
Total and Mitochondrial Transcriptomic and Proteomic Insights into Regulation of Bioenergetic Processes for Shoot Fast-Growth Initiation in Moso Bamboo

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Moso Bamboo Shoots
2.2. Measurement of Physiological Indexes
2.3. Mitochondrial Isolation and Detection of Mitochondrial Activity, Purity and Count
2.4. RNA-Seq Analysis
2.5. RT-qPCR Analysis
2.6. Label-Free Quantitative Proteomics and PRM Verification
2.7. Gene Cloning, Plasmid Construction, Transformation and Screening for Homozygous Lines
2.8. Cold-Stress Tolerance Analysis
2.9. Statistical Analysis
3. Results
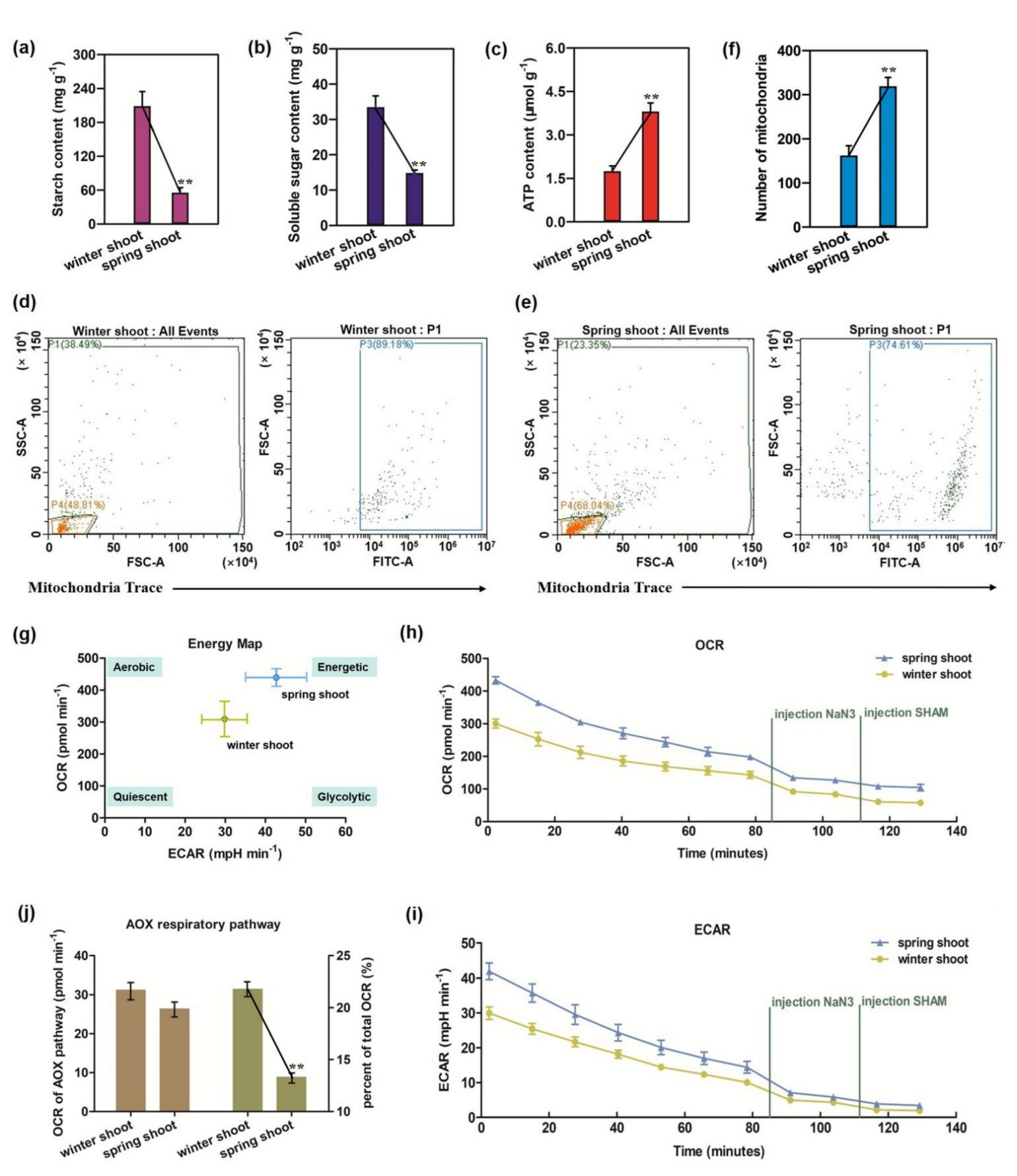
3.1. Enhanced Repiration and Energy Production and Mitochondria Biogenesis in Spring Shoots
