2015, 22, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Gagliardi, P.A.; Puliafito, A.; Primo, L. PDK1: At the crossroad of cancer signaling pathways. Semin. Cancer Biol. 2018, 48, 27–35. [Google Scholar] [CrossRef]
- Liu, T.; Yin, H. PDK1 promotes tumor cell proliferation and migration by enhancing the Warburg effect in non-small cell lung cancer. Oncol. Rep. 2017, 37, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.P.; Chen, Q.; Li, Y.; Wang, Y.; Zou, H.B.; Fu, W.J.; Niu, Q.; Pan, Q.G.; Jiang, P.; Xu, X.S.; et al. Mitochondrial pyruvate carrier 1 functions as a tumor suppressor and predicts the prognosis of human renal cell carcinoma. Lab. Investig. 2019, 99, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Y.; Wang, S.; Dong, Q.Z.; Shen, Z.; Wang, W.; Tao, S.; Gu, C.; Liu, J.; Xie, Y.; et al. Prospero-related homeobox 1 drives angiogenesis of hepatocellular carcinoma through selectively activating interleukin-8 expression. Hepatology 2017, 66, 1894–1909. [Google Scholar] [CrossRef]
- Procino, A. Overexpression of Prox-1 gene in omental adipose tissue and adipocytes compared with subcutaneous adipose tissue and adipocytes in healthy patients. Cell Biol. Int. 2014, 38, 888–891. [Google Scholar] [CrossRef]
- Ragusa, S.; Cheng, J.; Ivanov, K.I.; Zangger, N.; Ceteci, F.; Bernier-Latmani, J.; Milatos, S.; Joseph, J.M.; Tercier, S.; Bouzourene, H.; et al. PROX1 promotes metabolic adaptation and fuels outgrowth of Wnt(high) metastatic colon cancer cells. Cell Rep. 2014, 8, 1957–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Huang, J.; Zhou, J.; Hui, K.; Xu, S.; Fan, J.; Li, L.; Wang, X.; Hsieh, J.T.; He, D.; et al. DAB2IP regulates EMT and metastasis of prostate cancer through targeting PROX1 transcription and destabilizing HIF1alpha protein. Cell. Signal. 2016, 28, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Ueta, K.; Otowa, Y.; Kakeji, Y.; Hirashima, M. PROX1 Is Associated with Cancer Progression and Prognosis in Gastric Cancer. Anticancer Res. 2018, 38, 6139–6145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Ji, S.; Ma, F.; Ma, Q.; Lu, X.; Chen, X. miR-489 acts as a tumor suppressor in human gastric cancer by targeting PROX1. Am. J. Cancer Res. 2016, 6, 2021–2030. [Google Scholar] [PubMed]
- Charest-Marcotte, A.; Dufour, C.R.; Wilson, B.J.; Tremblay, A.M.; Eichner, L.J.; Arlow, D.H.; Mootha, V.K.; Giguere, V. The homeobox protein Prox1 is a negative modulator of ERRα/PGC-1α bioenergetic functions. Genes Dev. 2010, 24, 537–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, J.G.; Gryder, B.E.; Pavlovic, B.; Chung, Y.; Ngo, Q.A.; Frommelt, F.; Gstaiger, M.; Song, Y.; Benischke, K.; Laubscher, D.; et al. NuRD subunit CHD4 regulates super-enhancer accessibility in rhabdomyosarcoma and represents a general tumor dependency. eLife 2020, 9, e54993. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, J.; Yoo, J.; Ganesan, S.K.; Cook, S.C.; Aguilar, B.; Ramu, S.; Lee, J.; Hong, Y.K. Prox1 physically and functionally interacts with COUP-TFII to specify lymphatic endothelial cell fate. Blood 2009, 113, 1856–1859. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, H.; Qin, Y.; Liu, Y.; Xie, Y.; Liu, J. Prox1 directly interacts with LSD1 and recruits the LSD1/NuRD complex to epigenetically co-repress CYP7A1 transcription. PLoS ONE 2013, 8, e62192. [Google Scholar] [CrossRef] [Green Version]
- Takeda, Y.; Jetten, A.M. Prospero-related homeobox 1 (Prox1) functions as a novel modulator of retinoic acid-related orphan receptors alpha- and gamma-mediated transactivation. Nucleic Acids Res. 2013, 41, 6992–7008. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, A.; Politis, P.K. Nuclear receptor NR5A2 controls neural stem cell fate decisions during development. Nat. Commun. 2016, 7, 12230. [Google Scholar] [CrossRef] [Green Version]
