

Functional Diversity of Novel Lectins with Unique Structural Features in Marine Animals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Involvement of C-Type Lectins in Immunity
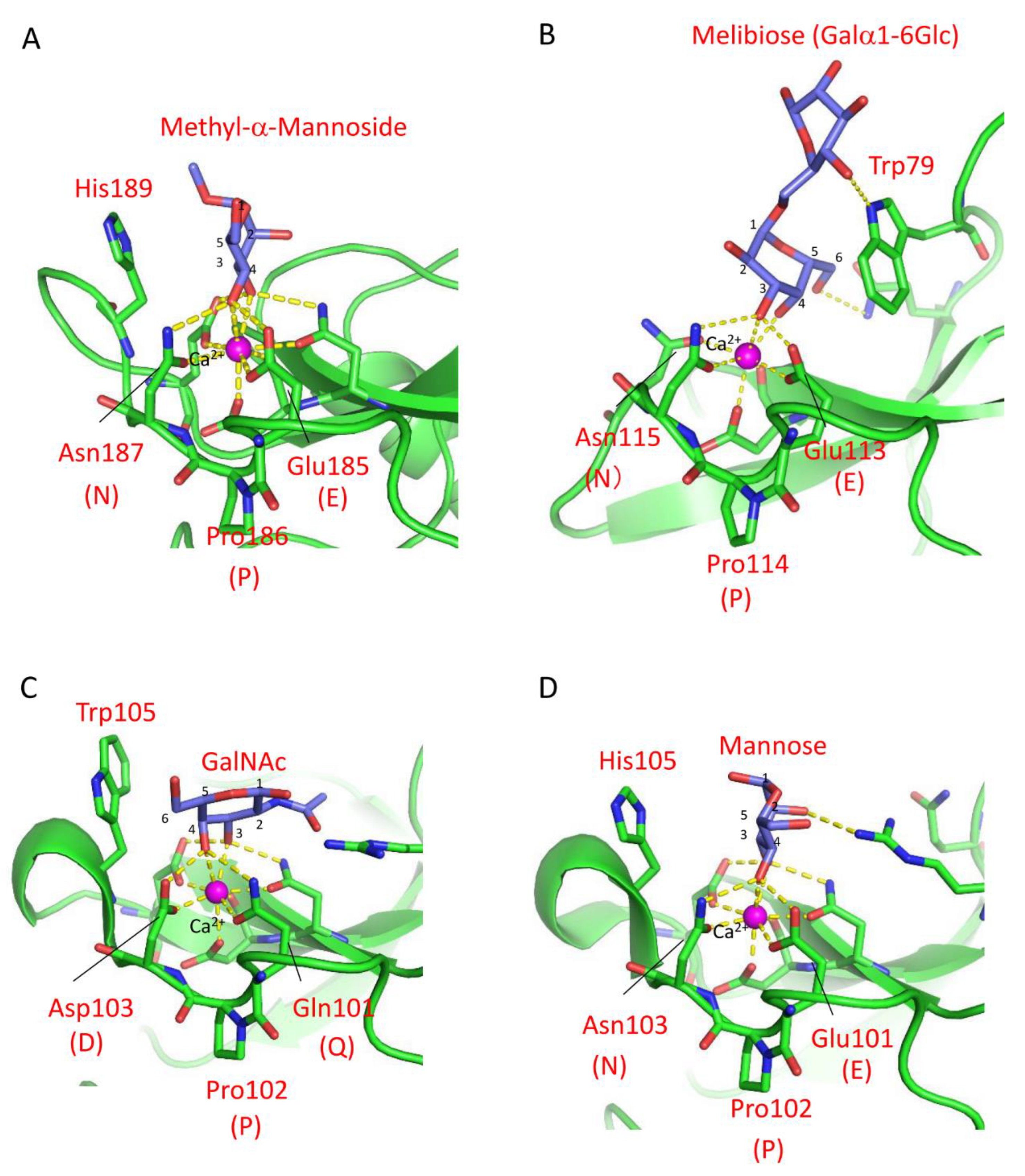
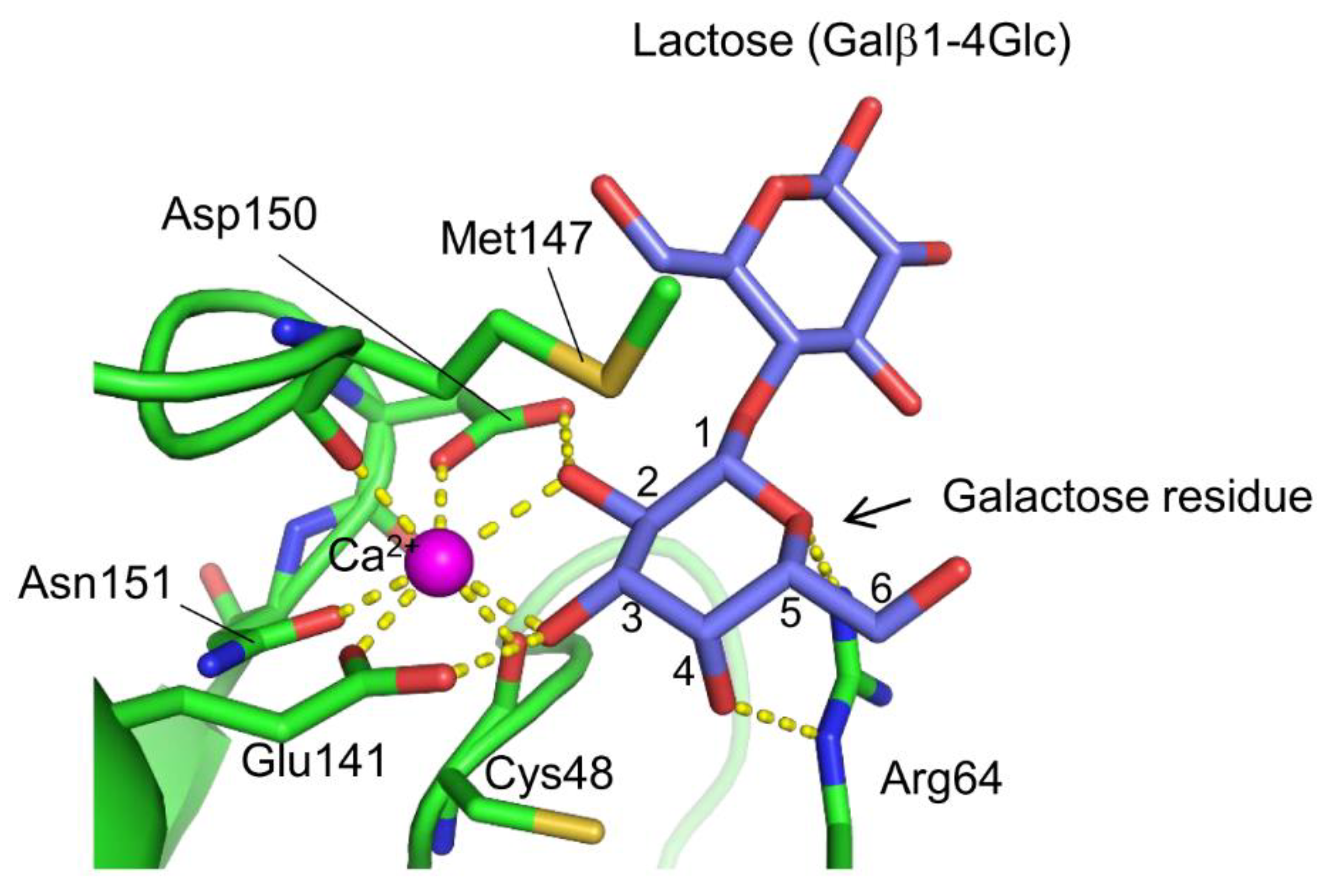
3. Carbohydrate Recognition Mechanisms of Marine Invertebrate C-Type Lectins
4. Marine Animal Lectins with Novel Structures and Carbohydrate-Recognition Mechanisms
4.1. Mannose-Binding Lectin CGL1 from the Pacific Oyster
4.2. Galactose-Binding Lectin AJLec from Sea Anemone
5. Lectins as Toxins from Marine Animals
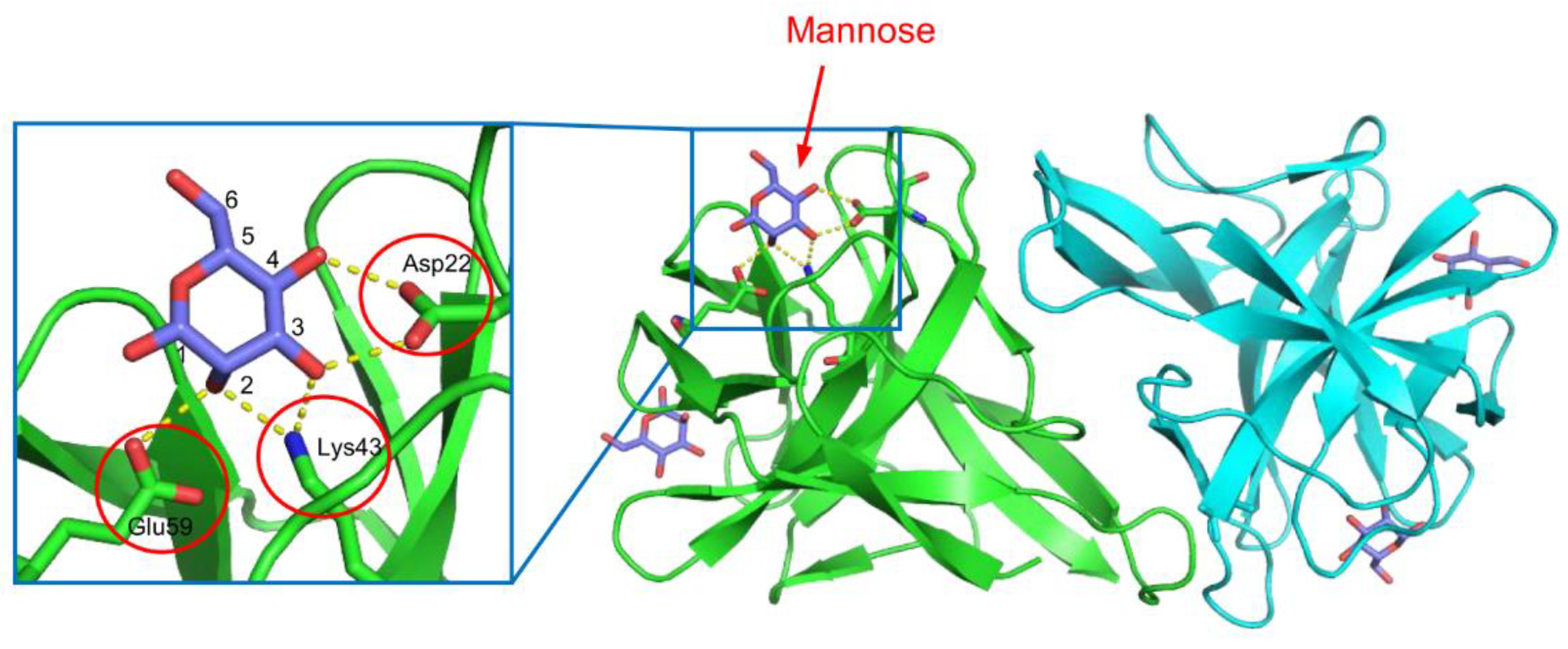
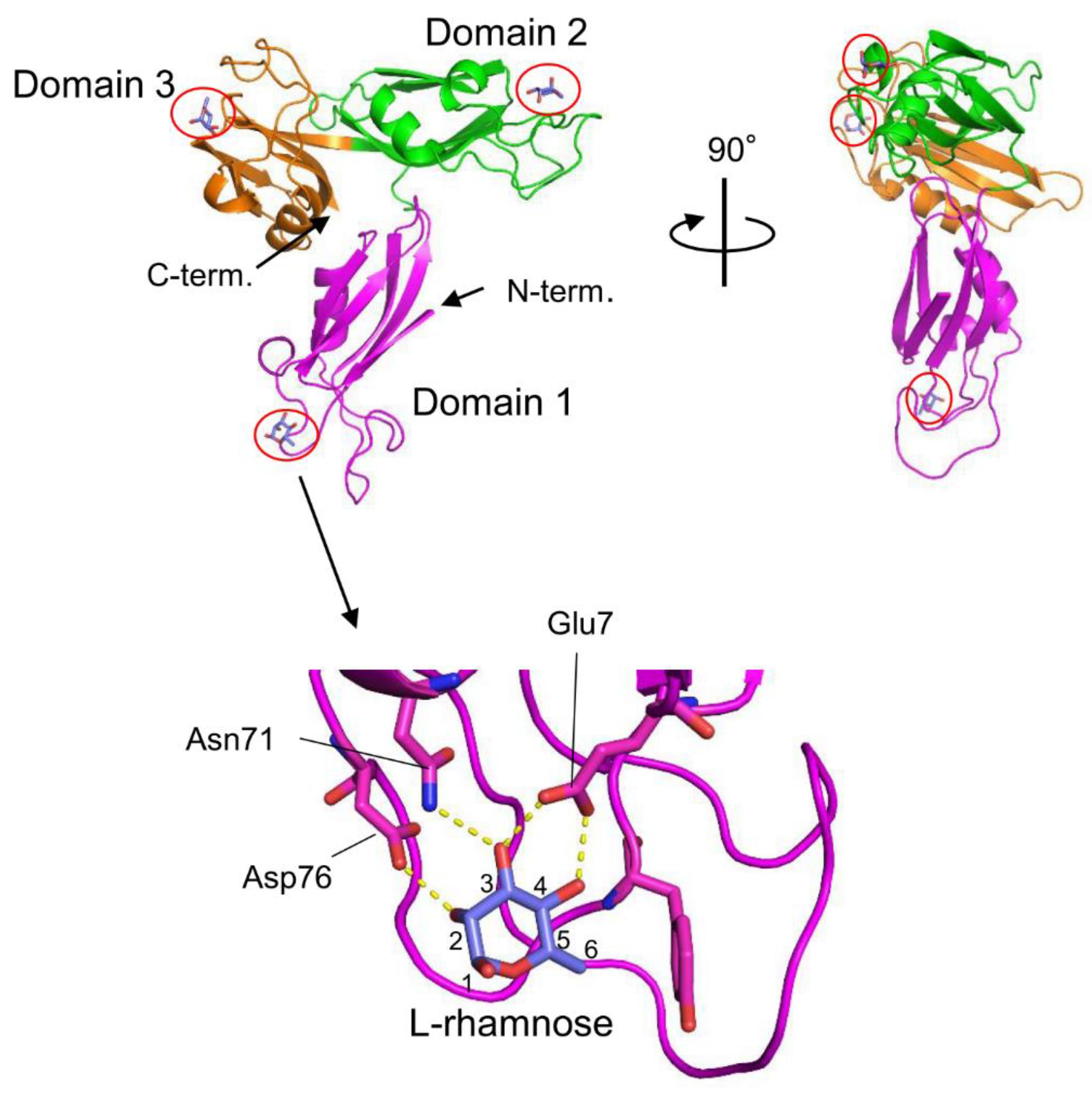
5.1. Lectin from the Venomous Sea Urchin Toxopneustes pileolus
5.2. Hemolytic Lectin from the Sea Cucumber C. echinata
5.3. Fish Spine Toxins Containing Lectin-like Domains
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharon, N.; Lis, H. History of Lectins: From Hemagglutinins to Biological Recognition Molecules. Glycobiology 2004, 14, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Kaltner, H.; Gabius, H.J. Sensing Glycans as Biochemical Messages by Tissue Lectins: The Sugar Code at Work in Vascular Biology. Thromb. Haemost. 2019, 119, 517–533. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, D.C. Animal Lectins: A Historical Introduction and Overview. Biochim. Biophys. Acta Gen. Subj. 2002, 1572, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.J. Animal Lectins. Eur. J. Biochem. 1997, 243, 543–576. [Google Scholar] [CrossRef] [PubMed]
- Leffler, H.; Carlsson, S.; Hedlund, M.; Qian, Y.; Poirier, F. Introduction to Galectins. Glycoconj. J. 2002, 19, 433–440. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Gready, J.E. The C-Type Lectin-like Domain Superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Modenutti, C.P.; Capurro, J.I.B.; Di Lella, S.; Martí, M.A. The Structural Biology of Galectin-Ligand Recognition: Current Advances in Modeling Tools, Protein Engineering, and Inhibitor Design. Front. Chem. 2019, 7, 823. [Google Scholar] [CrossRef]
- Drickamer, K. Two Distinct Classes of Carbohydrate-Recognition Domains in Animal Lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar] [CrossRef]
- Drickamer, K. C-Type Lectin-like Domains. Curr. Opin. Struct. Biol. 1999, 9, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-Type Lectin Superfamily in the Immune System. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Willment, J.A.; Whitehead, L. C-Type Lectins in Immunity and Homeostasis. Nat. Rev. Immunol. 2018, 18, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Cambi, A.; Koopman, M.; Figdor, C.G. How C-Type Lectins Detect Pathogens. Cell. Microbiol. 2005, 7, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Kuribara, T.; Usui, R.; Totani, K. Glycan Structure-Based Perspectives on the Entry and Release of Glycoproteins in the Calnexin/Calreticulin Cycle. Carbohydr. Res. 2021, 502, 108273. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, G.; Gehring, K. Calnexin Cycle—Structural Features of the ER Chaperone System. FEBS J. 2020, 287, 4322–4340. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, K.; Simons, K. A Putative Novel Class of Animal Lectins in the Secretory Pathway Homologous to Leguminous Lectins. Cell 1994, 77, 625–626. [Google Scholar] [CrossRef]
- Dahms, N.M.; Hancock, M.K. P-Type Lectins. Biochim. Biophys. Acta Gen. Subj. 2002, 1572, 317–340. [Google Scholar] [CrossRef]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and Their Roles in the Immune System. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef]
- Duan, S.; Paulson, J.C. Siglecs as Immune Cell Checkpoints in Disease. Annu. Rev. Immunol. 2020, 38, 365–395. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, G.; Weigle, B.; Crocker, P.R. Siglec and Anti-Siglec Therapies. Curr. Opin. Chem. Biol. 2021, 62, 34–42. [Google Scholar] [CrossRef]
- Viana, J.T.; Rocha, R.; Maggioni, R. Structural and Functional Diversity of Lectins Associated with Immunity in the Marine Shrimp Litopenaeus Vannamei. Fish Shellfish Immunol. 2022, 129, 152–160. [Google Scholar] [CrossRef]
- Vasta, G.R.; Feng, C.; González-Montalbán, N.; Mancini, J.; Yang, L.; Abernathy, K.; Frost, G.; Palm, C. Functions of Galectins as ’Self/Non-Self’-Recognition and Effector Factors. Pathog. Dis. 2017, 75, ftx046. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.Z.; Wang, L.; Xu, S.; Zhang, X.W.; Zhao, X.F.; Vasta, G.R.; Wang, J.X. A Galectin from the Kuruma Shrimp (Marsupenaeus Japonicus) Functions as an Opsonin and Promotes Bacterial Clearance from Hemolymph. PLoS ONE 2014, 9, 794. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Agius, J.; Ackerly, D.; Beddoe, T.; Helbig, K. The Role of Anti-Viral Effector Molecules in Mollusc Hemolymph. Biomolecules 2022, 12, 345. [Google Scholar] [CrossRef]
- Patnaik, B.B.; Baliarsingh, S.; Sarkar, A.; Hameed, A.S.S.; Lee, Y.S.; Jo, Y.H.; Han, Y.S.; Mohanty, J. The Role of Pattern Recognition Receptors in Crustacean Innate Immunity. Rev. Aquac. 2023, 1–44. [Google Scholar] [CrossRef]
- Wang, W.; Song, X.; Wang, L.; Song, L. Pathogen-Derived Carbohydrate Recognition in Molluscs Immune Defense. Int. J. Mol. Sci. 2018, 19, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, W.I.; Kahn, R.; Fourme, R.; Drickamer, K.; Hendrickson, W.A. Structure of the Calcium-Dependent Lectin Domain from a Rat Mannose-Binding Protein Determined by MAD Phasing. Science 1991, 254, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C. Mannan-Binding Lectin: Clinical Significance and Applications. Biochim. Biophys. Acta Gen. Subj. 2002, 1572, 401–413. [Google Scholar] [CrossRef]
- Świerzko, A.S.; Cedzyński, M. The Influence of the Lectin Pathway of Complement Activation on Infections of the Respiratory System. Front. Immunol. 2020, 11, 243. [Google Scholar] [CrossRef]