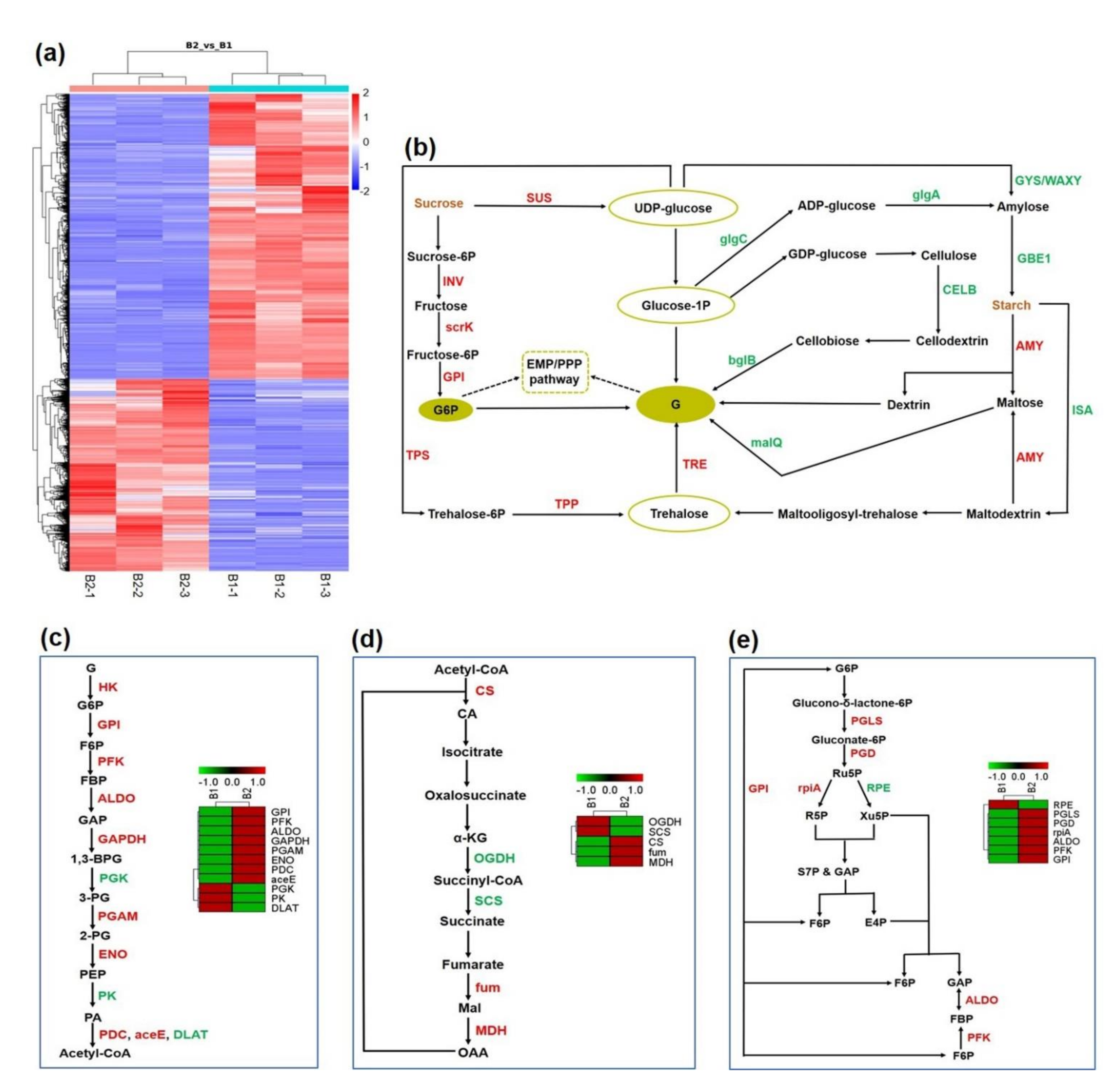
3.2. The Systemic Energy Generation Pathway Was Activated at the Transcriptional Level during Shoot Growth
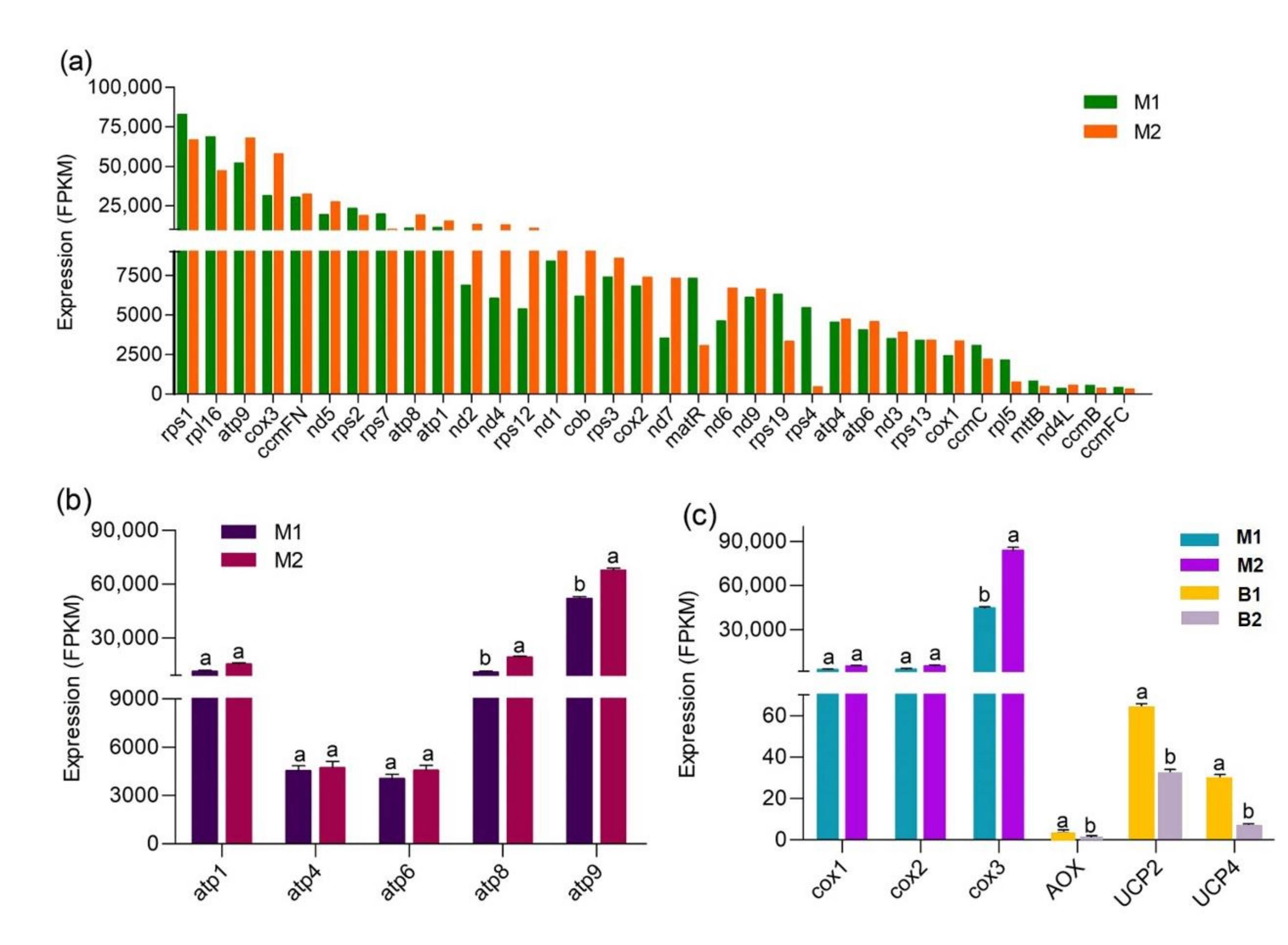
3.3. The Unique Gene-Expression Profile and Energy-Metabolism System in Mitochondria
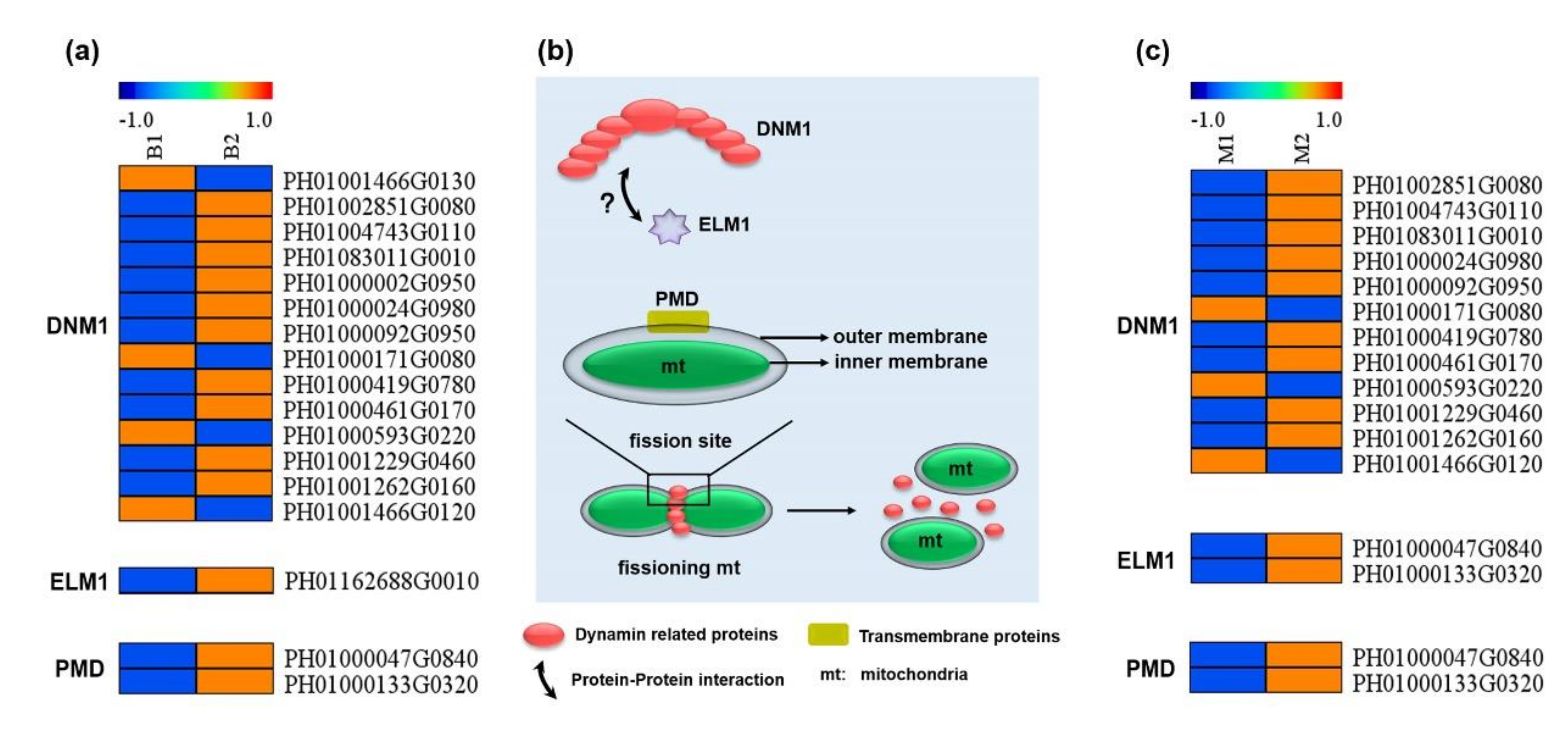
3.4. Enhanced Expression of Genes Related to Mitochondrial Fission in Spring Shoots
3.5. The Systemic Energy Generation Pathway Was Induced at the Protein Level during Shoot Growth
3.6. More Proteins Related to Energy Metabolism and Mitochondrial Fission Were Identified in the Mitochondrial Proteome
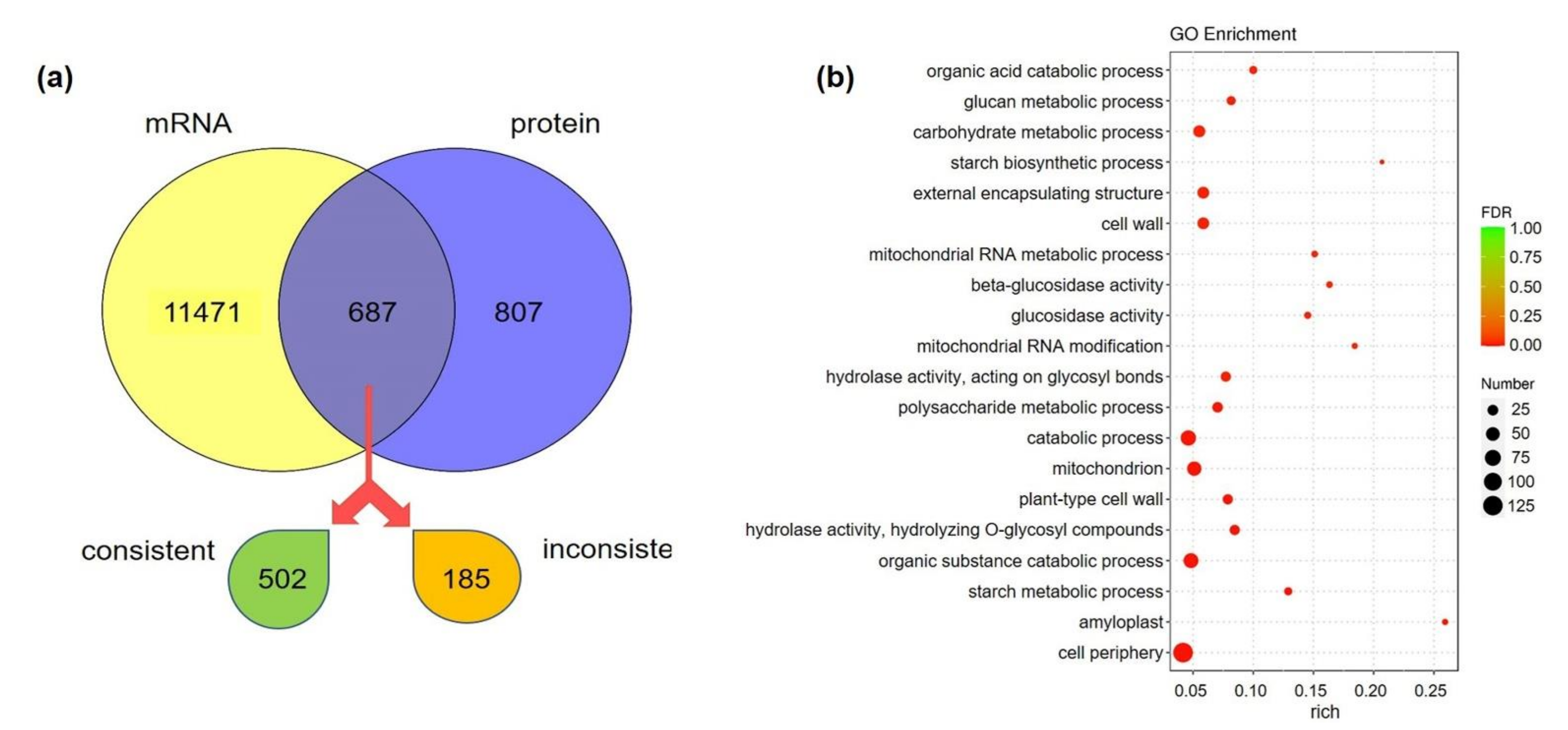
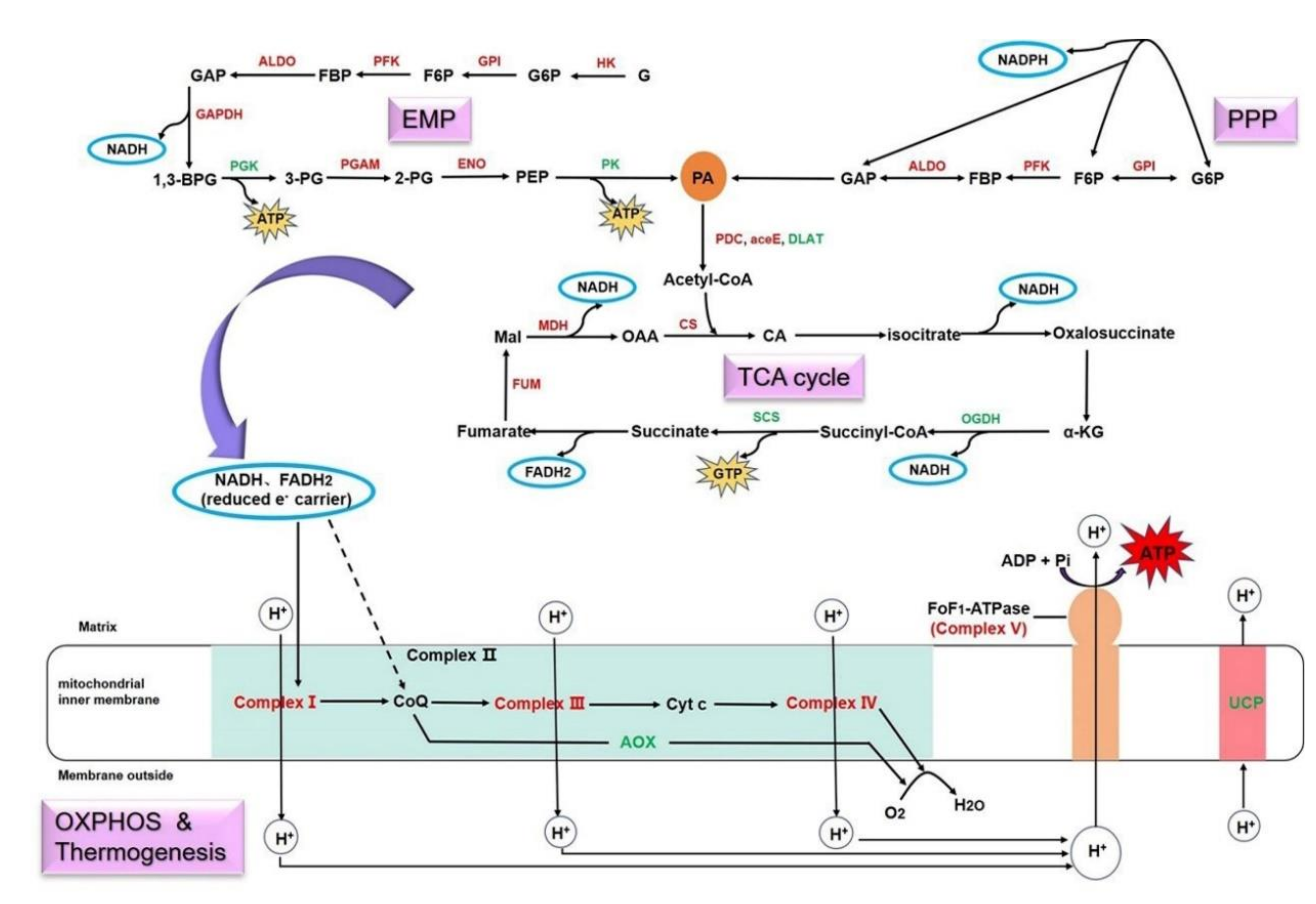
3.7. Conjoint Analysis Revealed the Energy Metabolic Mechanism of Fast-Growth Initiation in Bamboo Shoots
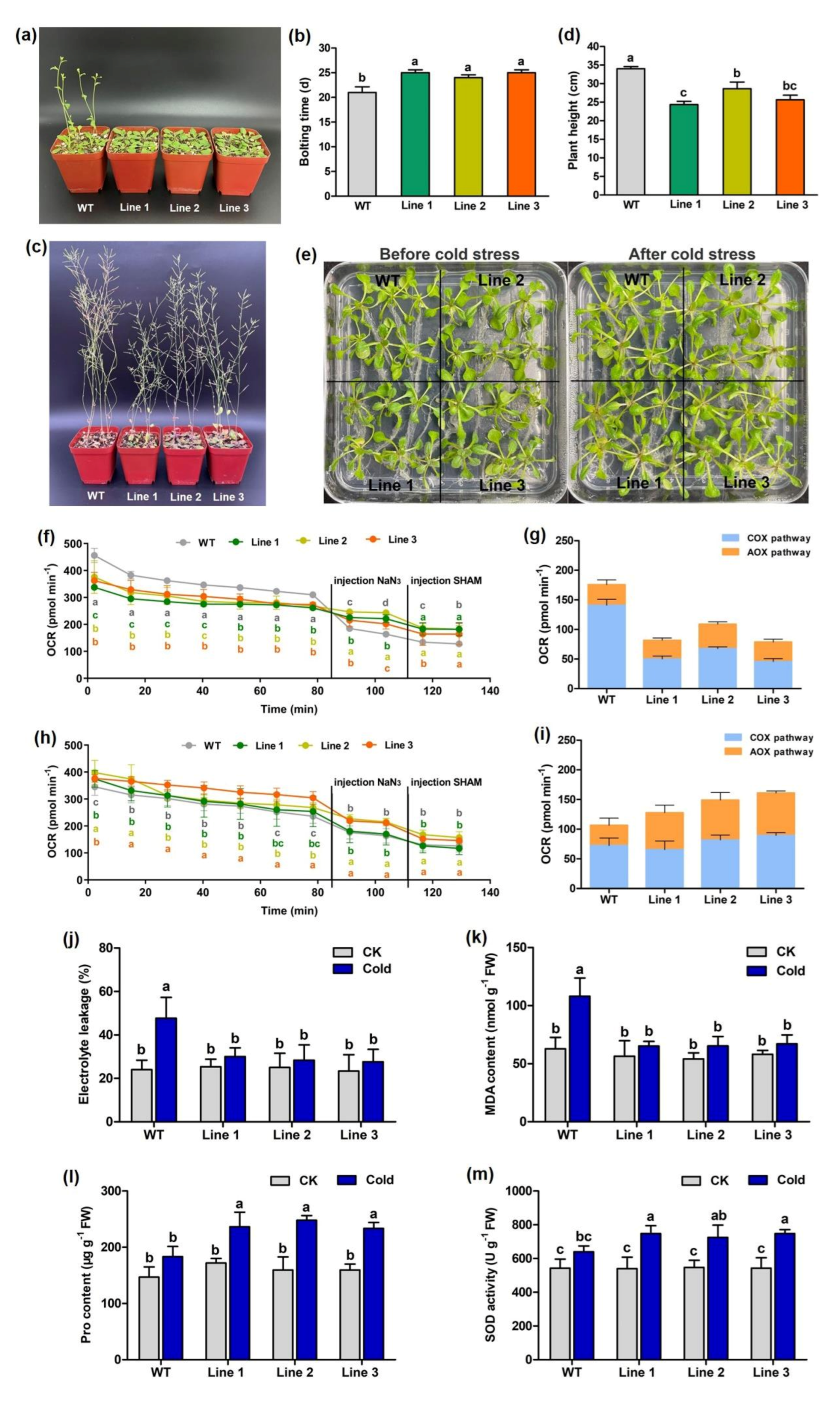
3.8. Heterologous Expression of PeAOX1b in Arabidopsis Affected Seedling Growth and Enhanced Cold-Stress Tolerance
4. Discussion
4.1. Respiration and Energy Production Increased during the Transition from Winter to Spring Shoots
4.2. Different Ways of Energy Production Activated in Spring Shoots
4.3. Fission Activity and Number of Mitochondria Increased during the Transition from Winter to Spring Shoots
4.4. Mitochondrial Energy-Dissipating Systems Involved in the Adaptation of Bamboo Shoots to the Winter Environment
5. Conclusions
- Efficient energy-regulation mechanisms are essential for initiation of fast growth in Moso bamboo shoots; this entails a continuous process of carbohydrate oxidation consisting of starch and sucrose catabolism, EMP, the TCA cycle, the PPP and OXPHOS;
- Mitochondrial fission increased starkly in accordance with the higher energy demand and cell division required for fast growth;
- The revelation regarding the AOX pathway and UCP pathway and their enhanced roles in winter shoots reflects different adaptive heat requirements during shoot growth. These hypotheses suggest new avenues for further research into the mechanisms of bamboo fast growth and potentially for applications in edible-winter-shoot storage.
