
Trophectoderm Transcriptome Analysis in LIN28 Knockdown Ovine Conceptuses Suggests Diverse Roles of the LIN28-let-7 Axis in Placental and Fetal Development

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trophectoderm Collection from Day 16 Conceptuses and RNA Sequencing
2.2. Data Pre-Processing
2.3. Identification of the Transcription Factors in the DEGs
2.4. Let-7 miRNAs and Prediction of Their Downstream Target Genes
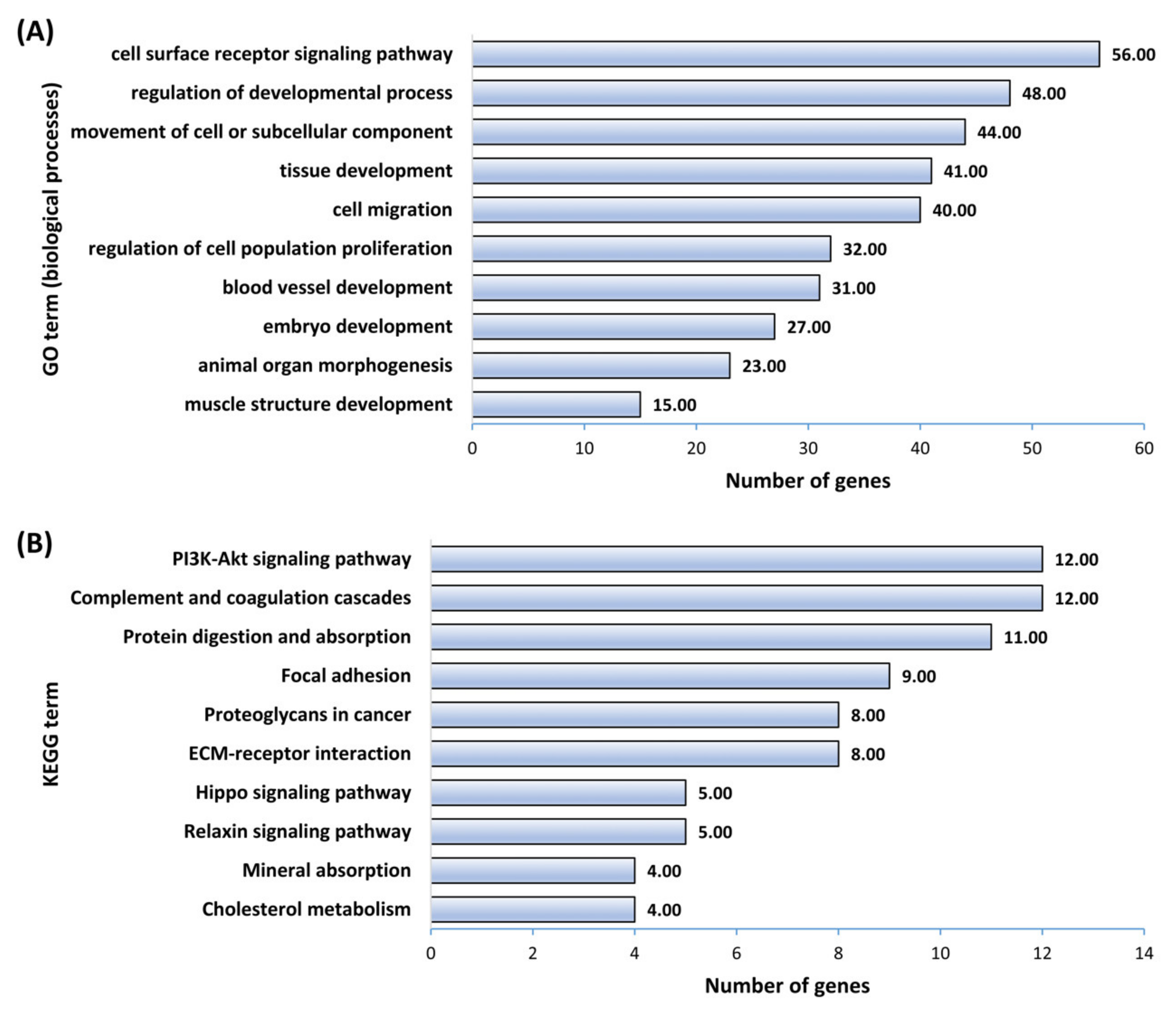
2.5. Gene Ontology and KEGG Pathway Enrichment Analysis
3. Results
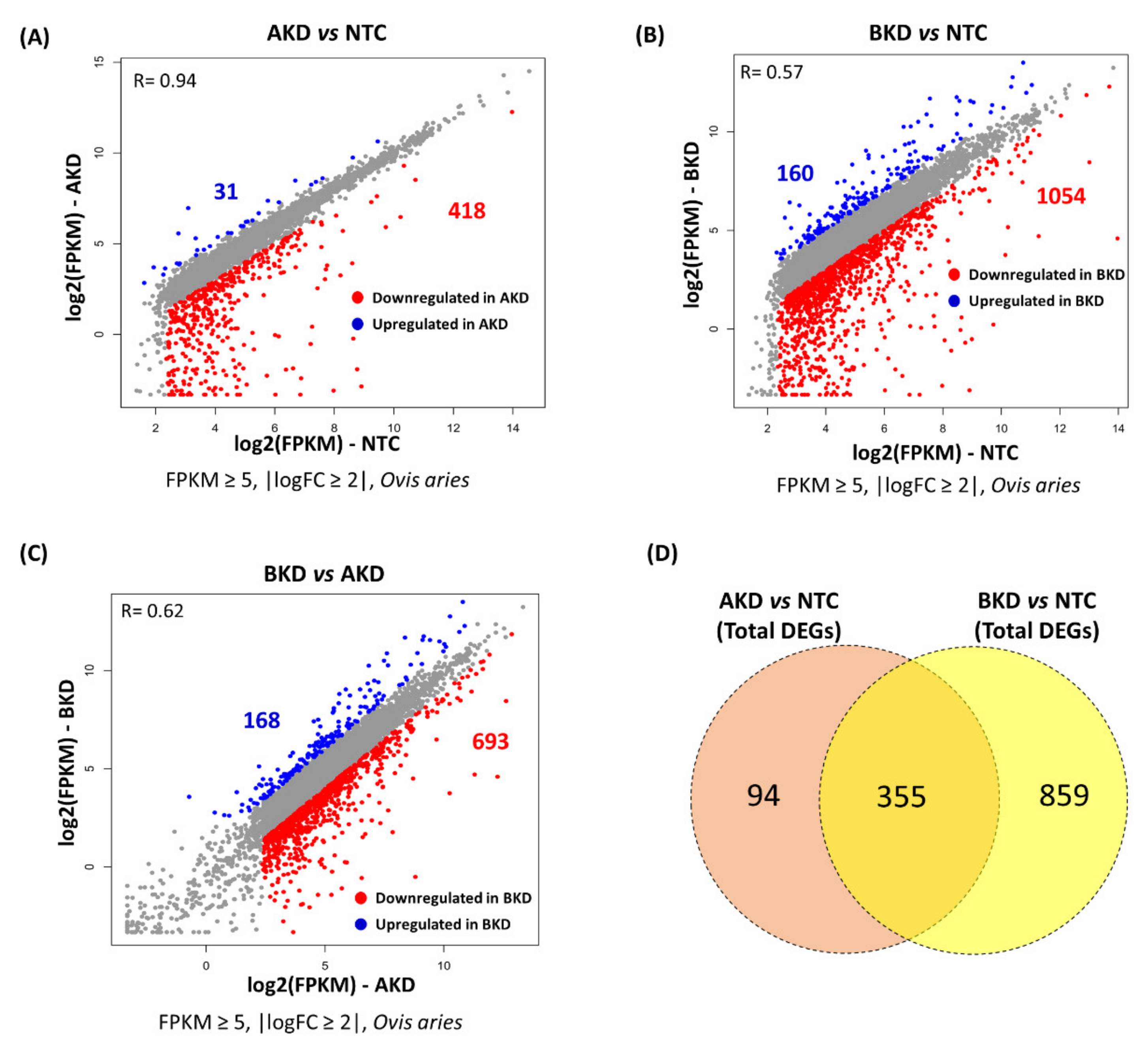
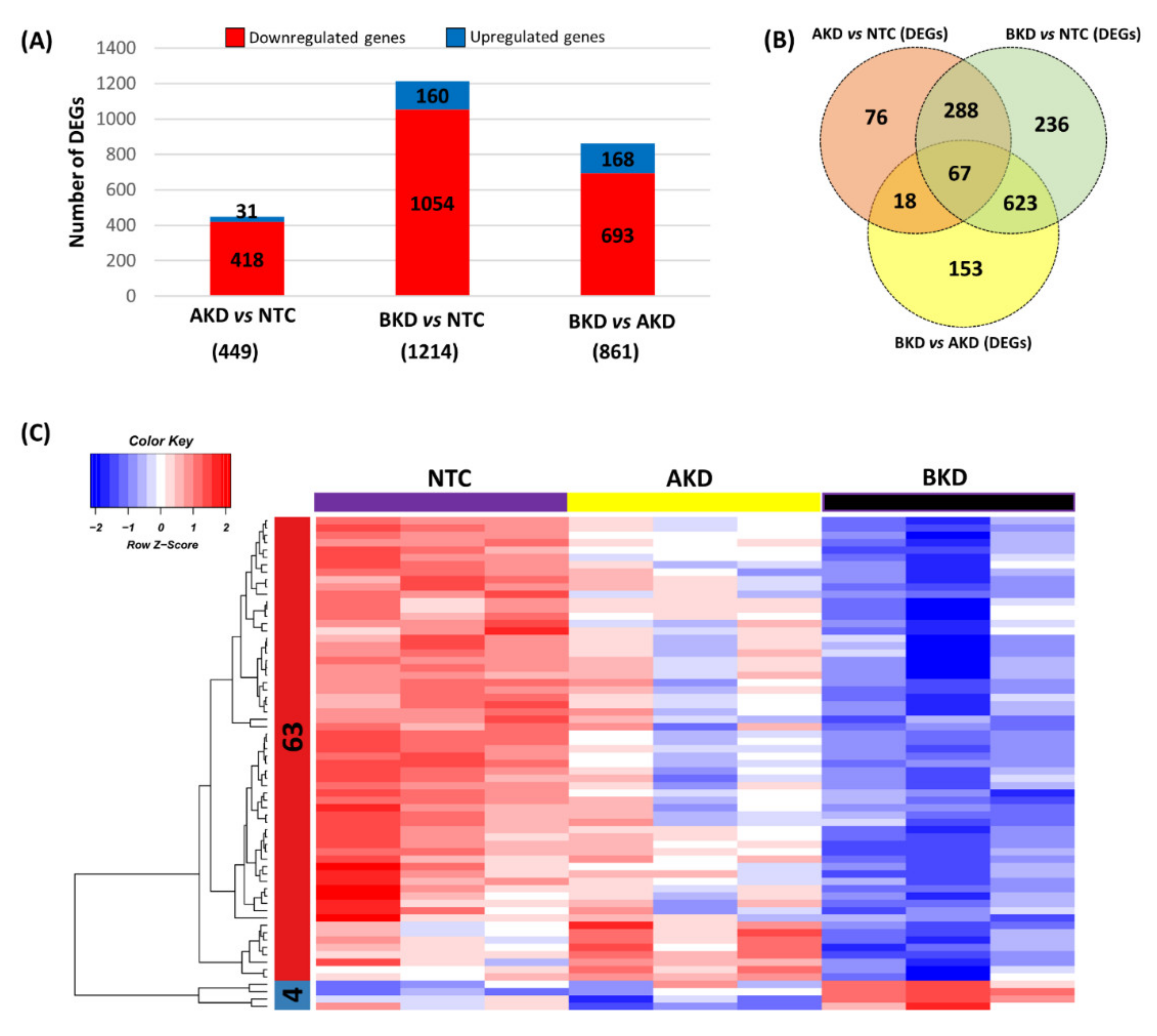
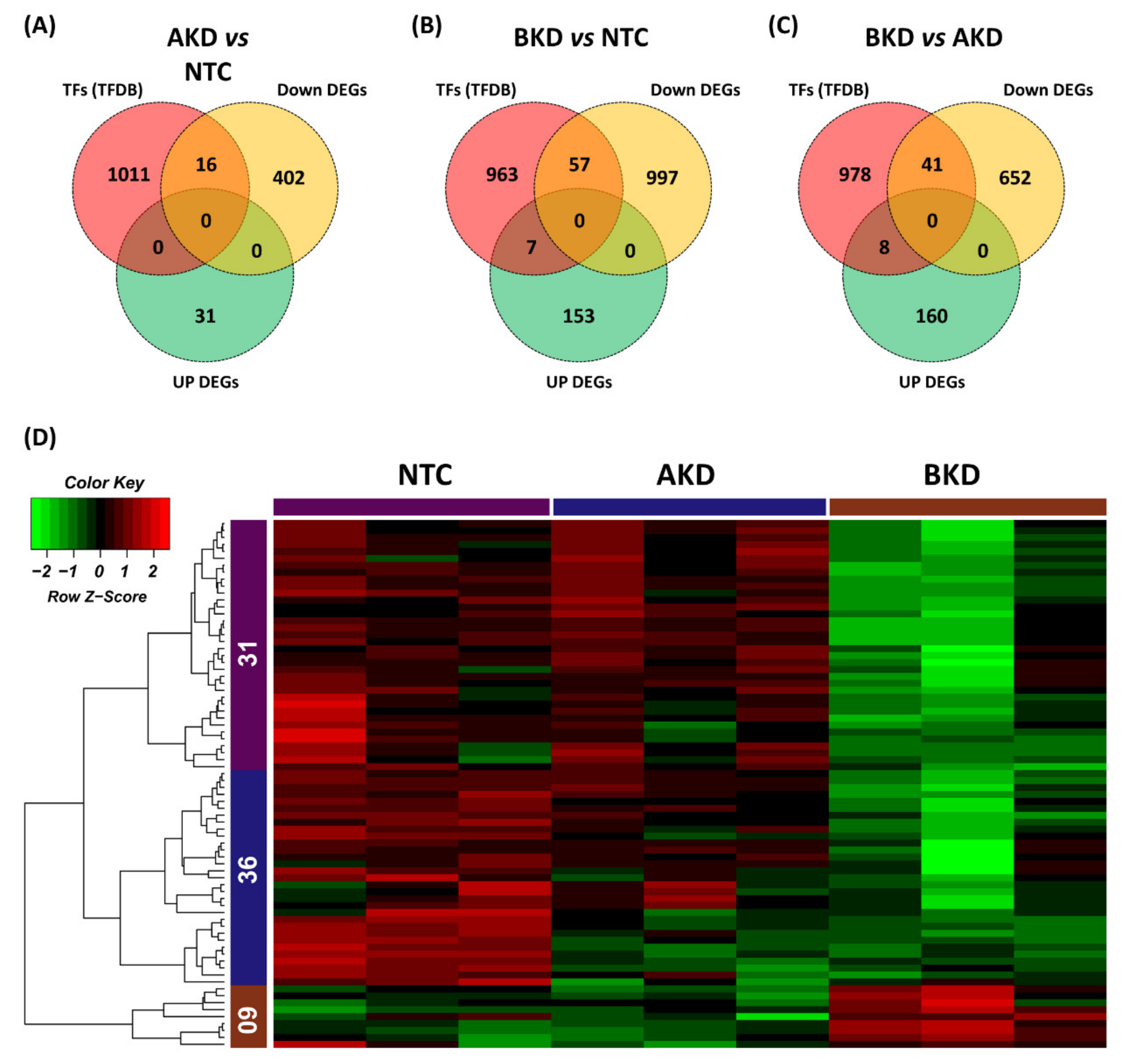
3.1. Differentially Expressed Genes
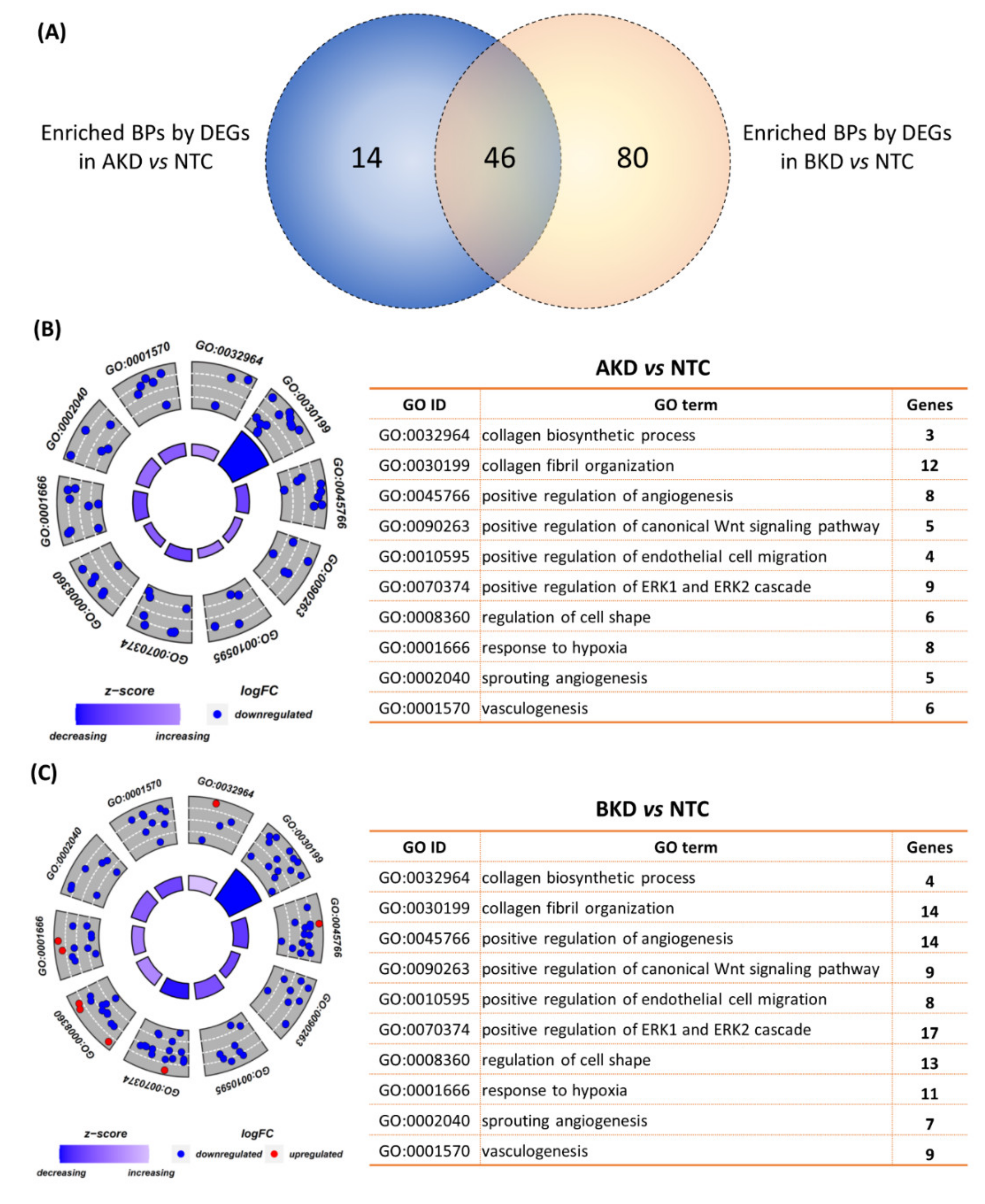
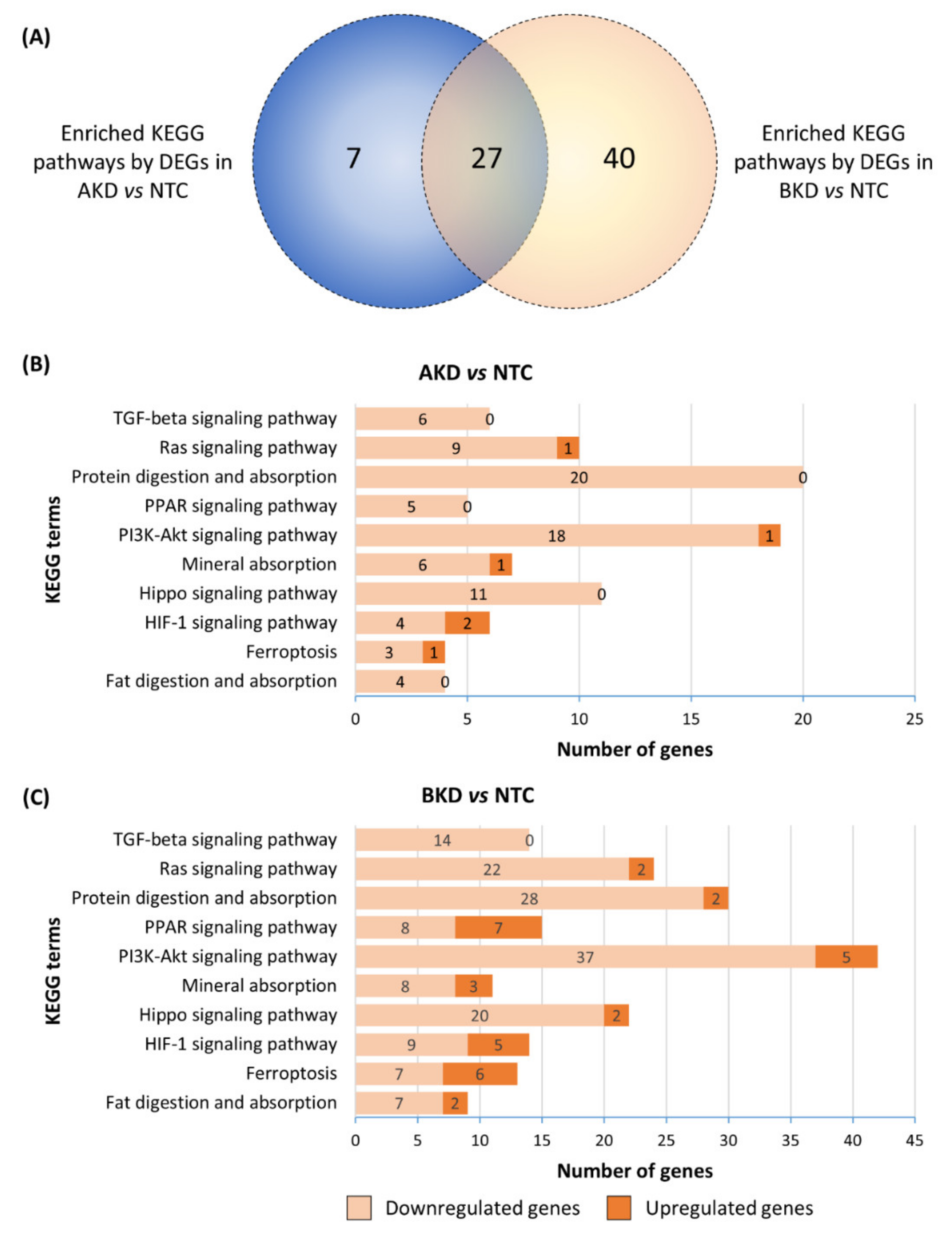
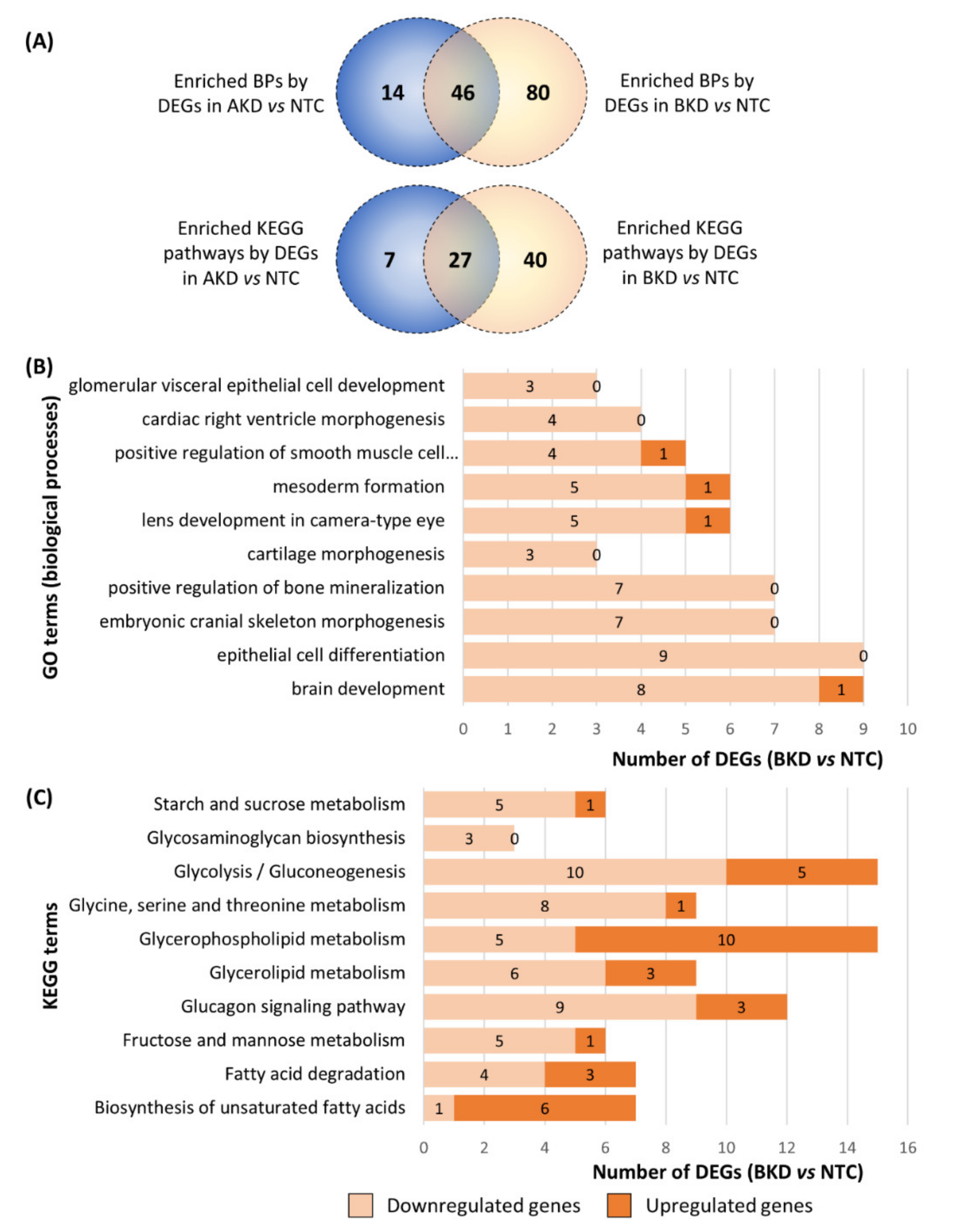
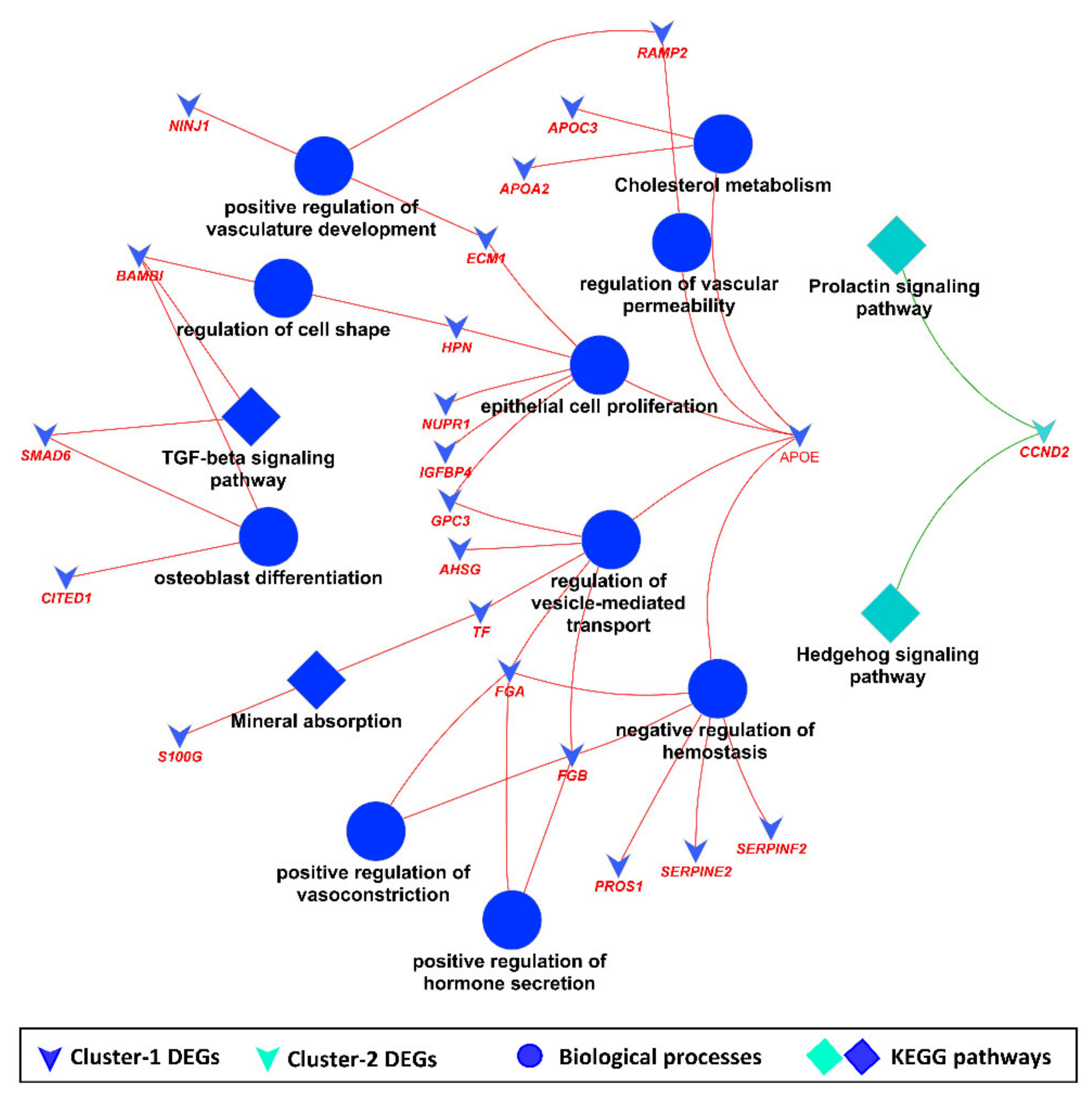
3.2. Downstream Enrichment Analysis of the DEGs
3.3. The Common DEGs in All Comparisons
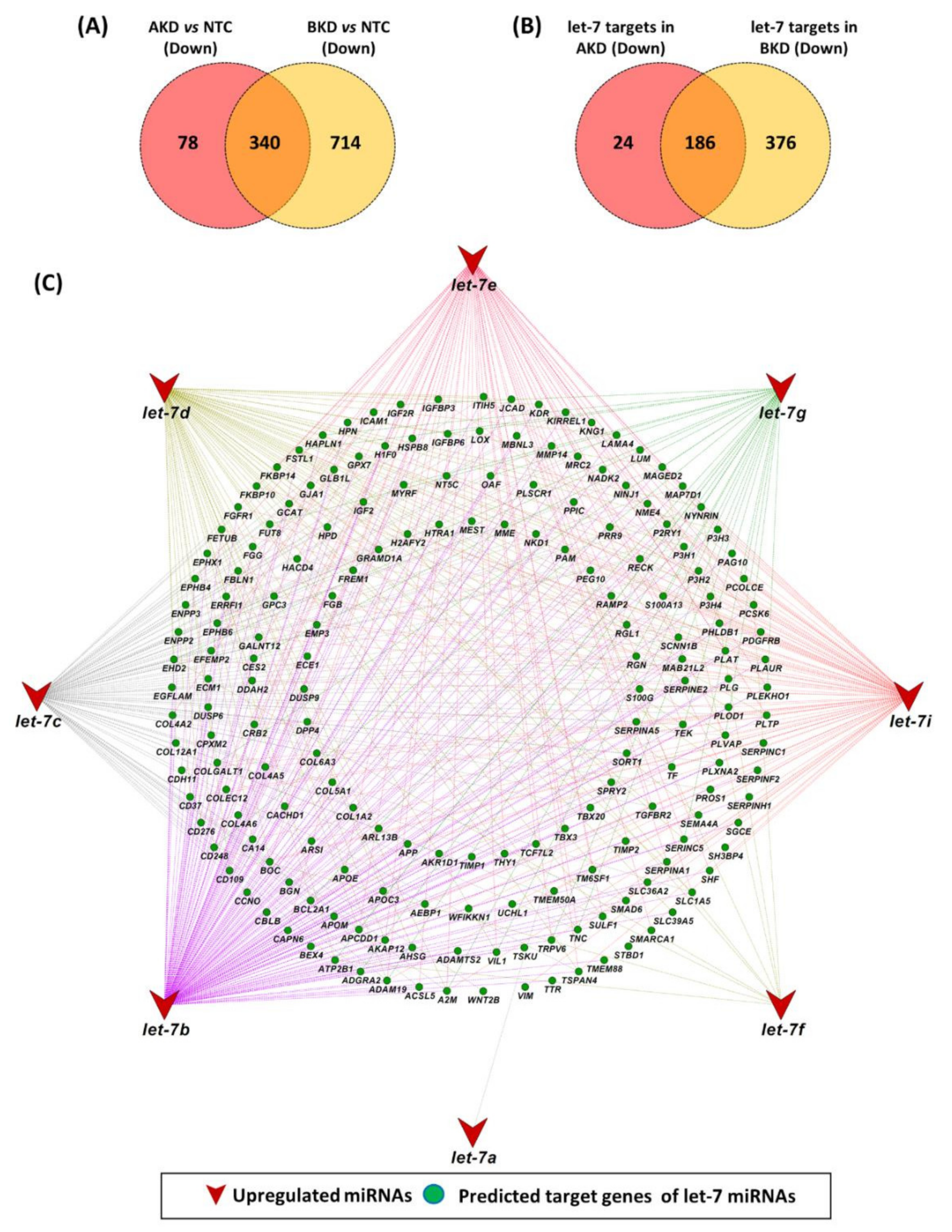
3.4. Let-7 miRNA Target Genes
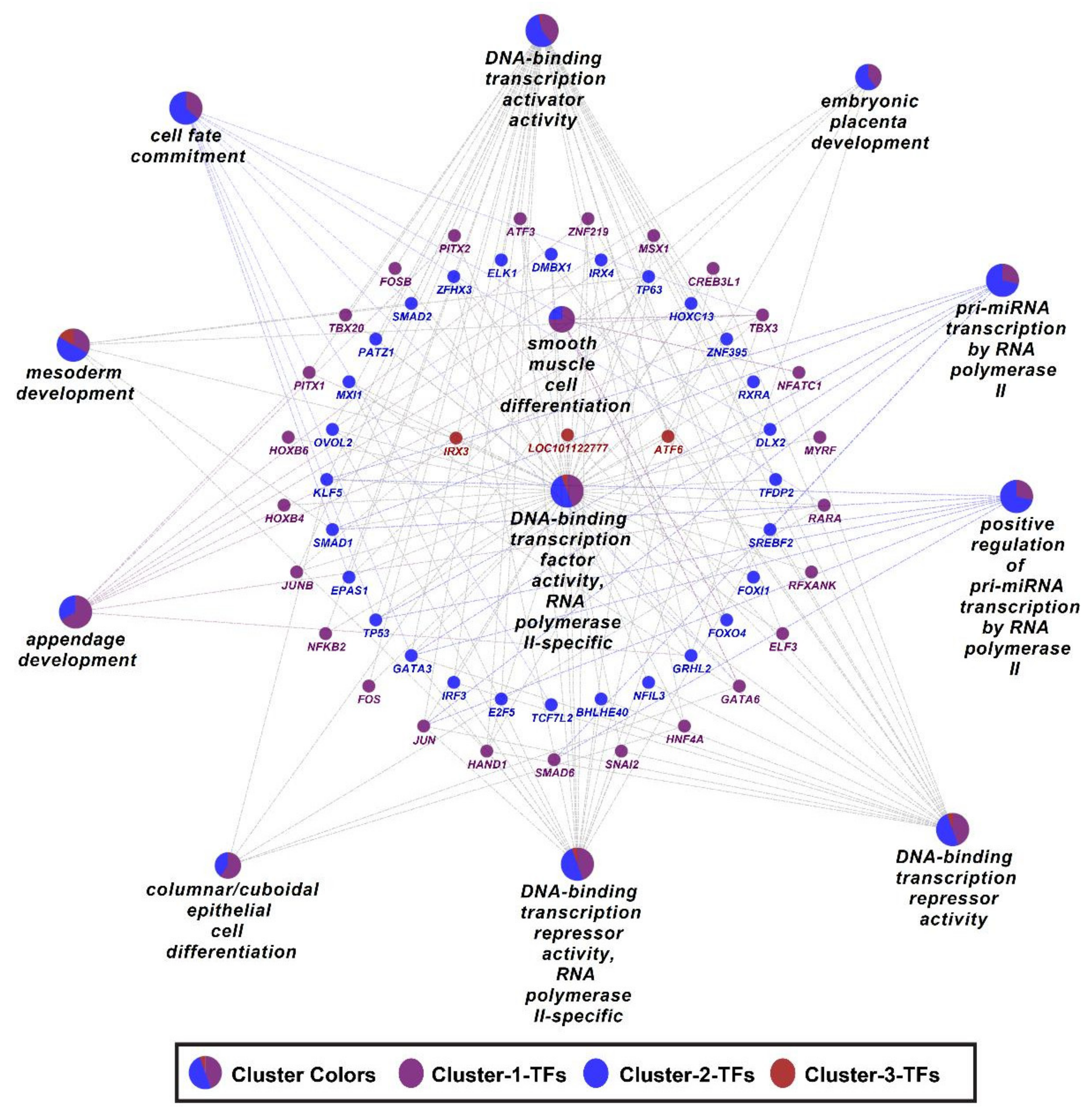
3.5. Differentially Expressed Transcription Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Brooks, K.; Burns, G.; Spencer, T.E. Conceptus elongation in ruminants: Roles of progesterone, prostaglandin, interferon tau and cortisol. J. Anim. Sci. Biotechnol. 2014, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wu, G.; Bazer, F.W. mTOR: The master regulator of conceptus development in response to uterine histotroph during pregnancy in ungulates. Mol. Med. mTOR 2016, 2016, 23–35. [Google Scholar]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Implantation mechanisms: Insights from the sheep. Reproduction 2004, 128, 657–668. [Google Scholar] [CrossRef]