- Risebro, C.A.; Petchey, L.K.; Smart, N.; Gomes, J.; Clark, J.; Vieira, J.M.; Yanni, J.; Dobrzynski, H.; Davidson, S.; Zuberi, Z.; et al. Epistatic rescue of Nkx2.5 adult cardiac conduction disease phenotypes by prospero-related homeobox protein 1 and HDAC3. Circ. Res. 2012, 111, e19–e31. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, K.R.; Holter, E.; Bavner, A.; Nilsson, M.; Pelto-Huikko, M.; Tomarev, S.; Treuter, E. Functional conservation of interactions between a homeodomain cofactor and a mammalian FTZ-F1 homologue. EMBO Rep. 2004, 5, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Dowhan, D.H.; Liang, D.; Moore, D.D.; Overbeek, P.A. CREB-binding protein/p300 co-activation of crystallin gene expression. J. Biol. Chem. 2002, 277, 24081–24089. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence | |
|---|---|---|
| hPROX1 | forward | ATCCCAGCTCCAATATGCTG |
| reverse | TTGACGTGCGTACTTCTCCA | |
| hCYCLIN D1 | forward | CCCTCGGTGTCCTACTTCAA |
| reverse | AGGAAGCGGTCCAGGTAGTT | |
| hCYCLIN E1 | forward | ATCCTCCAAAGTTGCACCAG |
| reverse | AGGGGACTTAAACGCCACTT | |
| hp27-kip1 | forward | AGAGTTAACCCGGGACTTGG |
| reverse | GCCCTCTAGGGGTTTGTGAT | |
| hp21-cip1 | forward | GGAAGACCATGTGGACCTGT |
| reverse | GGCGTTTGGAGTGGTAGAAA | |
| hIRS1 | forward | ATGAGTGATGAGTTCCGCCC |
| reverse | TGATGCTCTCAGTGCGTGAT | |
| hEGFR | forward | ATGTGGTGACAGATCACGGC |
| reverse | AGGCCCTTCGCACTTCTTAC | |
| hFOXA | forward | TACGCAGACACGCAGGAG |
| reverse | CCGCTCGTAGTCATGGTGTT | |
| hmTOR | forward | AACGAGCTGGTCCGAATCAG |
| reverse | AGGTTTTGTTCCGAAGCCCA | |
| hc-MYC | forward | CGTCCTCGGATTCTCTGCTC |
| reverse | GCCTGCCTCTTTTCCACAGA | |
| hHES1 | forward | ATGACAGTGAAGCACCTCCG |
| reverse | CGTTCATGCACTCGCTGAAG | |
| hGLUT-3 | forward | TGGCCCAGATCTTTGGTCTG |
| reverse | ATGGAAGGGCTGCACTTTGT | |
| hGLUT-1 | forward | TCTCGAAACTGGGCAAGTCC |
| reverse | TCACCCACATACATGGGCAC | |
| hHK2 | forward | ACAAATTTCCGGGTCCTGCT |
| reverse | TGTGGTCAAAGAGCTCGTCC | |
| hPDK1 | forward | TGCTGTATGGCCTGCAAGAT |
| reverse | ACATTCTGGCTGGTGACAGG | |
| hENO-2 | forward | CCTGCAATGTGACGGAGAGT |
| reverse | TTACACACGGCCAGAGACAC | |
| hTigar | forward | GGTTGTAGAAGGCAAAGCGC |
| reverse | ATTTTCACCTGGTCCAGCGT | |
| hCOX4i | forward | GCAGCCTCTCCATGGATGAG |
| reverse | CACAACCGTCTTCCACTCGT | |
| hMPC1 | forward | TCTTCCCATTGCTGCCATCA |
| reverse | AACAGAAGCCAGTTCCGAGG | |
| hNDUFA7 | forward | TGCGACGACATGATGATGGA |
| reverse | GCCCTCTAGGGGTTTGTGAT | |
| hGAPDH2 | forward | CCAGTATGAACTCCACTCACG |
| reverse | CTCCTGGAAGATTGGTGATGG | |
| hPPIA | forward | TGGACCCAACACAAATGGT |
| reverse | ATGCCTTCTTTCACTTTGCC | |
| hRPL13 | forward | GCGGACCCGTGCCGAGGTTAT |
| reverse | CACCATCCGCTTTTTCTTGCT | |
| hp16 | forward | GAGCAGCATGGAGCCTTC |
| reverse | CATCATCATGACCTGGATCG | |
| hSTAT3 | forward | CAACTTCAGACCCGTCAACA |
| reverse | CGATTCTCTCCTCCAGCATC | |
| Primers Name | Orientation | Sequence |
|---|---|---|
| Locus A | forward | CCCCTCCCATATTCTCCCGT |
| reverse | GAGACAAATCCCCTTTGCGC | |
| Locus B | forward | CAGGCAGACACATCTCAGGG |
| reverse | ACCTTCCACCCAGACTGAGT | |
| Locus C | forward | ACTCAGTCTGGGTGGAAGGT |
| reverse | CGTATACTTGGAGAGCGCGT | |
| Locus D | forward | AGTGTTCTTGGTAAAGTCCCTCA |
| reverse | GACATTGATGCCAATTCTTACCT | |
| Locus E | forward | AGGCAGGTGAGAAGGTGAGA |
| reverse | TCCTCACCTTCCTCCCAACT | |
| Locus F | forward | TGACTCACTTGGGAATCGGG |
| reverse | TGGAGAGTTGTTTTCCTTGCT | |
| Locus G | forward | TGACTATGCCCTTTATCCATGACA |
| reverse | TCCTTCCTCTTTATACATTCCATCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michail, A.; Gkikas, D.; Stellas, D.; Kaltezioti, V.; Politis, P.K. Prox1 Suppresses the Proliferation of Breast Cancer Cells via Direct Inhibition of c-Myc Gene Expression. Cells 2023, 12, 1869. https://doi.org/10.3390/cells12141869
Michail A, Gkikas D, Stellas D, Kaltezioti V, Politis PK. Prox1 Suppresses the Proliferation of Breast Cancer Cells via Direct Inhibition of c-Myc Gene Expression. Cells. 2023; 12(14):1869. https://doi.org/10.3390/cells12141869
Chicago/Turabian StyleMichail, Artemis, Dimitrios Gkikas, Dimitris Stellas, Valeria Kaltezioti, and Panagiotis K. Politis. 2023. "Prox1 Suppresses the Proliferation of Breast Cancer Cells via Direct Inhibition of c-Myc Gene Expression" Cells 12, no. 14: 1869. https://doi.org/10.3390/cells12141869