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, Z.H.; Lu, Y.; Li, L.B.; Zhao, Q.; Feng, Q.; Gao, Z.M.; Lu, H.Y.; Hu, T.; Yao, N.; Liu, K.Y.; et al. The draft genome of the fast-growing non-timber forest species Moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, M.; Yrjala, K.; Vinod, K.K.; Sharma, A.; Cho, J.N.; Satheesh, V.; Zhou, M.B. Genetics and genomics of Moso bamboo (Phyllostachys edulis): Current status, future challenges, and biotechnological opportunities toward a sustainable bamboo industry. Food Energy Secur. 2020, 9, e229. [Google Scholar] [CrossRef]
- Wu, R.H.; Shi, Y.R.; Zhang, Q.; Zheng, W.Q.; Chen, S.L.; Du, L.; Lu, C.F. Genome-wide identification and characterization of the UBP gene family in Moso bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 20, 4309. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.Q.; Zhang, Y.; Zhang, Q.; Wu, R.H.; Wang, X.W.; Feng, S.N.; Chen, S.L.; Lu, C.F.; Du, L. Genome-wide identification and characterization of hexokinase genes in Moso bamboo (Phyllostachys edulis). Front. Plant Sci. 2020, 11, 600. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kang, L.; Wu, R.H.; Chen, Y.Z.; Lu, C.F. Genome-wide identification and characterization of UDP-glucose dehydrogenase family genes in Moso bamboo and functional analysis of PeUGDH4 in hemicellulose synthesis. Sci. Rep. 2020, 10, 10124. [Google Scholar] [CrossRef] [PubMed]
- Gamuyao, R.; Nagai, K.; Ayano, M.; Mori, Y.; Minami, A.; Kojima, M.; Suzuki, T.; Sakakibara, H.; Higashiyama, T.; Ashikari, M.; et al. Hormone distribution and transcriptome profiles in bamboo shoots provide insights on bamboo stem emergence and growth. Plant Cell Physiol. 2017, 58, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Yan, X.Y.; Li, S.B.; Jing, Y.; Gu, L.F.; Zou, S.Q.; Zhang, J.; Liu, B.B. Genome-wide identification, evolution and expression analysis of the aspartic protease gene family during rapid growth of Moso bamboo (Phyllostachys edulis) shoots. BMC Genom. 2017, 22, 45. [Google Scholar] [CrossRef] [PubMed]
- Tao, G.Y.; Ramakrishnan, M.; Vinod, K.K.; Yrjala, K.; Satheesh, V.; Cho, J.; Fu, Y.; Zhou, M.B. Multi-omics analysis of cellular pathways involved in different rapid growth stages of Moso bamboo. Tree Physiol. 2020, 40, 1487–1508. [Google Scholar] [CrossRef]
- Li, L.; Cheng, Z.C.; Ma, Y.J.; Bai, Q.S.; Li, X.Y.; Cao, Z.H.; Wu, Z.N.; Gao, J. The association of hormone signalling genes, transcription and changes in shoot anatomy during Moso bamboo growth. Plant Biotechnol. J. 2018, 16, 72–85. [Google Scholar] [CrossRef]
- Wei, Q.; Jiao, C.; Ding, Y.L.; Gao, S.; Guo, L.; Chen, M.; Hu, P.; Xia, S.J.; Ren, G.D.; Fei, Z.J. Cellular and molecular characterizations of a slow-growth variant provide insights into the fast growth of bamboo. Tree Physiol. 2018, 38, 641–654. [Google Scholar] [CrossRef]
- Wei, Q.; Guo, L.; Jiao, C.; Fei, Z.J.; Chen, M.; Cao, J.J.; Ding, Y.L.; Yuan, Q.S. Characterization of the developmental dynamics of the elongation of a bamboo internode during the fast growth stage. Tree Physiol. 2019, 39, 1201–1214. [Google Scholar] [CrossRef]
- Zhao, J.W.; Gao, P.J.; Li, C.L.; Lin, X.C.; Guo, X.Q.; Liu, S.K. PhePEBP family genes regulated by plant hormones and drought are associated with the activation of lateral buds and seedling growth in Phyllostachys edulis. Tree Physiol. 2019, 39, 1387–1404. [Google Scholar] [CrossRef] [PubMed]
- Bhandawat, A.; Singh, G.; Seth, R.; Singh, P.; Sharma, R.K. Genome-wide transcriptional profiling to elucidate key candidates involved in bud burst and rattling growth in a subtropical bamboo (Dendrocalamus hamiltonii). Front. Plant Sci. 2017, 7, 2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.H.; Peng, H.Z.; Lin, E.P.; Jin, Q.Y.; Hua, X.Q.; Yao, S.; Bian, H.W.; Han, N.; Pan, J.W.; Wang, J.H.; et al. Identification of genes related to the development of bamboo rhizome bud. J. Exp. Bot. 2010, 61, 551–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.; Wang, W.J.; Yao, N.; Cai, C.Y.; Liu, Y.Y.; Lin, C.T.; Zuo, Z.C.; Zhu, Q. The transcriptional dynamics during de novo shoot organogenesis of Ma bamboo (Dendrocalamus latiflorus Munro): Implication of the contributions of the abiotic stress response in this process. Plant J. 2021, 107, 1513–1532. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Wang, H.Y.; Cai, D.W.; Gao, Y.B.; Zhang, H.X.; Wang, Y.S.; Lin, C.T.; Ma, L.Y.; Gu, L.F. Comprehensive profiling of rhizome-associated alternative splicing and alternative polyadenylation in Moso bamboo (Phyllostachys edulis). Plant J. 2017, 91, 684–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, S.B.; Costa-Pereira, R.; Fleming, T.; Goulding, M.; Anderson, J.T. Neotropical fish-fruit interactions: Eco-evolutionary dynamics and conservation. Biol. Rev. 2015, 90, 1263–1278. [Google Scholar] [CrossRef]
- Shou, Y.T.; Zhu, Y.H.; Ding, Y.L. Transcriptome analysis of lateral buds from Phyllostachys edulis rhizome during germination and early shoot stages. BMC Plant Biol. 2020, 20, 229. [Google Scholar] [CrossRef]
- Wei, Q.; Jiao, C.; Guo, L.; Ding, Y.L.; Cao, J.J.; Feng, J.Y.; Dong, X.B.; Mao, L.Y.; Sun, H.H.; Yu, F.; et al. Exploring key cellular processes and candidate genes regulating the primary thickening growth of Moso underground shoots. New Phytol. 2017, 214, 81–96. [Google Scholar] [CrossRef]
- Pan, F.; Wu, M.; Hu, W.F.; Liu, R.; Yan, H.W.; Xiang, Y. Genome-wide identification and expression analyses of the bZIP transcription factor genes in Moso bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 20, 2203. [Google Scholar] [CrossRef] [Green Version]
- He, Y.Q.; Cheng, J.P.; He, Y.; Yang, B.; Cheng, Y.H.; Yang, C.; Zhang, H.S.; Wang, Z.F. Influence of isopropylmalate synthase OsIPMS1 on seed vigour associated with amino acid and energy metabolism in rice. Plant Biotechnol. J. 2019, 17, 322–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domergue, J.B.; Abadie, C.; Limami, A.; Way, D.; Tcherkez, G. Seed quality and carbon primary metabolism. Plant Cell Environ. 2019, 42, 2776–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Li, J.Y. Molecular basis of plant architecture. Annu. Rev. Plant Biol. 2008, 59, 253–279. [Google Scholar] [CrossRef]
- Wang, Y.J.; Sun, X.P.; Ding, Y.L.; Fei, Z.J.; Jiao, C.; Fan, M.Y.; Yao, B.F.; Xin, P.Y.; Chu, J.F.; Wei, Q. Cellular and molecular characterization of a thick-walled variant reveal a pivotal role of shoot apical meristem in transverse development of bamboo culm. J. Exp. Bot. 2019, 70, 3911–3926. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Z.; Peng, C.H.; Zhou, G.M.; Gu, H.H.; Li, Q.; Zhang, C. Dynamic allocation and transfer of non-structural carbohydrates, a possible mechanism for the explosive growth of Moso bamboo (Phyllostachys heterocycla). Sci. Rep. 2016, 6, 25908. [Google Scholar] [CrossRef]
- Meyer, E.H.; Welchen, E.; Carrie, C. Assembly of the complexes of the oxidative phosphorylation system in land plant mitochondria. Annu. Rev. Plant Biol. 2019, 70, 23–50. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Neph, S.; Dinger, M.E.; Crawford, J.; Smith, M.A.; Shearwood, A.M.J.; Haugen, E.; Bracken, C.P.; Rackham, O.; Stamatoyannopoulos, J.A.; et al. The human mitochondrial transcriptome. Cell 2011, 146, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.J.; Yang, Y.M.; Wu, L.; Zou, M.T.; Huang, Y. The S-pombe mitochondrial transcriptome. RNA 2018, 24, 1241–1254. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.C.; Su, Y.Y.; Wu, C.H.; Liu, Y.C.; Huang, C.H.; Chang, C.C. Analysis of mitochondrial genomics and transcriptomics reveal abundant RNA edits and differential editing status in moth orchid, Phalaenopsis aphrodite subsp. formosana. Sci. Hortic. 2020, 267, 109304. [Google Scholar] [CrossRef]
- Nie, H.S.; Cheng, C.; Hua, J.P. Mitochondrial proteomic analysis reveals that proteins relate to oxidoreductase activity play a central role in pollen fertility in cotton. J. Proteom. 2020, 225, 103861. [Google Scholar] [CrossRef]
- Dahal, D.; Mooney, B.P.; Newton, K.J. Specific changes in total and mitochondrial proteomes are associated with higher levels of heterosis in maize hybrids. Plant J. 2012, 72, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.D.; Storchova, H. The application of RNA-seq to the comprehensive analysis of plant mitochondrial transcriptomes. Mol. Genet. Genom. 2015, 290, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sew, Y.S.; Stroher, E.; Holzmann, C.; Huang, S.B.; Taylor, N.L.; Jordana, X.; Millar, A.H. Multiplex micro-respiratory measurements of Arabidopsis tissues. New Phytol. 2013, 200, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Leger, J.L.; Jougleux, J.L.; Savadogo, F.; Pichaud, N.; Boudreau, L.H. Rapid isolation and purification of functional platelet mitochondria using a discontinuous Percoll gradient. Platelets 2020, 31, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, L.H.; Duchez, A.C.; Cloutier, N.; Soulet, D.; Martin, N.; Bollinger, J.; Paré, A.; Rousseau, M.; Naika, G.S.; Lévesque, T.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood 2014, 124, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.Z.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Wang, X.J.; Liang, H.Y.; Guo, D.L.; Guo, L.L.; Duan, X.G.; Jia, Q.S.; Hou, X.G. Integrated analysis of transcriptomic and proteomic data from tree peony (P. ostii) seeds reveals key developmental stages and candidate genes related to oil biosynthesis and fatty acid metabolism. Hortic. Res. 2019, 6, 111. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Yu, X.L.; Wang, Y.S.; Kohnen, M.V.; Piao, M.X.; Tu, M.; Gao, Y.B.; Lin, C.T.; Zuo, Z.C.; Gu, L.F. Large scale profiling of protein isoforms using Label-free quantitative proteomics revealed the regulation of nonsense-mediated decay in Moso bamboo (Phyllostachys edulis). Cells 2019, 8, 744. [Google Scholar] [CrossRef] [Green Version]
- Su, L.Y.; Dai, Z.W.; Li, S.H.; Xin, H.P. A novel system for evaluating drought-cold tolerance of grapevines using chlorophyll fluorescence. BMC Plant Biol. 2015, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schertl, P.; Braun, H.P. Respiratory electron transfer pathways in plant mitochondria. Front. Plant Sci. 2014, 5, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, N.L. Editorial for special issue “plant mitochondria”. Int. J. Mol. Sci. 2018, 19, 3849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, A.L.; Umbach, A.L.; Siedow, J.N. Structure-function relationships of the alternative oxidase of plant mitochondria: A model of the active site. J. Bioenerg. Biomembr. 1995, 27, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Laloi, M.; Klein, M.; Riesmeier, J.W.; Muller-Rober, B.; Fleury, C.; Bouillaud, F.; Ricquier, D. A plant cold-induced uncoupling protein. Nature 1997, 389, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S. Fission and fusion of plant mitochondria, and genome maintenance. Plant Physiol. 2018, 176, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, F.; Sheen, J. Sugar sensing and signalling networks in plants. Biochem. Soc. T. 2005, 33, 269–271. [Google Scholar] [CrossRef]
- Baena-Gonzalez, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938–942. [Google Scholar] [CrossRef]
- Wang, S.; Ding, Y.; Lin, S.; Ji, X.; Zhan, H. Seasonal changes of endogenous soluble sugar and starch in different developmental stages of Fargesia yunnanensis. J. Wood Sci. 2016, 62, 1–11. [Google Scholar] [CrossRef]
- Wang, S.G.; Pei, J.L.; Li, J.; Tang, G.J.; Zhao, J.W.; Peng, X.P.; Nie, S.X.; Ding, Y.L.; Wang, C.M. Sucrose and starch metabolism during Fargesia yunnanensis shoot growth. Physiol. Plantarum 2020, 168, 188–204. [Google Scholar] [CrossRef]
- Shang, C.Q.; Yang, H.L.; Ma, S.; Shen, Q.D.; Liu, L.; Hou, C.X.; Cao, X.; Cheng, J.L. Physiological and transcriptomic changes during the early phases of adventitious root formation in mulberry stem hardwood cuttings. Int. J. Mol. Sci. 2019, 20, 3707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Liu, Y.; Loka, D.A.; Zahoor, R.; Wang, S.S.; Zhou, Z.G. Drought limits pollen tube growth rate by altering carbohydrate metabolism in cotton (Gossypium hirsutum) pistils. Plant Sci. 2019, 286, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; He, C.Y.; Zhang, J.G.; Duan, A.G.; Zeng, Y.F. Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo. J. Proteome Res. 2012, 11, 2492–2507. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logan, D.C. Arabidopsis seed mitochondria are bioenergetically active immediately upon imbibition and specialize via biogenesis in preparation for autotrophic growth. Plant Cell 2017, 29, 109–128. [Google Scholar] [CrossRef]
- Law, S.R.; Narsai, R.; Whelan, J. Mitochondrial biogenesis in plants during seed germination. Mitochondrion 2014, 19, 214–221. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Kuroiwa, T. The discovery of the division apparatus of plastids and mitochondria. J. Electron Microsc. 2000, 49, 123–134. [Google Scholar] [CrossRef]
- Mishra, P.; Chan, D.C. Metabolic regulation of mitochondrial dynamics. J. Cell Biol. 2016, 212, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.Y.; Wu, C.C.; Lin, C.C.; Jane, W.N.; Suen, D.F. 3D imaging of tapetal mitochondria suggests the importance of mitochondrial fission in pollen growth. Plant Physiol. 2019, 180, 813–826. [Google Scholar] [CrossRef] [Green Version]
- Bleazard, W.; McCaffery, J.M.; King, E.J.; Bale, S.; Mozdy, A.; Tieu, Q.; Nunnari, J.; Shaw, J.M. The dynamin-related GTPase Dnm1 regulates mitochondrial fission in yeast. Nat. Cell Biol. 1999, 1, 298–304. [Google Scholar] [CrossRef]
- Fujimoto, M.; Arimura, S.; Mano, S.; Kondo, M.; Saito, C.; Ueda, T.; Nakazono, M.; Nakano, A.; Nishimura, M.; Tsutsumi, N. Arabidopsis dynamin-related proteins DRP3A and DRP3B are functionally redundant in mitochondrial fission, but have distinct roles in peroxisomal fission. Plant J. 2009, 58, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Mano, S.; Nakamori, C.; Kondo, M.; Hayashi, M.; Nishimura, M. An Arabidopsis dynamin-related protein, DRP3A, controls both peroxisomal and mitochondrial division. Plant J. 2004, 38, 487–498. [Google Scholar] [CrossRef]
- Arimura, S.; Fujimoto, M.; Doniwa, Y.; Kadoya, N.; Nakazono, M.; Sakamoto, W.; Tsutsumi, N. Arabidopsis ELONGATED MITOCHONDRIA1 is required for localization of DYNAMIN-RELATED PROTEIN3A to mitochondrial fission sites. Plant Cell 2008, 20, 1555–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, K.; Hu, J.P. The Arabidopsis tail-anchored protein PEROXISOMAL AND MITOCHONDRIAL DIVISION FACTOR1 is involved in the morphogenesis and proliferation of peroxisomes and mitochondria. Plant Cell 2011, 23, 4446–4461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onda, Y.; Kato, Y.; Abe, Y.; Ito, T.; Morohashi, M.; Ito, Y.; Ichikawa, M.; Matsukawa, K.; Kakizaki, Y.; Koiwa, H.; et al. Functional coexpression of the mitochondrial alternative oxidase and uncoupling protein underlies thermoregulation in the thermogenic florets of skunk cabbage. Plant Physiol. 2008, 146, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabelnych, O.I.; Kolesnichenko, A.V.; Pobezhimova, T.P.; Tourchaninova, V.V.; Korzun, A.M.; Koroleva, N.A.; Zykova, V.V.; Voinikov, V.K. The role of different plant seedling shoots mitochondrial uncoupling systems in thermogenesis during low-temperature stress. J. Therm. Biol. 2003, 28, 571–580. [Google Scholar] [CrossRef]
- Grant, N.M.; Miller, R.E.; Watling, J.R.; Robinson, S.A. Synchronicity of thermogenic activity, alternative pathway respiratory flux, AOX protein content, and carbohydrates in receptacle tissues of sacred lotus during floral development. J. Exp. Bot. 2008, 59, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Gibernau, M.; Barabe, D.; Moisson, M.; Trombe, A. Physical constraints on temperature difference in some thermogenic aroid inflorescences. Ann. Bot. 2005, 96, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Seymour, R.S. Scaling of heat production by thermogenic flowers: Limits to floral size and maximum rate of respiration. Plant Cell Environ. 2010, 33, 1474–1485. [Google Scholar] [CrossRef]
- Ito-Inaba, Y.; Sato, M.; Sato, M.P.; Kurayama, Y.; Yamamoto, H.; Ohata, M.; Ogura, Y.; Hayashi, T.; Toyooka, K.; Inaba, T. Alternative oxidase capacity of mitochondria in microsporophylls may function in cycad thermogenesis. Plant Physiol. 2019, 180, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Suinyuy, T.N.; Donaldson, J.S.; Johnson, S.D. Patterns of odour emission, thermogenesis and pollinator activity in cones of an African cycad: What mechanisms apply? Ann. Bot. 2013, 112, 891–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, I.; Walter, G.H.; Moore, C.; Roemer, R.; Hull, C. Odor-mediated push-pull pollination in cycads. Science 2007, 318, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, R.M. Heat production and temperature regulation in eastern skunk cabbage. Science 1974, 186, 746–747. [Google Scholar] [CrossRef] [PubMed]
- Chien, L.F.; Wu, Y.C.; Chen, H.P. Mitochondrial energy metabolism in young bamboo rhizomes from Bambusa oldhamii and Phyllostachys edulis during shooting stage. Plant Physiol. Bioch. 2011, 49, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M. PLANT MITOCHONDRIA AND OXIDATIVE STRESS: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Phys. 2001, 52, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Borovskii, G.; Panda, S.K. Alternative oxidase and plant stress tolerance. Plant Signal. Behav. 2016, 11, e1256530. [Google Scholar] [CrossRef]
- Li, C.R.; Liang, D.D.; Li, J.; Duan, Y.B.; Li, H.; Yang, Y.C.; Qin, R.Y.; Li, L.; Wei, P.C.; Yang, J.B. Unravelling mitochondrial retrograde regulation in the abiotic stress induction of rice ALTERNATIVE OXIDASE 1 genes. Plant Cell Environ. 2013, 36, 775–788. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Protein Description | Protein Name | Laber-Free | RNA-Seq | ||
|---|---|---|---|---|---|---|
| logFC | p-Value | logFC | p-Value | |||
| Carbohydrate Metabolism | ||||||
| PH01001178G0320 | 40S ribosomal protein S3a | rpiA | −3.260 | 0.043 | −0.289 | 0.026 |
| PH01003555G0050 | Phosphoglucomutase | pgm | −3.370 | 0.045 | −3.823 | 0.000 |
| PH01002199G0020 | Isocitrate dehydrogenase | IDH3 | 1.203 | 0.049 | 0.901 | 0.000 |
| PH01000533G0580 | Aldehyde dehydrogenase | ALDH | 2.862 | 0.008 | 2.611 | 0.000 |
| PH01000669G0370 | Hexokinase | HK | 1.227 | 0.048 | 0.985 | 0.058 |
| PH01001527G0160 | Pyruvate kinase | PK | −1.930 | 0.017 | −2.068 | 0.000 |
| PH01001262G0280 | Dehydrogenase E1 component domain containing protein | aceE | −3.307 | 0.005 | −9.750 | 0.000 |
| PH01002816G0060 | Citrate synthase | CS | 1.117 | 0.015 | 1.243 | 0.000 |
| PH01000246G0090 | 6-phospho gluconate dehydrogenase decarboxylating | PGD | 1.510 | 0.003 | 1.169 | 0.000 |
| PH01000947G0420 | Aldose1-epimerase | galM | 12.761 | 0.000 | 1.949 | 0.000 |
| Mitochondrial Energy Metabolism | ||||||
| PH01004743G0090 | 2Fe-2S iron-sulfur cluster binding domain containing protein | NDUFS1 | 1.237 | 0.044 | −0.467 | 0.288 |
| PH01000376G0230 | 4Fe-4S binding domain containing protein | NDUFS8 | 1.888 | 0.028 | −0.211 | 0.249 |
| PH01002667G0100 | NADH dehydrogenase 1 alpha subcomplex subunit 12 | NDUFA12 | 11.493 | 0.000 | 0.039 | 0.855 |
| PH01001330G0300 | Solublein organic pyrophosphatase | PPA | 10.091 | 0.005 | 0.789 | 0.000 |
| PH01000704G0310 | ATP synthase | ATPeF1B | 1.206 | 0.020 | 1.084 | 0.000 |
| Protein ID | Protein Description | Protein Name | Laber-free | PRM | ||
|---|---|---|---|---|---|---|
| logFC | p-Value | logFC | p-Value | |||
| Carbohydrate Metabolism | ||||||
| PH01000086G1010 | Succinyl-CoA ligase subunit alpha-2 | LSC1 | 10.721 | 0.000 | 0.571 | 0.050 |
| PH01002199G0020 | Isocitrate dehydrogenase | IDH3 | 2.838 | 0.011 | 0.863 | 0.032 |
| PH01000713G0340 | ATP-citrate synthase subunit1 | ACLY | −1.534 | 0.000 | 1.271 | 0.002 |
| PH01000947G0420 | Aldose1-epimerase | galM | 7.663 | 0.018 | NA | NA |
| PH01001262G0210 | Pyrophosphate-fructose6-phosphate1-phospho transferase subunit beta | pfp | 5.690 | 0.008 | 1.958 | 0.009 |
| PH01003425G0070 | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | −1.366 | 0.000 | 1.556 | 0.000 |
| PH01002248G0250 | Pyruvate phosphate dikinase | ppdK | 3.962 | 0.005 | 1.923 | 0.100 |
| PH01000031G1210 | Fructose-bisphospate aldolase isozyme | ALDO | 3.264 | 0.000 | 1.642 | 0.007 |
| PH01003555G0050 | Phosphorglucomutase | pgm | −9.163 | 0.001 | −1.459 | 0.239 |
| PH01001178G0320 | 40S ribosomal protein S3a | rpiA | −1.871 | 0.034 | −1.702 | 0.000 |
| PH01001444G0350 | Aldehyde dehydrogenase | gapN | 2.787 | 0.003 | −0.075 | 0.855 |
| PH01003304G0200 | Hexokinase | HK | 2.162 | 0.019 | 0.609 | 0.119 |
| PH01000891G0110 | Aldehyde dehydrogenase | ALDH | 10.936 | 0.002 | NA | NA |
| Mitochondrial Energy Metabolism | ||||||
| PH01000322G0030 | Pyruvate decarboxylase isozyme 2 | PDC | 2.396 | 0.020 | 1.601 | 0.138 |
| PH01000597G0060 | H+-transporting ATPase | PMA | 2.193 | 0.036 | 0.825 | 0.023 |
| PH01000965G0370 | F-type H+-transporting ATPase subunit epsilon | ATPF1E | 9.684 | 0.015 | 2.376 | 0.025 |
| PH01001308G0210 | Solublein organic pyrophosphatase | PPA | 4.246 | 0.000 | −0.389 | 0.562 |
| PH01001580G0330 | NADH dehydrogenase 1 alpha subcomplex subunit 9 | NDUFA9 | 2.149 | 0.023 | 1.365 | 0.000 |
| PH01000276G0370 | LYR motif containing protein | NDUFB9 | 2.022 | 0.033 | NA | NA |
| PH01003254G0190 | Ribosomal protein L51 | NDUFA2 | 1.977 | 0.025 | 0.723 | 0.101 |
| PH01002851G0110 | 2Fe-2Siron-sulfur cluster binding domain containing protein | NDUFS1 | 1.644 | 0.045 | 1.034 | 0.006 |
| PH01002719G0170 | Cytochrome b-c1 complex subunit 7 | fbcH | 1.647 | 0.048 | 0.875 | 0.000 |
| PH01000704G0310 | F-type H+-transporting ATPase subunit beta | ATPeF1B | 5.056 | 0.004 | 1.749 | 0.019 |
| Mitochondrial Fission | ||||||
| PH01001466G0130 | Dynamin 1 | Dnm1 | 12.187 | 0.000 | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Geng, X.; Yang, L.; Chen, Y.; Zhao, Z.; Shi, W.; Kang, L.; Wu, R.; Lu, C.; Gao, J. Total and Mitochondrial Transcriptomic and Proteomic Insights into Regulation of Bioenergetic Processes for Shoot Fast-Growth Initiation in Moso Bamboo. Cells 2022, 11, 1240. https://doi.org/10.3390/cells11071240
Wang X, Geng X, Yang L, Chen Y, Zhao Z, Shi W, Kang L, Wu R, Lu C, Gao J. Total and Mitochondrial Transcriptomic and Proteomic Insights into Regulation of Bioenergetic Processes for Shoot Fast-Growth Initiation in Moso Bamboo. Cells. 2022; 11(7):1240. https://doi.org/10.3390/cells11071240
Chicago/Turabian StyleWang, Xiaojing, Xin Geng, Lilin Yang, Yuzhen Chen, Zhiheng Zhao, Weijia Shi, Lan Kang, Ruihua Wu, Cunfu Lu, and Jian Gao. 2022. "Total and Mitochondrial Transcriptomic and Proteomic Insights into Regulation of Bioenergetic Processes for Shoot Fast-Growth Initiation in Moso Bamboo" Cells 11, no. 7: 1240. https://doi.org/10.3390/cells11071240