- Kjaer, T.R.; Le, L.T.M.; Pedersen, J.S.; Sander, B.; Golas, M.M.; Jensenius, J.C.; Andersen, G.R.; Thiel, S. Structural Insights into the Initiating Complex of the Lectin Pathway of Complement Activation. Structure 2015, 23, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Teh, C.; Kishore, U.; Reid, K.B.M. Collectins and Ficolins: Sugar Pattern Recognition Molecules of the Mammalian Innate Immune System. Nefrol. Biophys. Acta 2002, 1572, 387–400. [Google Scholar] [CrossRef]
- del Fresno, C.; Iborra, S.; Saz-Leal, P.; Martínez-López, M.; Sancho, D. Flexible Signaling of Myeloid C-Type Lectin Receptors in Immunity and Inflammation. Front. Immunol. 2018, 9, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Wang, L.; Zhang, H.; Yang, C.; Liu, R.; Xu, J.; Jia, Z.; Song, L. The Sequence Variation and Functional Differentiation of CRDs in a Scallop Multiple CRDs Containing Lectin. Dev. Comp. Immunol. 2017, 67, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Huang, M.; Zhang, H.; Wang, L.L.; Wang, H.; Wang, L.L.; Qiu, L.; Song, L. CfLec-3 from Scallop: An Entrance to Non-Self Recognition Mechanism of Invertebrate C-Type Lectin. Sci. Rep. 2015, 5, 068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-Type Lectins in Innate Immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef]
- de Melo, A.A.; Carneiro, R.F.; de Melo Silva, W.; da Moura, R.M.; Silva, G.C.; de Sousa, O.V.; de Sousa Saboya, J.P.; do Nascimento, K.S.; Saker-Sampaio, S.; Nagano, C.S.; et al. HGA-2, a Novel Galactoside-Binding Lectin from the Sea Cucumber Holothuria Grisea Binds to Bacterial Cells. Int. J. Biol. Macromol. 2014, 64, 435–442. [Google Scholar] [CrossRef]
- Kvennefors, E.C.E.; Leggat, W.; Hoegh-Guldberg, O.; Degnan, B.M.; Barnes, A.C. An Ancient and Variable Mannose-Binding Lectin from the Coral Acropora Millepora Binds Both Pathogens and Symbionts. Dev. Comp. Immunol. 2008, 32, 1582–1592. [Google Scholar] [CrossRef]
- Runsaeng, P.; Puengyam, P.; Utarabhand, P. A Mannose-Specific C-Type Lectin from Fenneropenaeus Merguiensis Exhibited Antimicrobial Activity to Mediate Shrimp Innate Immunity. Mol. Immunol. 2017, 92, 87–98. [Google Scholar] [CrossRef]
- Tetreau, G.; Pinaud, S.; Portet, A.; Galinier, R.; Gourbal, B.; Duval, D. Specific Pathogen Recognition by Multiple Innate Immune Sensors in an Invertebrate. Front. Immunol. 2017, 8, 1249. [Google Scholar] [CrossRef]
- Wang, X.-W.; Gao, J.; Xu, Y.-H.; Xu, J.-D.; Fan, Z.-X.; Zhao, X.-F.; Wang, J.-X. Novel Pattern Recognition Receptor Protects Shrimp by Preventing Bacterial Colonization and Promoting Phagocytosis. J. Immunol. 2017, 198, 3045–3057. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, L.; Zhang, D.; Li, F.; Wang, M.; Huang, M.; Zhang, H.; Song, L. A Novel C-Type Lectin from Crab Eriocheir Sinensis Functions as Pattern Recognition Receptor Enhancing Cellular Encapsulation. Fish Shellfish Immunol. 2013, 34, 832–842. [Google Scholar] [CrossRef]
- Zhang, X.W.; Xu, W.T.; Wang, X.W.; Mu, Y.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A Novel C-Type Lectin with Two CRD Domains from Chinese Shrimp Fenneropenaeus Chinensis Functions as a Pattern Recognition Protein. Mol. Immunol. 2009, 46, 1626–1637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Guo, X.; Litman, G.W.; Dishaw, L.J.; Zhang, G. Massive Expansion and Functional Divergence of Innate Immune Genes in a Protostome. Sci. Rep. 2015, 5, 693. [Google Scholar] [CrossRef] [Green Version]
- Pees, B.; Yang, W.; Zárate-Potes, A.; Schulenburg, H.; Dierking, K. High Innate Immune Specificity through Diversified C-Type Lectin-like Domain Proteins in Invertebrates. J. Innate Immun. 2016, 8, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Saco, A.; Suárez, H.; Novoa, B.; Figueras, A. A Genomic and Transcriptomic Analysis of the C-Type Lectin Gene Family Reveals Highly Expanded and Diversified Repertoires in Bivalves. Mar. Drugs 2023, 21, 254. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Engineering Galactose-Binding Activity into a C-Type Mannose-Binding Protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, L.; Zhang, H.; Qiu, L.; Wang, H.; Song, L. C-Type Lectin in Chlamys Farreri (CfLec-1) Mediating Immune Recognition and Opsonization. PLoS ONE 2011, 6, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhao, X.; Wang, Z.; Shao, Y.; Zhang, W.; Bao, Y.; Li, C. Novel Ca2+-Independent C-Type Lectin Involved in Immune Defense of the Razor Clam Sinonovacula Constricta. Fish Shellfish Immunol. 2019, 84, 502–508. [Google Scholar] [CrossRef]
- Lin, J.Y.; Ma, K.Y.; Bai, Z.Y.; Li, J. Le Molecular Cloning and Characterization of Perlucin from the Freshwater Pearl Mussel, Hyriopsis Cumingii. Gene 2013, 526, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Song, X.; Wang, L.; Kong, P.; Yang, J.; Liu, L.; Qiu, L.; Zhang, Y.; Qiu, L.; Song, L. AiCTL-6, a Novel C-Type Lectin from Bay Scallop Argopecten Irradians with a Long C-Type Lectin-like Domain. Fish Shellfish Immunol. 2011, 30, 17–26. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Kamiya, T.; Kusunoki, M.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Goda, S.; Unno, H. Galactose Recognition by a Tetrameric C-Type Lectin, CEL-IV, Containing the EPN Carbohydrate Recognition Motif. J. Biol. Chem. 2011, 286, 576. [Google Scholar] [CrossRef] [Green Version]
- Iobst, S.T.; Drickamer, K. Binding of Sugar Ligands to Ca2+-Dependent Animal Lectins. II. Generation of High-Affinity Galactose Binding by Site-Directed Mutagenesis. J. Biol. Chem. 1994, 269, 15512–15519. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Ishimine, T.; Baba, T.; Kimura, M.; Unno, H.; Goda, S. Alteration of the Carbohydrate-Binding Specificity of a C-Type Lectin CEL-I Mutant with an EPN Carbohydrate-Binding Motif. Protein Pept. Lett. 2013, 20, 796–801. [Google Scholar] [CrossRef]
- Moriuchi, H.; Unno, H.; Goda, S.; Tateno, H.; Hirabayashi, J.; Hatakeyama, T. Mannose-Recognition Mutant of the Galactose/N-Acetylgalactosamine-Specific C-Type Lectin CEL-I Engineered by Site-Directed Mutagenesis. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Unno, H.; Itakura, S.; Higuchi, S.; Goda, S.; Yamaguchi, K.; Hatakeyama, T. Novel Ca2+-Independent Carbohydrate Recognition of the C-Type Lectins, SPL-1 and SPL-2, from the Bivalve Saxidomus Purpuratus. Protein Sci. 2019, 28, 766–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liao, K.; Shi, P.; Xu, J.; Ran, Z.; Zhou, C.; Zhang, L.; Cao, J.; Yan, X. Involvement of a Novel Ca2+-Independent C-Type Lectin from Sinonovacula Constricta in Food Recognition and Innate Immunity. Fish Shellfish Immunol. 2020, 104, 374–382. [Google Scholar] [CrossRef]
- Unno, H.; Higuchi, S.; Goda, S.; Hatakeyama, T. Novel Carbohydrate-Recognition Mode of the Invertebrate C-Type Lectin SPL-1 from Saxidomus Purpuratus Revealed by the GlcNAc-Complex Crystal in the Presence of Ca2+. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2020, 76, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Unno, H.; Matsuyama, K.; Tsuji, Y.; Goda, S.; Hiemori, K.; Tateno, H.; Hirabayashi, J.; Hatakeyama, T. Identification, Characterization, and X-Ray Crystallographic Analysis of a Novel Type of Mannose-Specific Lectin CGL1 from the Pacific Oyster Crassostrea Gigas. Sci. Rep. 2016, 6, 29135. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Wang, L.; Huang, M.; Jia, Z.; Weinert, T.; Warkentin, E.; Liu, C.; Song, X.; Zhang, H.; Witt, J.; et al. DM9 Domain Containing Protein Functions as a Pattern Recognition Receptor with Broad Microbial Recognition Spectrum. Front. Immunol. 2017, 8, 1607. [Google Scholar] [CrossRef] [Green Version]
- Ponting, C.P.; Mott, R.; Bork, P.; Copley, R.R. Novel Protein Domains and Repeats in Drosophila Melanogaster: Insights into Structure, Function, and Evolution. Genome Res. 2001, 11, 1996–2008. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, P.; Wang, W.; Dong, M.; Wang, M.; Gong, C.; Jia, Z.; Liu, Z.; Zhang, A.; Wang, L.; et al. A DM9-Containing Protein from Oyster Crassostrea Gigas (CgDM9CP-2) Serves as a Multipotent Pattern Recognition Receptor. Dev. Comp. Immunol. 2018, 84, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, W.; Zhao, Q.; Yuan, P.; Li, J.; Song, X.; Liu, Z.; Ding, D.; Wang, L.; Song, L. A DM9-Containing Protein from Oyster Crassostrea gigas (CgDM9CP-3) Mediating Immune Recognition and Encapsulation. Dev. Comp. Immunol. 2021, 116, 103937. [Google Scholar] [CrossRef] [PubMed]
- Phadungsil, W.; Grams, R. Agglutination Activity of Fasciola Gigantica DM9-1, a Mannose-Binding Lectin. Korean J. Parasitol. 2021, 59, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Phadungsil, W.; Smooker, P.M.; Vichasri-Grams, S.; Grams, R. Characterization of a Fasciola Gigantica Protein Carrying Two DM9 Domains Reveals Cellular Relocalization Property. Mol. Biochem. Parasitol. 2016, 205, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Chertemps, T.; Mitri, C.; Perrot, S.; Sautereau, J.; Jacques, J.C.; Thiery, I.; Bourgouin, C.; Rosinski-Chupin, I. Anopheles Gambiae PRS1 Modulates Plasmodium Development at Both Midgut and Salivary Gland Steps. PLoS ONE 2010, 5, 538. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, G.S.; Lopes-Ferreira, M.; Junqueira-De-Azevedo, I.L.M.; Spencer, P.J.; Araújo, M.S.; Portaro, F.C.V.; Ma, L.; Valente, R.H.; Juliano, L.; Fox, J.W.; et al. Natterins, a New Class of Proteins with Kininogenase Activity Characterized from Thalassophryne Nattereri Fish Venom. Biochimie 2005, 87, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, T.; Kishigawa, A.; Unno, H. Molecular Cloning and Characterization of the Two Putative Toxins Expressed in the Venom of the Devil Stinger Inimicus Japonicus. Toxicon 2021, 201, 9–20. [Google Scholar] [CrossRef]
- Unno, H.; Nakamura, A.; Mori, S.; Goda, S.; Yamaguchi, K.; Hiemori, K.; Tateno, H.; Hatakeyama, T. Identification, Characterization, and X-Ray Crystallographic Analysis of a Novel Type of Lectin AJLec from the Sea Anemone Anthopleura Japonica. Sci. Rep. 2018, 8, 980. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.E.; Drickamer, K. Mammalian Sugar-Binding Receptors: Known Functions and Unexplored Roles. FEBS J. 2019, 286, 1800–1814. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.E.; Drickamer, K. Convergent and Divergent Mechanisms of Sugar Recognition across Kingdoms. Curr. Opin. Struct. Biol. 2014, 28, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Olsnes, S. The History of Ricin, Abrin and Related Toxins. Toxicon 2004, 44, 361–370. [Google Scholar] [CrossRef]
- Lord, J.M.; Roberts, L.M.; Robertus, J.D. Ricin: Structure, Mode of Action, and Some Current Applications. FASEB J. 1994, 8, 201–208. [Google Scholar] [CrossRef]
- Hartley, M.R.; Lord, J.M. Cytotoxic Ribosome-Inactivating Lectins from Plants. Biochim. Biophys. Acta—Proteins Proteom. 2004, 1701, 1–14. [Google Scholar] [CrossRef]
- Odumosu, O.; Nicholas, D.; Yano, H.; Langridge, W. AB Toxins: A Paradigm Switch from Deadly to Desirable. Toxins 2010, 2, 1612–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.A.; Abe, J.; Siddiq, A.; Nakagawa, H.; Honda, S.; Wada, T.; Ichida, S. UT841 Purified from Sea Urchin (Toxopneustes Pileolus) Venom Inhibits Time-Dependent 45Ca2+ Uptake in Crude Synaptosome Fraction from Chick Brain. Toxicon 2001, 39, 1223–1229. [Google Scholar] [CrossRef]
- Nakagawa, H.; Hashimoto, T.; Hayashi, H.; Shinohara, M.; Ohura, K.; Tachikawa, E. Isolation of a Novel Lectin from the Globiferous Pedicellariae of the Sea Urchin Toxopneustes Pileolus. Adv. Exp. Med. Biol. 1996, 391, 213–223. [Google Scholar]
- Takei, M.; Nakagawa, H. A Sea Urchin Lectin, SUL-1, from the Toxopneustid Sea Urchin Induces DC Maturation from Human Monocyte and Drives Th1 Polarization in Vitro. Toxicol. Appl. Pharmacol. 2006, 213, 27–36. [Google Scholar] [CrossRef]
- Tateno, H. SUEL-Related Lectins, a Lectin Family Widely Distributed throughout Organisms. Biosci. Biotechnol. Biochem. 2010, 74, 1141–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, T.; Ichise, A.; Yonekura, T.; Unno, H.; Goda, S.; Nakagawa, H. CDNA Cloning and Characterization of a Rhamnose-Binding Lectin SUL-I from the Toxopneustid Sea Urchin Toxopneustes Pileolus Venom. Toxicon 2015, 94, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, T.; Ichise, A.; Unno, H.; Goda, S.; Oda, T.; Tateno, H.; Hirabayashi, J.; Sakai, H.; Nakagawa, H. Carbohydrate Recognition by the Rhamnose-Binding Lectin SUL-I with a Novel Three-Domain Structure Isolated from the Venom of Globiferous Pedicellariae of the Flower Sea Urchin Toxopneustes Pileolus. Protein Sci. 2017, 26, 1574–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirai, T.; Watanabe, Y.; Lee, M.; Ogawa, T.; Muramoto, K. Structure of Rhamnose-Binding Lectin CSL3: Unique Pseudo-Tetrameric Architecture of a Pattern Recognition Protein. J. Mol. Biol. 2009, 391, 390–403. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Unno, H.; Kouzuma, Y.; Uchida, T.; Eto, S.; Hidemura, H.; Kato, N.; Yonekura, M.; Kusunoki, M. C-Type Lectin-like Carbohydrate Recognition of the Hemolytic Lectin CEL-III Containing Ricin-Type β-Trefoil Folds. J. Biol. Chem. 2007, 282, 37826–37835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, T.; Kohzaki, H.; Nagatomo, H.; Yamasaki, N. Purification and Characterization of Four Ca2+-Dependent Lectins from the Marine Invertebrate, Cucumaria Echinata. J. Biochem. 1994, 116, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Tabata, S.; Sugihara, K.; Kouzuma, Y.; Kimura, M.; Yamasaki, N. Primary Structure of Hemolytic Lectin CEL-III from Marine Invertebrate Cucumaria Echinata and Its CDNA: Structural Similarity to the B-Chain from Plant Lectin, Ricin. Biochim. Biophys. Acta—Protein Struct. Mol. Enzymol. 1999, 1435, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Unno, H.; Goda, S.; Hatakeyama, T. Hemolytic Lectin CEL-III Heptamerizes via a Large Structural Transition from α-Helices to a β-Barrel during the Transmembrane Pore Formation Process. J. Biol. Chem. 2014, 289, 12805–12812. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, T.; Furukawa, M.; Nagatomo, H.; Yamasaki, N.; Mori, T. Oligomerization of the Hemolytic Lectin CEL-III from the Marine Invertebrate Cucumaria Echinata Induced by the Binding of Carbohydrate Ligands. J. Biol. Chem. 1996, 271, 16915–16920. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, T.; Sato, T.; Taira, E.; Kuwahara, H.; Niidome, T.; Aoyagi, H. Characterization of the Interaction of Hemolytic Lectin CEL-III from the Marine Invertebrate, Cucumaria Echinata, with Artificial Lipid Membranes: Involvement of Neutral Sphingoglycolipids in the Pore-Forming Process. J. Biochem. 1999, 125, 277–284. [Google Scholar] [CrossRef]
- Hazes, B. The Qxw Motif: A Flexilble Lectin Fold. Protein Sci. 1996, 5, 1490–1501. [Google Scholar] [CrossRef] [Green Version]
- Mancheño, J.M.; Tateno, H.; Goldstein, I.J.; Martínez-Ripoll, M.; Hermoso, J.A. Structural Analysis of the Laetiporus Sulphureus Hemolytic Pore-Forming Lectin in Complex with Sugars. J. Biol. Chem. 2005, 280, 17251–17259. [Google Scholar] [CrossRef] [Green Version]
- Podobnik, M.; Kisovec, M.; Anderluh, G. Molecular Mechanism of Pore Formation by Aerolysin-like Proteins. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160209. [Google Scholar] [CrossRef] [Green Version]
- Rajan, B.; Patel, D.M.; Kitani, Y.; Viswanath, K.; Brinchmann, M.F. Novel Mannose Binding Natterin-like Protein in the Skin Mucus of Atlantic Cod (Gadus morhua). Fish Shellfish Immunol. 2017, 68, 452–457. [Google Scholar] [CrossRef]
- Lima, C.; Disner, G.R.; Alice, M.; Falc, P.; Seni-silva, A.C.; Luis, A.; Maleski, A.; Souza, M.M.; Cristina, M.; Tonello, R.; et al. The Natterin Proteins Diversity: A Review on Phylogeny, Structure, and Immune Function. Toxins 2021, 13, 538. [Google Scholar] [CrossRef]
- Jia, N.; Liu, N.; Cheng, W.; Jiang, Y.; Sun, H.; Chen, L.; Peng, J.; Zhang, Y.; Ding, Y.; Zhang, Z.; et al. Structural Basis for Receptor Recognition and Pore Formation of a Zebrafish Aerolysin-like Protein. EMBO Rep. 2016, 17, 235–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Rosado, C.J.; Kondos, S.; Bull, T.E.; Kuiper, M.J.; Law, R.H.P.; Buckle, A.M.; Voskoboinik, I.; Bird, P.I.; Trapani, J.A.; Whisstock, J.C.; et al. The MACPF/CDC Family of Pore-Forming Toxins. Cell. Microbiol. 2008, 10, 1765–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenabeele, P.; Bultynck, G.; Savvides, S.N. Pore-Forming Proteins as Drivers of Membrane Permeabilization in Cell Death Pathways. Nat. Rev. Mol. Cell Biol. 2023, 24, 312–333. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatakeyama, T.; Unno, H. Functional Diversity of Novel Lectins with Unique Structural Features in Marine Animals. Cells 2023, 12, 1814. https://doi.org/10.3390/cells12141814
Hatakeyama T, Unno H. Functional Diversity of Novel Lectins with Unique Structural Features in Marine Animals. Cells. 2023; 12(14):1814. https://doi.org/10.3390/cells12141814
Chicago/Turabian StyleHatakeyama, Tomomitsu, and Hideaki Unno. 2023. "Functional Diversity of Novel Lectins with Unique Structural Features in Marine Animals" Cells 12, no. 14: 1814. https://doi.org/10.3390/cells12141814