- Wintenberger-Torres, S.; Flechon, J. Ultrastructural evolution of the trophoblast cells of the pre-implantation sheep blastocyst from day 8 to day 18. J. Anat. 1974, 118, 143. [Google Scholar] [PubMed]
- Wang, J.; Guillomot, M.; Hue, I. Cellular organization of the trophoblastic epithelium in elongating conceptuses of ruminants. Comptes Rendus Biol. 2009, 332, 986–997. [Google Scholar] [CrossRef]
- Rowson, L.; Moor, R. Development of the sheep conceptus during the first fourteen days. J. Anat. 1966, 100, 777. [Google Scholar]
- Wooding, F. The synepitheliochorial placenta of ruminants: Binucleate cell fusions and hormone production. Placenta 1992, 13, 101–113. [Google Scholar] [CrossRef]
- Guillomot, M.; Flechon, J.; Wintenberger-Torres, S. Conceptus attachment in the ewe: An ultrastructural study. Placenta 1981, 2, 169–182. [Google Scholar] [CrossRef]
- Wooding, P.; Burton, G. Comparative Placentation: Structures, Functions and Evolution; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Spencer, T.E.; Hansen, T.R. Implantation and establishment of pregnancy in ruminants. Regul. Implant. Establ. Pregnancy Mamm. 2015, 216, 105–135. [Google Scholar]
- Aires, M.; Dekagi, K.; Dantzer, V.; Yamada, A. Bovine placentome development during early pregnancy. Microscope 2014, 1, 390–396. [Google Scholar]
- Roberts, R.M.; Chen, Y.; Ezashi, T.; Walker, A.M. Interferons and the maternal–conceptus dialog in mammals. Semin. Cell Dev. Biol. 2008, 19, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thatcher, W.; Guzeloglu, A.; Mattos, R.; Binelli, M.; Hansen, T.; Pru, J. Uterine-conceptus interactions and reproductive failure in cattle. Theriogenology 2001, 56, 1435–1450. [Google Scholar] [CrossRef]
- Wooding, F.; Flint, A. Placentation. In Marshall’s Physiology of Reproduction; Lamming, G.H., Ed.; Springer: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Morgan, G.; Wooding, F.; Care, A.; Jones, G. Genetic regulation of placental function: A quantitative in situ hybridization study of calcium binding protein (calbindin-D9k) and calcium ATPase mRNAs in sheep placenta. Placenta 1997, 18, 211–218. [Google Scholar] [CrossRef]
- Roberts, R.; Ealy, A.; Alexenko, A.; Han, C.-S.; Ezashi, T. Trophoblast interferons. Placenta 1999, 20, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Godkin, J.; Bazer, F.; Moffatt, J.; Sessions, F.; Roberts, R. Purification and properties of a major, low molecular weight protein released by the trophoblast of sheep blastocysts at day 13–21. Reproduction 1982, 65, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Kappes, S.; Warren, W.; Pratt, S.; Liang, R.; Anthony, R. Quantification and cellular localization of ovine placental lactogen messenger ribonucleic acid expression during mid-and late gestation. Endocrinology 1992, 131, 2829–2838. [Google Scholar] [CrossRef]
- Ali, A.; Swanepoel, C.M.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Chorionic somatomammotropin RNA interference alters fetal liver glucose utilization. J. Endocrinol. 2020, 247, 169–180. [Google Scholar] [CrossRef]
- Tanner, A.R.; Lynch, C.S.; Kennedy, V.C.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction. Int. J. Mol. Sci. 2021, 22, 8150. [Google Scholar] [CrossRef]
- Sadovsky, Y.; Mouillet, J.-F.; Ouyang, Y.; Bayer, A.; Coyne, C.B. The function of trophomiRs and other microRNAs in the human placenta. Cold Spring Harb. Perspect. Med. 2015, 5, a023036. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Hadlich, F.; Abbas, M.W.; Iqbal, M.A.; Tesfaye, D.; Bouma, G.J.; Winger, Q.A.; Ponsuksili, S. MicroRNA–mRNA Networks in Pregnancy Complications: A Comprehensive Downstream Analysis of Potential Biomarkers. Int. J. Mol. Sci. 2021, 22, 2313. [Google Scholar] [CrossRef]
- Ali, A.; Anthony, R.V.; Bouma, G.J.; Winger, Q.A. LIN28-let-7 axis regulates genes in immortalized human trophoblast cells by targeting the ARID3B-complex. FASEB J. 2019, 33, 12348–12363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Stenglein, M.D.; Spencer, T.E.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. Trophectoderm-specific knockdown of Lin28 decreases expression of genes necessary for cell proliferation and reduces elongation of sheep conceptus. Int. J. Mol. Sci. 2020, 21, 2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. The role of LIN28-let-7-ARID3B pathway in placental development. Int. J. Mol. Sci. 2020, 21, 3637. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, T.-T.; Hsu, W.-H.; Ho, C.-H.; Hwang, W.-L.; Lan, H.-Y.; Lo, T.; Chang, C.-C.; Tai, S.-K.; Yang, M.-H. Let-7 modulates chromatin configuration and target gene repression through regulation of the ARID3B complex. Cell Rep. 2016, 14, 520–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, S.R.; Daley, G.Q. Lin28: A microRNA regulator with a macro role. Cell 2010, 140, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Chen, L.L.; Lei, X.X.; Yang, L.; Lin, H.; Carmichael, G.G.; Huang, Y. Genome-wide studies reveal that Lin28 enhances the translation of genes important for growth and survival of human embryonic stem cells. Stem Cells 2011, 29, 496–504. [Google Scholar] [CrossRef]
- Shyh-Chang, N.; Daley, G.Q. Lin28: Primal regulator of growth and metabolism in stem cells. Cell Stem Cell 2013, 12, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.M.; Goetzmann, L.N.; Cantlon, J.D.; Jeckel, K.M.; Winger, Q.A.; Anthony, R.V. Development of ovine chorionic somatomammotropin hormone-deficient pregnancies. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2016, 310, R837–R846. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Miao, Y.-R.; Jia, L.-H.; Yu, Q.-Y.; Zhang, Q.; Guo, A.-Y. AnimalTFDB 3.0: A comprehensive resource for annotation and prediction of animal transcription factors. Nucleic Acids Res. 2019, 47, D33–D38. [Google Scholar] [CrossRef]
- Rehmsmeier, M.; Steffen, P.; Höchsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: MicroRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S. gplots: Various R programming tools for plotting data. In R Package; Version 2; CRAN: Vienna, Austria, 2009; p. 1. [Google Scholar]
- Wang, T.; Han, P.; He, Y.; Zhao, C.; Wang, G.; Yang, W.; Shan, M.; Zhu, Y.; Yang, C.; Weng, M. Lin28A enhances chemosensitivity of colon cancer cells to 5-FU by promoting apoptosis in a let-7 independent manner. Tumor Biol. 2016, 37, 7657–7665. [Google Scholar] [CrossRef]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; LaPierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef] [Green Version]
- Hafner, M.; Max, K.E.; Bandaru, P.; Morozov, P.; Gerstberger, S.; Brown, M.; Molina, H.; Tuschl, T. Identification of mRNAs bound and regulated by human LIN28 proteins and molecular requirements for RNA recognition. RNA 2013, 19, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.-W.; Lai, Z.-Z.; Yang, H.-L.; Yang, S.-L.; Wang, C.-J.; Ao, D.; Ruan, L.-Y.; Shen, H.-H.; Zhou, W.-J.; Mei, J. Collagen at the maternal-fetal interface in human pregnancy. Int. J. Biol. Sci. 2020, 16, 2220. [Google Scholar] [CrossRef]
- Chen, D.b.; Zheng, J. Regulation of placental angiogenesis. Microcirculation 2014, 21, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, L.P.; Borowicz, P.P.; Vonnahme, K.A.; Johnson, M.L.; Grazul-Bilska, A.T.; Redmer, D.A.; Caton, J.S. Placental angiogenesis in sheep models of compromised pregnancy. J. Physiol. 2005, 565, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa-Shindo, Y.; Sakurai, T.; Kamiyoshi, A.; Kawate, H.; Iinuma, N.; Yoshizawa, T.; Koyama, T.; Fukuchi, J.; Iimuro, S.; Moriyama, N. The GPCR modulator protein RAMP2 is essential for angiogenesis and vascular integrity. J. Clin. Investig. 2008, 118, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial cell migration during angiogenesis. Circ. Res. 2007, 100, 782–794. [Google Scholar] [CrossRef]
- Norton, K.-A.; Popel, A.S. Effects of endothelial cell proliferation and migration rates in a computational model of sprouting angiogenesis. Sci. Rep. 2016, 6, 36992. [Google Scholar] [CrossRef]
- Rubinfeld, H.; Seger, R. The ERK cascade. Mol. Biotechnol. 2005, 31, 151–174. [Google Scholar] [CrossRef]
- Chambard, J.-C.; Lefloch, R.; Pouysségur, J.; Lenormand, P. ERK implication in cell cycle regulation. Biochim. Biophys. Acta BBA Mol. Cell Res. 2007, 1773, 1299–1310. [Google Scholar] [CrossRef]
- Shapiro, P.S.; Vaisberg, E.; Hunt, A.J.; Tolwinski, N.S.; Whalen, A.M.; McIntosh, J.R.; Ahn, N.G. Activation of the MKK/ERK pathway during somatic cell mitosis: Direct interactions of active ERK with kinetochores and regulation of the mitotic 3F3/2 phosphoantigen. J. Cell Biol. 1998, 142, 1533–1545. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, V.; Charron, J. Essential role of the ERK/MAPK pathway in blood-placental barrier formation. Development 2014, 141, 2825–2837. [Google Scholar] [CrossRef] [Green Version]
- Daoud, G.; Amyot, M.; Rassart, É.; Masse, A.; Simoneau, L.; Lafond, J. ERK1/2 and p38 regulate trophoblasts differentiation in human term placenta. J. Physiol. 2005, 566, 409–423. [Google Scholar] [CrossRef]
- Nayeem, S.B.; Arfuso, F.; Dharmarajan, A.; Keelan, J.A. Role of Wnt signalling in early pregnancy. Reprod. Fertil. Dev. 2016, 28, 525–544. [Google Scholar] [CrossRef]
- Huang, Z.; Li, S.; Fan, W.; Ma, Q. Transforming growth factor β1 promotes invasion of human JEG-3 trophoblast cells via TGF-β/Smad3 signaling pathway. Oncotarget 2017, 8, 33560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Fu, Z.; Hu, Y.; Li, Y.; Xu, Q.; Sun, D.; Tan, Y. Effects of transforming growth factor-β1 on the proliferation and invasion of the HTR-8/SVneo cell line. Oncol. Lett. 2014, 8, 2187–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Sivasubramaniyam, T.; Yinon, Y.; Tagliaferro, A.; Ray, J.; Nevo, O.; Post, M.; Caniggia, I. Aberrant TGFβ signaling contributes to altered trophoblast differentiation in preeclampsia. Endocrinology 2016, 157, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zheng, X.; Liu, J.; Zheng, R.; Yang, R.; Wang, Y.; Sun, L. PPAR signaling pathway in the first trimester placenta from in vitro fertilization and embryo transfer. Biomed. Pharmacother. 2019, 118, 109251. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Yang, H.; Ye, Y.; Ma, Z.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Makrigiannakis, A.; Jeschke, U.; von Schönfeldt, V. Role of peroxisome proliferator-activated receptors (PPARs) in trophoblast functions. Int. J. Mol. Sci. 2021, 22, 433. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, J.; Lei, Y.; Cong, C.; Tan, D.; