
Chromosome Territories in Hematological Malignancies


 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Nuclear Architecture: Organization from Nucleotides to Chromosome Territories
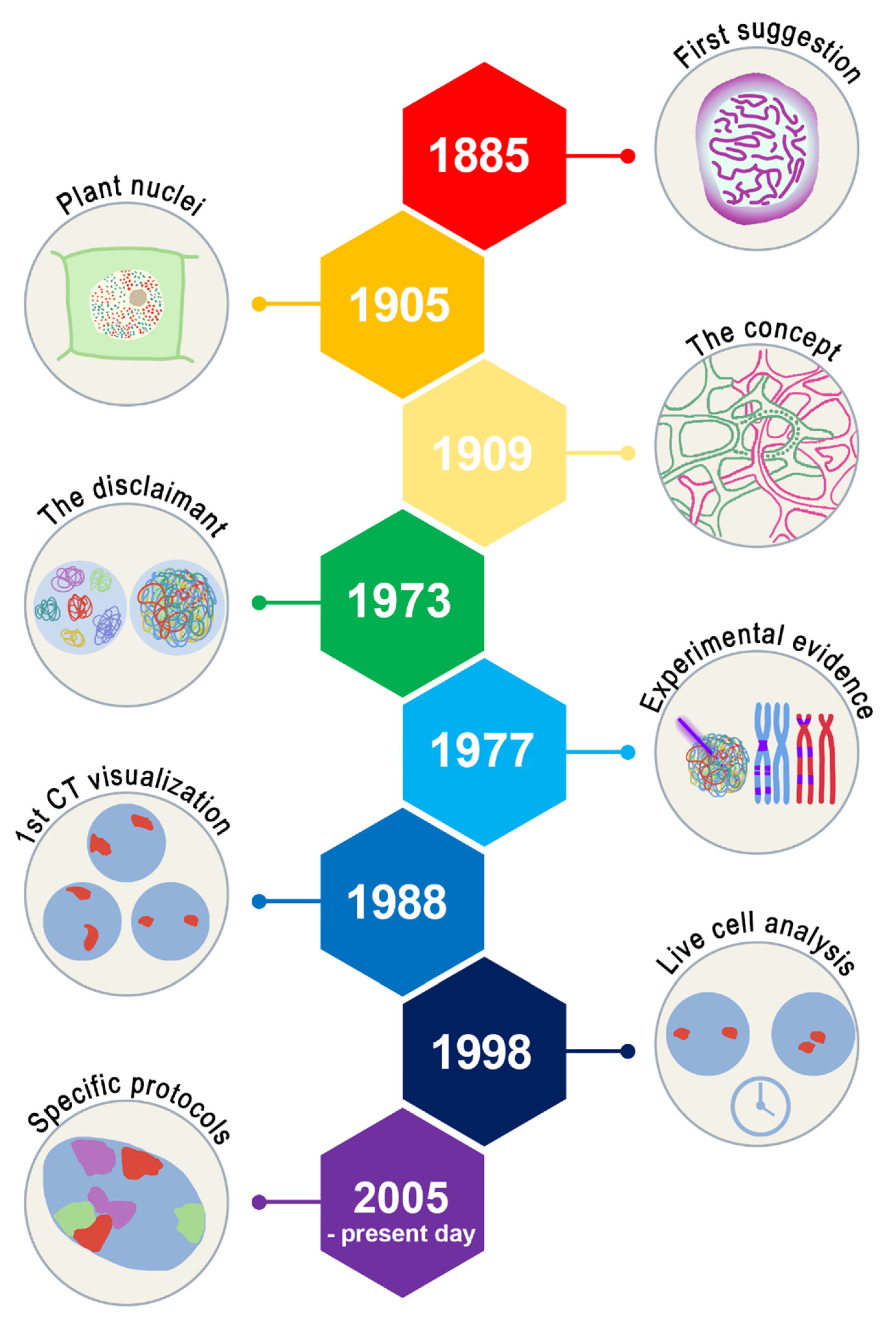
2. History of Chromosome Territories
2.1. Chromosome Territories: Stablished Concepts
2.2. Cell Differentiation and Changes in CT of Hematological Cells
2.3. Clinical Applications
3. Chromosome Territories in Hematological Cancers
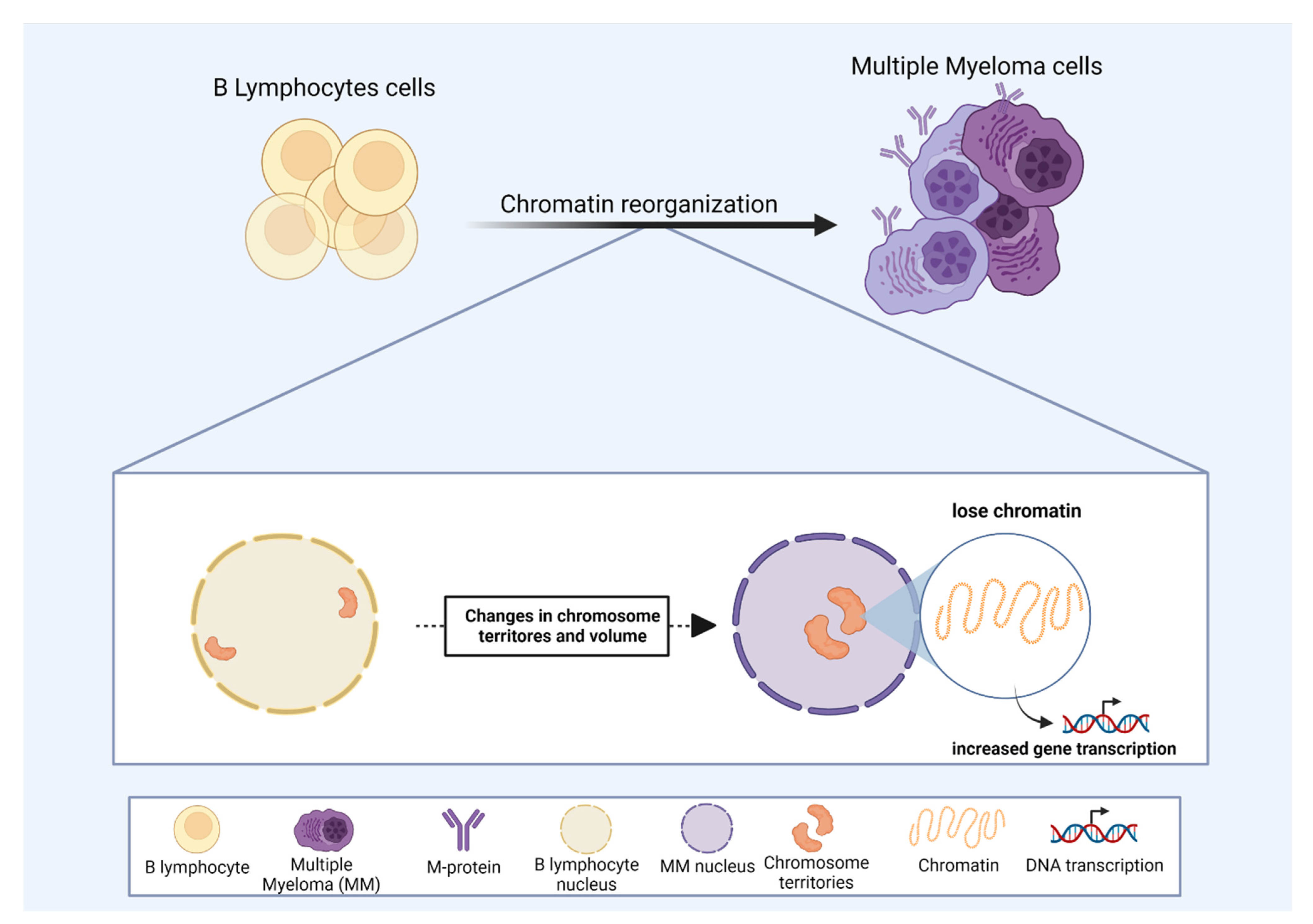
3.1. Multiple Myeloma
3.2. Acute Myeloid Leukemia and Secondary Leukemia
3.3. Radiation Effects on CT in Hematological Malignancies
3.4. Lymphomas
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berezney, R. Regulating the mammalian genome: The role of nuclear architecture. Adv. Enzym. Regul. 2002, 42, 39–52. [Google Scholar] [CrossRef]
- Chang, P.; Gohain, M.; Yen, M.-R.; Chen, P.-Y. Computational Methods for Assessing Chromatin Hierarchy. Comput. Struct. Biotechnol. J. 2018, 16, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Soler-Vila, P.; Cuscó, P.; Farabella, I.; Di Stefano, M.; Marti-Renom, M.A. Hierarchical chromatin organization detected by TADpole. Nucleic Acids Res. 2020, 48, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinchenko, A.; Berezhnoy, N.V.; Chen, Q.; Nordenskiöld, L. Compaction of Single-Molecule Megabase-Long Chromatin under the Influence of Macromolecular Crowding. Biophys. J. 2018, 114, 2326–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakovchuk, P. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res. 2006, 34, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Privalov, P.L.; Crane-Robinson, C. Forces maintaining the DNA double helix and its complexes with transcription factors. Prog. Biophys. Mol. Biol. 2018, 135, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Oudet, P.; Gross-Bellard, M.; Chambon, P. Electron microscopic and biochemical evidence that chromatin structure is a repeating unit. Cell 1975, 4, 281–300. [Google Scholar] [CrossRef]
- Belmont, A.S.; Bruce, K. Visualization of G1 chromosomes: A folded, twisted, supercoiled chromonema model of interphase chromatid structure. J. Cell Biol. 1994, 127, 287–302. [Google Scholar] [CrossRef]
- Adkins, N.L.; Watts, M.; Georgel, P.T. To the 30-nm chromatin fiber and beyond. Biochim. Biophys. Acta-Gene Struct. Expr. 2004, 1677, 12–23. [Google Scholar] [CrossRef]
- Maeshima, K.; Ide, S.; Babokhov, M. Dynamic chromatin organization without the 30-nm fiber. Curr. Opin. Cell Biol. 2019, 58, 95–104. [Google Scholar] [CrossRef]
- Kuznetsova, M.A.; Sheval, E.V. Chromatin fibers: From classical descriptions to modern interpretation. Cell Biol. Int. 2016, 40, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.P. Chromosome structure in relation to the chromosome cycle. Bot. Rev. 1936, 2, 529–553. [Google Scholar] [CrossRef]
- Sparvoli, E.; Gay, H.; Kaufmann, B.P. Number and pattern of association of chromonemata in the chromosomes of Tradescantia. Chromosoma 1965, 16, 415–435. [Google Scholar] [CrossRef] [Green Version]
- Baranetsky, J. Die Kerntheilung in den Pollenmutterzellen einiger Tradescantien. Bot. Zeit. 1880, 38, 241–248. [Google Scholar]
- Chu, L.; Liang, Z.; Mukhina, M.; Fisher, J.; Vincenten, N.; Zhang, Z.; Hutchinson, J.; Zickler, D.; Kleckner, N. The 3D Topography of Mitotic Chromosomes. Mol. Cell 2020, 79, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Münkel, C.; Eils, R.; Dietzel, S.; Zink, D.; Mehring, C.; Wedemann, G.; Cremer, T.; Langowski, J. Compartmentalization of Interphase Chromosomes Observed in Simulation and Experiment. J. Mol. Biol. 1999, 285, 1053–1065. [Google Scholar] [CrossRef] [Green Version]
- Sachs, R.K.; van den Engh, G.; Trask, B.; Yokota, H.; Hearst, J.E. A random-walk/giant-loop model for interphase chromosomes. Proc. Natl. Acad. Sci. USA 1995, 92, 2710–2714. [Google Scholar] [CrossRef] [Green Version]
- Yokota, H.; van den Engh, G.; Hearst, J.E.; Sachs, R.K.; Trask, B.J. Evidence for the organization of chromatin in megabase pair-sized loops arranged along a random walk path in the human G0/G1 interphase nucleus. J. Cell Biol. 1995, 130, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Hirota, T. Folding the genome into mitotic chromosomes. Curr. Opin. Cell Biol. 2019, 60, 19–26. [Google Scholar] [CrossRef]
- Hansen, A.S. CTCF as a boundary factor for cohesin-mediated loop extrusion: Evidence for a multi-step mechanism. Nucleus 2020, 11, 132–148. [Google Scholar] [CrossRef]
- Nuebler, J.; Fudenberg, G.; Imakaev, M.; Abdennur, N.; Mirny, L.A. Chromatin organization by an interplay of loop extrusion and compartmental segregation. Proc. Natl. Acad. Sci. USA 2018, 115, E6697–E6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Q.; Cheng, A.Y.; Gu, X.; Harada, B.T.; Yu, M.; Wu, T.; Ren, B.; Ouyang, Z.; He, C. Direct DNA crosslinking with CAP-C uncovers transcription-dependent chromatin organization at high resolution. Nat. Biotechnol. 2021, 39, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Rowley, M.J.; Lyu, X.; Rana, V.; Ando-Kuri, M.; Karns, R.; Bosco, G.; Corces, V.G. Condensin II Counteracts Cohesin and RNA Polymerase II in the Establishment of 3D Chromatin Organization. Cell Rep. 2019, 26, 2890–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.S.P.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [Green Version]
- Merkenschlager, M.; Nora, E.P. CTCF and Cohesin in Genome Folding and Transcriptional Gene Regulation. Annu. Rev. Genom. Hum. Genet. 2016, 17, 17–43. [Google Scholar] [CrossRef]
- Alipour, E.; Marko, J.F. Self-organization of domain structures by DNA-loop-extruding enzymes. Nucleic Acids Res. 2012, 40, 11202–11212. [Google Scholar] [CrossRef] [Green Version]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.F.; Bauer, B.; Goetz, D.; Tang, W.; Wutz, G.; Peters, J.-M. DNA loop extrusion by human cohesin. Science 2019, 366, 1338–1345. [Google Scholar] [CrossRef]
- Sanborn, A.L.; Rao, S.S.P.; Huang, S.-C.; Durand, N.C.; Huntley, M.H.; Jewett, A.I.; Bochkov, I.D.; Chinnappan, D.; Cutkosky, A.; Li, J.; et al. Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl. Acad. Sci. USA 2015, 112, E6456–E6465. [Google Scholar] [CrossRef] [Green Version]
- Mifsud, B.; Tavares-Cadete, F.; Young, A.N.; Sugar, R.; Schoenfelder, S.; Ferreira, L.; Wingett, S.W.; Andrews, S.; Grey, W.; Ewels, P.A.; et al. Mapping long-range promoter contacts in human cells with high-resolution capture Hi-C. Nat. Genet. 2015, 47, 598–606. [Google Scholar] [CrossRef]
- Schoenfelder, S.; Furlan-Magaril, M.; Mifsud, B.; Tavares-Cadete, F.; Sugar, R.; Javierre, B.-M.; Nagano, T.; Katsman, Y.; Sakthidevi, M.; Wingett, S.W.; et al. The pluripotent regulatory circuitry connecting promoters to their long-range interacting elements. Genome Res. 2015, 25, 582–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Abnousi, A.; Zhang, Y.; Li, G.; Lee, L.; Chen, Z.; Fang, R.; Lagler, T.M.; Yang, Y.; Wen, J.; et al. SnapHiC: A computational pipeline to identify chromatin loops from single-cell Hi-C data. Nat. Methods 2021, 18, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.R.; Hartl, T.A.; Bosco, G. Condensin II Promotes the Formation of Chromosome Territories by Inducing Axial Compaction of Polyploid Interphase Chromosomes. PLoS Genet. 2012, 8, e1002873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, L.C.; Kalitsis, P.; Chang, T.M.; Cipetic, M.; Kim, J.H.; Marshall, O.; Turnbull, L.; Whitchurch, C.B.; Vagnarelli, P.; Samejima, K.; et al. Contrasting roles of condensin I and II in mitotic chromosome formation. J. Cell Sci. 2012, 125, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Paulson, J.R.; Bakhrebah, M.; Kim, J.H.; Nowell, C.; Kalitsis, P.; Hudson, D.F. Condensin I and II behaviour in interphase nuclei and cells undergoing premature chromosome condensation. Chromosome Res. 2016, 24, 243–269. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, T.; Imai, R.; Tanbo, M.; Nagashima, R.; Tamura, S.; Tani, T.; Joti, Y.; Tomita, M.; Hibino, K.; Kanemaki, M.T.; et al. Dynamic Organization of Chromatin Domains Revealed by Super-Resolution Live-Cell Imaging. Mol. Cell 2017, 67, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Iashina, E.G.; Grigoriev, S.V. Large-Scale Structure of Chromatin: A Fractal Globule or a Logarithmic Fractal? J. Exp. Theor. Phys. 2019, 129, 455–458. [Google Scholar] [CrossRef]
- Shaban, H.A.; Barth, R.; Bystricky, K. Formation of correlated chromatin domains at nanoscale dynamic resolution during transcription. Nucleic Acids Res. 2018, 46, e77. [Google Scholar] [CrossRef] [Green Version]
- Cremer, T.; Cremer, M. Chromosome Territories. Cold Spring Harb. Perspect. Biol. 2010, 2, a003889. [Google Scholar] [CrossRef] [Green Version]
- Meaburn, K.J.; Misteli, T. Chromosome territories. Nature 2007, 445, 379–381. [Google Scholar] [CrossRef]
- Kinney, N.A.; Sharakhov, I.V.; Onufriev, A.V. Chromosome–nuclear envelope attachments affect interphase chromosome territories and entanglement. Epigenetics Chromatin 2018, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Ginno, P.A.; Burger, L.; Seebacher, J.; Iesmantavicius, V.; Schübeler, D. Cell cycle-resolved chromatin proteomics reveals the extent of mitotic preservation of the genomic regulatory landscape. Nat. Commun. 2018, 9, 4048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramo, K.; Valton, A.-L.; Venev, S.V.; Ozadam, H.; Fox, A.N.; Dekker, J. A chromosome folding intermediate at the condensin-to-cohesin transition during telophase. Nat. Cell Biol. 2019, 21, 1393–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrate, Z.; Solé, M.; Vidal, F.; Anton, E.; Blanco, J. Chromosome positioning and male infertility: It comes with the territory. J. Assist. Reprod. Genet. 2018, 35, 1929–1938. [Google Scholar] [CrossRef]
- Champroux, A.; Goubely, C.; Henry-Berger, J.; Drevet, J.R.; Kocer, A. Three-Dimensional Confocal Analysis of Chromosome Positioning Coupled with Immunofluorescence in Mouse Sperm Nuclei. In DNA Modifications; Humana: New York, NY, USA, 2021; pp. 255–268. [Google Scholar]
- de Castro, I.J.; Toner, B.; Xie, S.Q.; Swingland, J.; Hodges, A.; Tabrizi, S.J.; Turkheimer, F.; Pombo, A.; Khalil, A. Altered nuclear architecture in blood cells from Huntington’s disease patients. Neurol. Sci. 2021, 43, 379–385. [Google Scholar] [CrossRef]
- Rabl, C. Uber Zelltheilung; W. Engelmann: Leipzig, Germany, 1885; Volume 10, pp. 214–330. [Google Scholar]
- Boveri, T. Die Blastomerenkerne von Ascaris Megalocephala und die Theorie der Chromosomenindividualität; Arch Zellforsch: Leipzig, Germany, 1909; Volume 3, pp. 181–182. [Google Scholar]
- Strasburger, E. “Die” stofflichen Grundlagen der Vererbung im organischen Reich: Versuche einer gemeinverständlichen Darstellung; Fischer: Ried im Innkreis, Austria, 1905. [Google Scholar]
- Bolzer, A.; Kreth, G.; Solovei, I.; Koehler, D.; Saracoglu, K.; Fauth, C.; Müller, S.; Eils, R.; Cremer, C.; Speicher, M.R.; et al. Three-Dimensional Maps of All Chromosomes in Human Male Fibroblast Nuclei and Prometaphase Rosettes. PLoS Biol. 2005, 3, e157. [Google Scholar] [CrossRef] [Green Version]
- Wischnitzer, S. The Submicroscopic Morphology of the Interphase Nucleus. Int. Rev. Cytol. 1973, 34, 1–48. [Google Scholar]
- Stack, S.M.; Brown, D.B.; Dewey, W.C. Visualization of interphase chromosomes. J. Cell Sci. 1977, 26, 281–299. [Google Scholar] [CrossRef]
- Zorn, C.; Cremer, C.; Cremer, T.; Zimmer, J. Unscheduled DNA synthesis after partial UV irradiation of the cell nucleus. Exp. Cell Res. 1979, 124, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Cremer, T.; Baumann, H.; Nakanishi, K.; Cremer, C. Correlation between interphase and metaphase chromosome arrangements as studied by laser-uv-microbeam experiments. In Chromosomes Today; Springer: Dordrecht, The Netherlands, 1984; pp. 203–212. [Google Scholar]
- Lichter, P.; Cremer, T.; Borden, J.; Manuelidis, L.; Ward, D.C. Delineation of individual human chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries. Hum. Genet. 1988, 80, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Manuelidis, L. Individual interphase chromosome domains revealed by in situ hybridization. Hum. Genet. 1985, 71, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Zink, D.; Cremer, T. Cell nucleus: Chromosome dynamics in nuclei of living cells. Curr. Biol. 1998, 8, R321–R324. [Google Scholar] [CrossRef] [Green Version]
- Zink, D.; Cremer, T.; Saffrich, R.; Fischer, R.; Trendelenburg, M.F.; Ansorge, W.; Stelzer, E.H.K. Structure and dynamics of human interphase chromosome territories in vivo. Hum. Genet. 1998, 102, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Gasser, S.M. Visualizing Chromatin Dynamics in Interphase Nuclei. Science 2002, 296, 1412–1416. [Google Scholar] [CrossRef]
- Pliss, A.; Malyavantham, K.; Bhattacharya, S.; Zeitz, M.; Berezney, R. Chromatin dynamics is correlated with replication timing. Chromosoma 2009, 118, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Pliss, A.; Malyavantham, K.S.; Bhattacharya, S.; Berezney, R. Chromatin dynamics in living cells: Identification of oscillatory motion. J. Cell. Physiol. 2013, 228, 609–616. [Google Scholar] [CrossRef]
- Cremer, M.; Grasser, F.; Lanctôt, C.; Müller, S.; Neusser, M.; Zinner, R.; Solovei, I.; Cremer, T. Multicolor 3D Fluorescence In Situ Hybridization for Imaging Interphase Chromosomes. In The Nucleus; Humana Press: Totowa, NJ, USA, 2012; pp. 205–239. [Google Scholar]
- Tanabe, H.; Habermann, F.A.; Solovei, I.; Cremer, M.; Cremer, T. Non-random radial arrangements of interphase chromosome territories: Evolutionary considerations and functional implications. Mutat. Res. Mol. Mech. Mutagen. 2002, 504, 37–45. [Google Scholar] [CrossRef]
- Mayer, R.; Brero, A.; von Hase, J.; Schroeder, T.; Cremer, T.; Dietzel, S. Common themes and cell type specific variations of higher order chromatin arrangements in the mouse. BMC Cell Biol. 2005, 6, 44. [Google Scholar] [CrossRef] [Green Version]
- Khalil, A.; Grant, J.L.; Caddle, L.B.; Atzema, E.; Mills, K.D.; Arneodo, A. Chromosome territories have a highly nonspherical morphology and nonrandom positioning. Chromosome Res. 2007, 15, 899–916. [Google Scholar] [CrossRef]
- Fritz, A.J.; Stojkovic, B.; Ding, H.; Xu, J.; Bhattacharya, S.; Gaile, D.; Berezney, R. Wide-scale alterations in interchromosomal organization in breast cancer cells: Defining a network of interacting chromosomes. Hum. Mol. Genet. 2014, 23, 5133–5146. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, H.; Küpper, K.; Ishida, T.; Neusser, M.; Mizusawa, H. Inter- and intra-specific gene-density-correlated radial chromosome territory arrangements are conserved in Old World monkeys. Cytogenet. Genome Res. 2005, 108, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahy, N.L.; Perry, P.E.; Bickmore, W.A. Gene density and transcription influence the localization of chromatin outside of chromosome territories detectable by FISH. J. Cell Biol. 2002, 159, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Kreth, G.; Finsterle, J.; von Hase, J.; Cremer, M.; Cremer, C. Radial Arrangement of Chromosome Territories in Human Cell Nuclei: A Computer Model Approach Based on Gene Density Indicates a Probabilistic Global Positioning Code. Biophys. J. 2004, 86, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Malyavantham, K.S.; Bhattacharya, S.; Alonso, W.D.; Acharya, R.; Berezney, R. Spatio-temporal dynamics of replication and transcription sites in the mammalian cell nucleus. Chromosoma 2008, 117, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Malyavantham, K.S.; Bhattacharya, S.; Barbeitos, M.; Mukherjee, L.; Xu, J.; Fackelmayer, F.O.; Berezney, R. Identifying functional neighborhoods within the cell nucleus: Proximity analysis of early S-phase replicating chromatin domains to sites of transcription, RNA polymerase II, HP1γ, matrin 3 and SAF-A. J. Cell. Biochem. 2008, 105, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Neusser, M.; Schubel, V.; Koch, A.; Cremer, T.; Müller, S. Evolutionarily conserved, cell type and species-specific higher order chromatin arrangements in interphase nuclei of primates. Chromosoma 2007, 116, 307–320. [Google Scholar] [CrossRef]
- Han, Z.; Cui, K.; Placek, K.; Hong, N.; Lin, C.; Chen, W.; Zhao, K.; Jin, W. Diploid genome architecture revealed by multi-omic data of hybrid mice. Genome Res. 2020, 30, 1097–1106. [Google Scholar] [CrossRef]
- Brickner, J. Genetic and epigenetic control of the spatial organization of the genome. Mol. Biol. Cell 2017, 28, 364–369. [Google Scholar] [CrossRef] [Green Version]
- Baroux, C. Three-dimensional genome organization in epigenetic regulations: Cause or consequence? Curr. Opin. Plant Biol. 2021, 61, 102031. [Google Scholar] [CrossRef]
- Kurz, A.; Lampel, S.; Nickolenko, J.E.; Bradl, J.; Benner, A.; Zirbel, R.M.; Cremer, T.; Lichter, P. Active and inactive genes localize preferentially in the periphery of chromosome territories. J. Cell Biol. 1996, 135, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.R.E. Transcription and the territory: The ins and outs of gene positioning. Trends Genet. 2003, 19, 298–302. [Google Scholar] [CrossRef]
- Wegel, E.; Shaw, P. Gene activation and deactivation related changes in the three-dimensional structure of chromatin. Chromosoma 2005, 114, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.L.; Zullo, J.M.; Bertolino, E.; Singh, H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature 2008, 452, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Peric-Hupkes, D.; van Steensel, B. Role of the Nuclear Lamina in Genome Organization and Gene Expression. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shevelyov, Y.Y.; Ulianov, S.V. Role of Nuclear Lamina in Gene Repression and Maintenance of Chromosome Architecture in the Nucleus. Biochem. 2018, 83, 359–369. [Google Scholar] [CrossRef]
- Shevelyov, Y.Y.; Ulianov, S.V. The Nuclear Lamina as an Organizer of Chromosome Architecture. Cells 2019, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Clowney, E.J.; LeGros, M.A.; Mosley, C.P.; Clowney, F.G.; Markenskoff-Papadimitriou, E.C.; Myllys, M.; Barnea, G.; Larabell, C.A.; Lomvardas, S. Nuclear Aggregation of Olfactory Receptor Genes Governs Their Monogenic Expression. Cell 2012, 151, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Armelin-Correa, L.M.; Malnic, B. Capturing genome folds in single sensory neurons. Nat. Struct. Mol. Biol. 2019, 26, 254–255. [Google Scholar] [CrossRef]
- Armelin-Correa, L.M.; Nagai, M.H.; Silva, A.G.L.; Malnic, B. Nuclear architecture and gene silencing in olfactory sensory neurons. Bioarchitecture 2014, 4, 160–163. [Google Scholar] [CrossRef] [Green Version]
- Solovei, I.; Kreysing, M.; Lanctôt, C.; Kösem, S.; Peichl, L.; Cremer, T.; Guck, J.; Joffe, B. Nuclear Architecture of Rod Photoreceptor Cells Adapts to Vision in Mammalian Evolution. Cell 2009, 137, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.B.; Shen, J.; Yokota, H. Size-Dependent Positioning of Human Chromosomes in Interphase Nuclei. Biophys. J. 2000, 79, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Pliss, A.; Fritz, A.J.; Stojkovic, B.; Ding, H.; Mukherjee, L.; Bhattacharya, S.; Xu, J.; Berezney, R. Non-Random Patterns in the Distribution of NOR-Bearing Chromosome Territories in Human Fibroblasts: A Network Model of Interactions. J. Cell. Physiol. 2015, 230, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitz, M.J.; Marella, N.V.; Malyavantham, K.S.; Goetze, S.; Bode, J.; Raska, I.; Berezney, R. Organization of the amplified type I interferon gene cluster and associated chromosome regions in the interphase nucleus of human osteosarcoma cells. Chromosome Res. 2009, 17, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Foster, H.A.; Griffin, D.K.; Bridger, J.M. Interphase chromosome positioning in in vitro porcine cells and ex vivo porcine tissues. BMC Cell Biol. 2012, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Kemeny, S.; Tatout, C.; Salaun, G.; Pebrel-Richard, C.; Goumy, C.; Ollier, N.; Maurin, E.; Pereira, B.; Vago, P.; Gouas, L. Spatial organization of chromosome territories in the interphase nucleus of trisomy 21 cells. Chromosoma 2018, 127, 247–259. [Google Scholar] [CrossRef]
- Kim, J.-H.; Noskov, V.N.; Ogurtsov, A.Y.; Nagaraja, R.; Petrov, N.; Liskovykh, M.; Walenz, B.P.; Lee, H.-S.; Kouprina, N.; Phillippy, A.M.; et al. The genomic structure of a human chromosome 22 nucleolar organizer region determined by TAR cloning. Sci. Rep. 2021, 11, 2997. [Google Scholar] [CrossRef]
- Mangan, H.; McStay, B. Human nucleoli comprise multiple constrained territories, tethered to individual chromosomes. Genes Dev. 2021, 35, 483–488. [Google Scholar] [CrossRef]
- van Sluis, M.; van Vuuren, C.; Mangan, H.; McStay, B. NORs on human acrocentric chromosome p-arms are active by default and can associate with nucleoli independently of rDNA. Proc. Natl. Acad. Sci. USA 2020, 117, 10368–10377. [Google Scholar] [CrossRef] [Green Version]
- L’Abbate, A.; Tolomeo, D.; De Astis, F.; Lonoce, A.; Cunsolo, C.L.; Mühlematter, D.; Schoumans, J.; Vandenberghe, P.; Van Hoof, A.; Palumbo, O.; et al. t(15;21) translocations leading to the concurrent downregulation of RUNX1 and its transcription factor partner genes SIN3A and TCF12 in myeloid disorders. Mol. Cancer 2015, 14, 211. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Tanabe, H.; Yoshida, K.; Oikawa, K.; Saito, A.; Kiyuna, T.; Mizusawa, H.; Mukai, K. Alteration of chromosome positioning during adipocyte differentiation. J. Cell Sci. 2004, 117, 5897–5903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, N.; Seifert, B.; Ding, H.; Chen, Z.; Stojkovic, B.; Bhattacharya, S.; Xu, J.; Berezney, R. Reorganization of the interchromosomal network during keratinocyte differentiation. Chromosoma 2016, 125, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Mehta, I.S.; Amira, M.; Harvey, A.J.; Bridger, J.M. Rapid chromosome territory relocation by nuclear motor activity in response to serum removal in primary human fibroblasts. Genome Biol. 2010, 11, R5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, O.; Corcoran, C.J.; Noma, K. Involvement of condensin-directed gene associations in the organization and regulation of chromosome territories during the cell cycle. Nucleic Acids Res. 2016, 44, 3618–3628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada, L.A.; McQueen, P.G.; Misteli, T. Tissue-specific spatial organization of genomes. Genome Biol. 2004, 5, R44. [Google Scholar] [CrossRef] [Green Version]
- Bártová, E.; Kozubek, S.; Kozubek, M.; Jirsová, P.; Lukášová, E.; Skalníková, M.; Buchníčková, K. The influence of the cell cycle, differentiation and irradiation on the nuclear location of the abl, bcr and c-myc genes in human leukemic cells. Leuk. Res. 2000, 24, 233–241. [Google Scholar] [CrossRef]
- Kosak, S.T. Subnuclear Compartmentalization of Immunoglobulin Loci During Lymphocyte Development. Science 2002, 296, 158–162. [Google Scholar] [CrossRef]
- Lomiento, M.; Mammoli, F.; Mazza, E.M.C.; Bicciato, S.; Ferrari, S. Chromosome positioning in interphase nuclei of hematopoietic stem cell and myeloid precursor. Hematol. Rep. 2018, 10, 7515. [Google Scholar] [CrossRef] [Green Version]
- Hübner, B.; Lomiento, M.; Mammoli, F.; Illner, D.; Markaki, Y.; Ferrari, S.; Cremer, M.; Cremer, T. Remodeling of nuclear landscapes during human myelopoietic cell differentiation maintains co-aligned active and inactive nuclear compartments. Epigenetics Chromatin 2015, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; McQueen, P.G.; Lichtman, M.K.; Shevach, E.M.; Parada, L.A.; Misteli, T. Spatial genome organization during T-cell differentiation. Cytogenet. Genome Res. 2004, 105, 292–301. [Google Scholar] [CrossRef]
- Alcobia, I.; Dilão, R.; Parreira, L. Spatial associations of centromeres in the nuclei of hematopoietic cells: Evidence for cell-type-specific organizational patterns. Blood 2000, 95, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Chaly, N.; Munro, S.B. Centromeres Reposition to the Nuclear Periphery during L6E9 Myogenesisin Vitro. Exp. Cell Res. 1996, 223, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Rahemtullah, A.; Reichard, K.K.; Preffer, F.I.; Harris, N.L.; Hasserjian, R.P. A Double-Positive CD4+CD8+ T-Cell Population Is Commonly Found in Nodular Lymphocyte Predominant Hodgkin Lymphoma. Am. J. Clin. Pathol. 2006, 126, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Alshaker, H.; Mills, R.; Hunter, E.; Salter, M.; Ramadass, A.; Skinner, B.M.; Westra, W.; Green, J.; Akoulitchev, A.; Winkler, M.; et al. Chromatin conformation changes in peripheral blood can detect prostate cancer and stratify disease risk groups. J. Transl. Med. 2021, 19, 46. [Google Scholar] [CrossRef] [PubMed]
- Zalensky, A.; Zalenskaya, I. Organization of chromosomes in spermatozoa: An additional layer of epigenetic information? Biochem. Soc. Trans. 2007, 35, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Finch, K.A.; Fonseka, K.G.L.; Abogrein, A.; Ioannou, D.; Handyside, A.H.; Thornhill, A.R.; Hickson, N.; Griffin, D.K. Nuclear organization in human sperm: Preliminary evidence for altered sex chromosome centromere position in infertile males. Hum. Reprod. 2008, 23, 1263–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, K.E.; Mandawala, A.A.; Griffin, D.K. The role of chromosome segregation and nuclear organisation in human subfertility. Biochem. Soc. Trans. 2019, 47, 425–432. [Google Scholar] [CrossRef]
- Marella, N.V.; Bhattacharya, S.; Mukherjee, L.; Xu, J.; Berezney, R. Cell type specific chromosome territory organization in the interphase nucleus of normal and cancer cells. J. Cell. Physiol. 2009, 221, 130–138. [Google Scholar] [CrossRef]
- Parl, F.F.; Dupont, W.D.; Crooke, P.S. Interchromosomal Translocations as a Means to Map Chromosome Territories in Breast Cancer. Cancer Inform. 2019, 18, 117693511984257. [Google Scholar] [CrossRef]
- Feng, L.; Li, H.; Zhang, Y.; Shen, H.; Yu, H.; Zhong, F.; Qin, L.; He, F.; Yang, P.; Tang, Z. Metastasis-related genes in hepatocellular carcinoma cell-lines are clustered on chromosome territories predicted by transcriptome and proteome. Sci. China Chem. 2016, 59, 380–382. [Google Scholar] [CrossRef]
- Branco, M.R.; Pombo, A. Intermingling of Chromosome Territories in Interphase Suggests Role in Translocations and Transcription-Dependent Associations. PLoS Biol. 2006, 4, e138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brianna Caddle, L.; Grant, J.L.; Szatkiewicz, J.; van Hase, J.; Shirley, B.-J.; Bewersdorf, J.; Cremer, C.; Arneodo, A.; Khalil, A.; Mills, K.D. Chromosome neighborhood composition determines translocation outcomes after exposure to high-dose radiation in primary cells. Chromosome Res. 2007, 15, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.A.; Resch, W.; Jankovic, M.; Oliveira, T.; Yamane, A.; Nakahashi, H.; Di Virgilio, M.; Bothmer, A.; Nussenzweig, A.; Robbiani, D.F.; et al. Translocation-Capture Sequencing Reveals the Extent and Nature of Chromosomal Rearrangements in B Lymphocytes. Cell 2011, 147, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roix, J.J.; McQueen, P.G.; Munson, P.J.; Parada, L.A.; Misteli, T. Spatial proximity of translocation-prone gene loci in human lymphomas. Nat. Genet. 2003, 34, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Pozzo, A.; Sisdelli, L.; Cordioli, M.I.V.; Vaisman, F.; Caria, P.; Mai, S.; Cerutti, J.M. Genetic Landscape of Papillary Thyroid Carcinoma and Nuclear Architecture: An Overview Comparing Pediatric and Adult Populations. Cancers 2020, 12, 3146. [Google Scholar] [CrossRef] [PubMed]
- Wiech, T.; Stein, S.; Lachenmaier, V.; Schmitt, E.; Schwarz-Finsterle, J.; Wiech, E.; Hildenbrand, G.; Werner, M.; Hausmann, M. Spatial allelic imbalance of BCL2 genes and chromosome 18 territories in nonneoplastic and neoplastic cervical squamous epithelium. Eur. Biophys. J. 2009, 38, 793–806. [Google Scholar] [CrossRef]
- Schwaederle, M.; Chattopadhyay, R.; Kato, S.; Fanta, P.T.; Banks, K.C.; Choi, I.S.; Piccioni, D.E.; Ikeda, S.; Talasaz, A.; Lanman, R.B.; et al. Genomic Alterations in Circulating Tumor DNA from Diverse Cancer Patients Identified by Next-Generation Sequencing. Cancer Res. 2017, 77, 5419–5427. [Google Scholar] [CrossRef] [Green Version]
- Knijnenburg, T.A.; Wang, L.; Zimmermann, M.T.; Chambwe, N.; Gao, G.F.; Cherniack, A.D.; Fan, H.; Shen, H.; Way, G.P.; Greene, C.S.; et al. Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep. 2018, 23, 239–254. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, S.R.; Ha, G.; Hoff, A.M.; Wala, J.A.; Carrot-Zhang, J.; Whelan, C.W.; Haradhvala, N.J.; Freeman, S.S.; Reed, S.C.; Rhoades, J.; et al. Structural Alterations Driving Castration-Resistant Prostate Cancer Revealed by Linked-Read Genome Sequencing. Cell 2018, 174, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Bridger, J.M.; Arican-Gotkas, H.D.; Foster, H.A.; Godwin, L.S.; Harvey, A.; Kill, I.R.; Knight, M.; Mehta, I.S.; Ahmed, M.H. The Non-random Repositioning of Whole Chromosomes and Individual Gene Loci in Interphase Nuclei and Its Relevance in Disease, Infection, Aging, and Cancer. Cancer Biol. Nucl. Envel. 2014, 773, 263–279. [Google Scholar]
- Vargas-Rondón, N.; Villegas, V.; Rondón-Lagos, M. The Role of Chromosomal Instability in Cancer and Therapeutic Responses. Cancers 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansregret, L.; Vanhaesebroeck, B.; Swanton, C. Determinants and clinical implications of chromosomal instability in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, H.P.; Barbash, O.; Creasy, C.L. Targeting epigenetic modifications in cancer therapy: Erasing the roadmap to cancer. Nat. Med. 2019, 25, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, M.A.; Terpos, E. Multiple myeloma. Ann. Oncol. 2010, 21, vii143–vii150. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Anderson, K. Multiple Myeloma. N. Engl. J. Med. 2011, 364, 1046–1060. [Google Scholar] [CrossRef] [Green Version]
- Padala, S.A.; Barsouk, A.; Barsouk, A.; Rawla, P.; Vakiti, A.; Kolhe, R.; Kota, V.; Ajebo, G.H. Epidemiology, Staging, and Management of Multiple Myeloma. Med. Sci. 2021, 9, 3. [Google Scholar] [CrossRef]
- Hideshima, T.; Mitsiades, C.; Tonon, G.; Richardson, P.G.; Anderson, K.C. Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nat. Rev. Cancer 2007, 7, 585–598. [Google Scholar] [CrossRef]
- Abdallah, N.; Rajkumar, S.V.; Greipp, P.; Kapoor, P.; Gertz, M.A.; Dispenzieri, A.; Baughn, L.B.; Lacy, M.Q.; Hayman, S.R.; Buadi, F.K.; et al. Cytogenetic abnormalities in multiple myeloma: Association with disease characteristics and treatment response. Blood Cancer J. 2020, 10, 82. [Google Scholar] [CrossRef]
- Su, C.T.; Chen, L.; Chen, J.; Parkin, B.; Polk, A.; Kandarpa, M.; Cole, C.E.; Campagnaro, E.; Vo, J.; Robinson, D.; et al. Role of Aneuploidy in Transcriptional Regulation and Clinical Prognosis in Relapsed and/or Refractory Multiple Myeloma (RRMM). Blood 2020, 136, 45–46. [Google Scholar] [CrossRef]
- Barwick, B.G.; Neri, P.; Bahlis, N.J.; Nooka, A.K.; Dhodapkar, M.V.; Jaye, D.L.; Hofmeister, C.C.; Kaufman, J.L.; Gupta, V.A.; Auclair, D.; et al. Multiple myeloma immunoglobulin lambda translocations portend poor prognosis. Nat. Commun. 2019, 10, 1911. [Google Scholar] [CrossRef] [Green Version]
- Morgan, G.J.; Walker, B.A.; Davies, F.E. The genetic architecture of multiple myeloma. Nat. Rev. Cancer 2012, 12, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Hakim, O.; Resch, W.; Yamane, A.; Klein, I.; Kieffer-Kwon, K.-R.; Jankovic, M.; Oliveira, T.; Bothmer, A.; Voss, T.C.; Ansarah-Sobrinho, C.; et al. DNA damage defines sites of recurrent chromosomal translocations in B lymphocytes. Nature 2012, 484, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Balajee, A.S.; Sanders, J.T.; Golloshi, R.; Shuryak, I.; McCord, R.P.; Dainiak, N. Investigation of Spatial Organization of Chromosome Territories in Chromosome Exchange Aberrations after Ionizing Radiation Exposure. Health Phys. 2018, 115, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Righolt, C.H.; Wiener, F.; Taylor-Kashton, C.; Harizanova, J.; Vermolen, B.J.; Garini, Y.; Young, I.T.; Mai, S. Translocation frequencies and chromosomal proximities for selected mouse chromosomes in primary B lymphocytes. Cytom. Part A 2011, 79, 276–283. [Google Scholar] [CrossRef]
- Wang, Y.; Nagarajan, M.; Uhler, C.; Shivashankar, G.V. Orientation and repositioning of chromosomes correlate with cell geometry–dependent gene expression. Mol. Biol. Cell 2017, 28, 1997–2009. [Google Scholar] [CrossRef]
- Schwope, R.; Magris, G.; Miculan, M.; Paparelli, E.; Celii, M.; Tocci, A.; Marroni, F.; Fornasiero, A.; De Paoli, E.; Morgante, M. Open chromatin in grapevine marks candidate CREs and with other chromatin features correlates with gene expression. Plant J. 2021, 107, 1631–1647. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, K.; De Paepe, A.; Hellqvist, E.; Krstic, A.D.; Metang, L.; Gustafsson, C.; Davis, R.E.; Levy, Y.M.; Surapaneni, R.; et al. Active enhancer and chromatin accessibility landscapes chart the regulatory network of primary multiple myeloma. Blood 2018, 131, 2138–2150. [Google Scholar] [CrossRef] [Green Version]
- Ordoñez, R.; Kulis, M.; Russiñol, N.; Chapaprieta, V.; Carrasco-Leon, A.; García-Torre, B.; Charalampopoulou, S.; Clot, G.; Beekman, R.; Meydan, C.; et al. Chromatin activation as a unifying principle underlying pathogenic mechanisms in multiple myeloma. Genome Res. 2020, 30, 1217–1227. [Google Scholar] [CrossRef]
- Sathitruangsak, C.; Righolt, C.H.; Klewes, L.; Tung Chang, D.; Kotb, R.; Mai, S. Distinct and shared three-dimensional chromosome organization patterns in lymphocytes, monoclonal gammopathy of undetermined significance and multiple myeloma. Int. J. Cancer 2017, 140, 400–410. [Google Scholar] [CrossRef]
- Sathitruangsak, C.; Righolt, C.H.; Klewes, L.; Tammur, P.; Ilus, T.; Tamm, A.; Punab, M.; Olujohungbe, A.; Mai, S. Quantitative Superresolution Microscopy Reveals Differences in Nuclear DNA Organization of Multiple Myeloma and Monoclonal Gammopathy of Undetermined Significance. J. Cell. Biochem. 2015, 116, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Broyl, A.; Hose, D.; Lokhorst, H.; de Knegt, Y.; Peeters, J.; Jauch, A.; Bertsch, U.; Buijs, A.; Stevens-Kroef, M.; Beverloo, H.B.; et al. Gene expression profiling for molecular classification of multiple myeloma in newly diagnosed patients. Blood 2010, 116, 2543–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Valent, E.T.; Beers, E.H.; Kuiper, R.; Oliva, S.; Haferlach, T.; Chng, W.; Vliet, M.H.; Sonneveld, P.; Larocca, A. Prognostic gene expression analysis in a retrospective, multinational cohort of 155 multiple myeloma patients treated outside clinical trials. Int. J. Lab. Hematol. 2021, 44, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ezoe, S. Secondary Leukemia Associated with the Anti-Cancer Agent, Etoposide, a Topoisomerase II Inhibitor. Int. J. Environ. Res. Public Health 2012, 9, 2444–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reikvam, H.; Hatfield, K.J.; Kittang, A.O.; Hovland, R.; Bruserud, Ø. Acute Myeloid Leukemia with the t(8;21) Translocation: Clinical Consequences and Biological Implications. J. Biomed. Biotechnol. 2011, 2011, 104631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubtsov, M.A.; Terekhov, S.M.; Razin, S.V.; Iarovaia, O.V. Repositioning of ETO gene in cells treated with VP-16, an inhibitor of DNA-Topoisomerase II. J. Cell. Biochem. 2008, 104, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, M.A.; Glukhov, S.I.; Allinne, J.; Pichugin, A.; Vassetzky, Y.S.; Razin, S.V.; Iarovaia, O.V. Treatment of lymphoid cells with the topoisomerase II poison etoposide leads to an increased juxtaposition of AML1 and ETO genes on the surface of nucleoli. Biopolym. Cell 2011, 27, 398–403. [Google Scholar] [CrossRef]
- Gole, B.; Wiesmüller, L. Leukemogenic rearrangements at the mixed lineage leukemia gene (MLL)—Multiple rather than a single mechanism. Front. Cell Dev. Biol. 2015, 3, 41. [Google Scholar] [CrossRef]
- Super, H.G.; Strissel, P.L.; Sobulo, O.M.; Burian, D.; Reshmi, S.C.; Roe, B.; Zeleznik-Le, N.J.; Diaz, M.O.; Rowley, J.D. Identification of complex genomic breakpoint junctions in the t(9;11) MLL-AF9 fusion gene in acute leukemia. Genes Chromosomes Cancer 1997, 20, 185–195. [Google Scholar] [CrossRef]
- Zhang, Y.; Strissel, P.; Strick, R.; Chen, J.; Nucifora, G.; Le Beau, M.M.; Larson, R.A.; Rowley, J.D. Genomic DNA breakpoints in AML1/RUNX1 and ETO cluster with topoisomerase II DNA cleavage and DNase I hypersensitive sites in t(8;21) leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 3070–3075. [Google Scholar] [CrossRef] [Green Version]
- Glukhov, S.I.; Rubtsov, M.A.; Alexeyevsky, D.A.; Alexeevski, A.V.; Razin, S.V.; Iarovaia, O.V. The Broken MLL Gene Is Frequently Located Outside the Inherent Chromosome Territory in Human Lymphoid Cells Treated with DNA Topoisomerase II Poison Etoposide. PLoS ONE 2013, 8, e75871. [Google Scholar] [CrossRef] [Green Version]
- Huret, J.; Dessen, P.; Bernheim, A. An Atlas on Chromosomes in Hematological Malignancies. Example: 11q23 and MLL partners. Leukemia 2001, 15, 987–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huret, J.L.; Senon, S.; Bernheim, A.; Dessen, P. An Atlas on genes and chromosomes in oncology and haematology. Cell. Mol. Biol. (Noisy-Le-Grand) 2004, 50, 805–807. [Google Scholar] [PubMed]
- Gué, M.; Sun, J.-S.; Boudier, T. Simultaneous localization of MLL, AF4 and ENL genes in interphase nuclei by 3D-FISH: MLL translocation revisited. BMC Cancer 2006, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutoglou, E.; Dorn, J.F.; Sengupta, K.; Jasin, M.; Nussenzweig, A.; Ried, T.; Danuser, G.; Misteli, T. Positional stability of single double-strand breaks in mammalian cells. Nat. Cell Biol. 2007, 9, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Aten, J.A. Dynamics of DNA Double-Strand Breaks Revealed by Clustering of Damaged Chromosome Domains. Science 2004, 303, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Nikiforova, M.N.; Stringer, J.R.; Blough, R.; Medvedovic, M.; Fagin, J.A.; Nikiforov, Y.E. Proximity of Chromosomal Loci That Participate in Radiation-Induced Rearrangements in Human Cells. Science 2000, 290, 138–141. [Google Scholar] [CrossRef]
- Huang, L.; Snyder, A.R.; Morgan, W.F. Radiation-induced genomic instability and its implications for radiation carcinogenesis. Oncogene 2003, 22, 5848–5854. [Google Scholar] [CrossRef] [Green Version]
- Sachs, R.K.; Chen, A.M.; Brenner, D.J. Review: Proximity effects in the production of chromosome aberrations by ionizing radiation. Int. J. Radiat. Biol. 1997, 71, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lukášová, E.; Kozubek, S.; Kozubek, M.; Kjeronská, J.; Rýznar, L.; Horáková, J.; Krahulcová, E.; Horneck, G. Localisation and distance between ABL and BCR genes in interphase nuclei of bone marrow cells of control donors and patients with chronic myeloid leukaemia. Hum. Genet. 1997, 100, 525–535. [Google Scholar] [CrossRef]
- Kozubek, S.; Lukášová, E.; Rýznar, L.; Kozubek, M.; Lišková, A.; Govorun, R.D.; Krasavin, E.A.; Horneck, G. Distribution of ABL and BCR Genes in Cell Nuclei of Normal and Irradiated Lymphocytes. Blood 1997, 89, 4537–4545. [Google Scholar] [CrossRef] [Green Version]
- Boerma, E.G.; Siebert, R.; Kluin, P.M.; Baudis, M. Translocations involving 8q24 in Burkitt lymphoma and other malignant lymphomas: A historical review of cytogenetics in the light of todays knowledge. Leukemia 2009, 23, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafourková, A.; Lukásová, E.; Kozubek, S.; Kozubek, M.; Govorun, R.D.; Koutná, I.; Bártová, E.; Skalníková, M.; Jirsová, P.; Paseková, R.; et al. Exchange aberrations among 11 chromosomes of human lymphocytes induced by γ-rays. Int. J. Radiat. Biol. 2001, 77, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Boei, J.J.W.A.; Fomina, J.; Darroudi, F.; Nagelkerke, N.J.D.; Mullenders, L.H.F. Interphase Chromosome Positioning Affects the Spectrum of Radiation-Induced Chromosomal Aberrations. Radiat. Res. 2006, 166, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Stevens, D.L.; Goodhead, D.T. M-FISH analysis shows that complex chromosome aberrations induced by -particle tracks are cumulative products of localized rearrangements. Proc. Natl. Acad. Sci. USA 2002, 99, 12167–12172. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.M.; Papworth, D.G.; Stevens, D.L.; Sumption, N.D.; Goodhead, D.T. Increased complexity of radiation-induced chromosome aberrations consistent with a mechanism of sequential formation. Cytogenet. Genome Res. 2006, 112, 35–44. [Google Scholar] [CrossRef]
- Küppers, R.; Rajewsky, K. The origin of Hodgkin and Reed/Sternberg cells in Hodgkin’s disease. Annu. Rev. Immunol. 1998, 16, 471–493. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [Green Version]
- Guffei, A.; Sarkar, R.; Klewes, L.; Righolt, C.; Knecht, H.; Mai, S. Dynamic chromosomal rearrangements in Hodgkin’s lymphoma are due to ongoing three-dimensional nuclear remodeling and breakage-bridge-fusion cycles. Haematologica 2010, 95, 2038–2046. [Google Scholar] [CrossRef]
- Knecht, H.; Sawan, B.; Lichtensztejn, D.; Lemieux, B.; Wellinger, R.J.; Mai, S. The 3D nuclear organization of telomeres marks the transition from Hodgkin to Reed–Sternberg cells. Leukemia 2009, 23, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Righolt, C.H.; Knecht, H.; Mai, S. DNA Superresolution Structure of Reed-Sternberg Cells Differs Between Long-Lasting Remission Versus Relapsing Hodgkin’s Lymphoma Patients. J. Cell. Biochem. 2016, 117, 1633–1637. [Google Scholar] [CrossRef]
- Germini, D.; Tsfasman, T.; Klibi, M.; El-Amine, R.; Pichugin, A.; Iarovaia, O.V.; Bilhou-Nabera, C.; Subra, F.; Bou Saada, Y.; Sukhanova, A.; et al. HIV Tat induces a prolonged MYC relocalization next to IGH in circulating B-cells. Leukemia 2017, 31, 2515–2522. [Google Scholar] [CrossRef] [PubMed]
- Hapgood, G.; Savage, K.J. The biology and management of systemic anaplastic large cell lymphoma. Blood 2015, 126, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathas, S.; Kreher, S.; Meaburn, K.J.; Johrens, K.; Lamprecht, B.; Assaf, C.; Sterry, W.; Kadin, M.E.; Daibata, M.; Joos, S.; et al. Gene deregulation and spatial genome reorganization near breakpoints prior to formation of translocations in anaplastic large cell lymphoma. Proc. Natl. Acad. Sci. USA 2009, 106, 5831–5836. [Google Scholar] [CrossRef] [Green Version]
- Steininger, A.; Ebert, G.; Becker, B.V.; Assaf, C.; Möbs, M.; Schmidt, C.A.; Grabarczyk, P.; Jensen, L.R.; Przybylski, G.K.; Port, M.; et al. Genome-Wide Analysis of Interchromosomal Interaction Probabilities Reveals Chained Translocations and Overrepresentation of Translocation Breakpoints in Genes in a Cutaneous T-Cell Lymphoma Cell Line. Front. Oncol. 2018, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Tavares-Cadete, F.; Norouzi, D.; Dekker, B.; Liu, Y.; Dekker, J. Multi-contact 3C reveals that the human genome during interphase is largely not entangled. Nat. Struct. Mol. Biol. 2020, 27, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Su, J.-H.; Zheng, P.; Kinrot, S.S.; Bintu, B.; Zhuang, X. Genome-Scale Imaging of the 3D Organization and Transcriptional Activity of Chromatin. Cell 2020, 182, 1641–1659. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Allen, W.E.; Deisseroth, K. Three-dimensional in situ sequencing of single cells in intact tissue. FASEB J. 2019, 33, 221.3. [Google Scholar] [CrossRef]
- Wang, X.; Allen, W.E.; Wright, M.A.; Sylwestrak, E.L.; Samusik, N.; Vesuna, S.; Evans, K.; Liu, C.; Ramakrishnan, C.; Liu, J.; et al. Three-dimensional intact-tissue sequencing of single-cell transcriptional states. Science 2018, 361, eaat5691. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Tang, F. Recent advances in single-cell sequencing technologies. Precis. Clin. Med. 2022, 5, pbac002. [Google Scholar] [CrossRef]
- Biancalani, T.; Scalia, G.; Buffoni, L.; Avasthi, R.; Lu, Z.; Sanger, A.; Tokcan, N.; Vanderburg, C.R.; Segerstolpe, Å.; Zhang, M.; et al. Deep learning and alignment of spatially resolved single-cell transcriptomes with Tangram. Nat. Methods 2021, 18, 1352–1362. [Google Scholar] [CrossRef]
- Fabian-Morales, E.; Vallejo-Escamilla, D.; Gudiño, A.; Rodríguez, A.; González-Barrios, R.; Rodríguez Torres, Y.L.; Castro Hernández, C.; Torre-Luján, A.H.; Oliva-Rico, D.A.; Ornelas Guzmán, E.C.; et al. Large-scale topological disruption of chromosome territories 9 and 22 is associated with nonresponse to treatment in CML. Int. J. Cancer 2022, 150, 1455–1470. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, M.F.d.; Lisboa, M.d.O.; Terceiro, L.E.L.; Rangel-Pozzo, A.; Mai, S. Chromosome Territories in Hematological Malignancies. Cells 2022, 11, 1368. https://doi.org/10.3390/cells11081368
Lima MFd, Lisboa MdO, Terceiro LEL, Rangel-Pozzo A, Mai S. Chromosome Territories in Hematological Malignancies. Cells. 2022; 11(8):1368. https://doi.org/10.3390/cells11081368
Chicago/Turabian StyleLima, Matheus Fabiao de, Mateus de Oliveira Lisboa, Lucas E. L. Terceiro, Aline Rangel-Pozzo, and Sabine Mai. 2022. "Chromosome Territories in Hematological Malignancies" Cells 11, no. 8: 1368. https://doi.org/10.3390/cells11081368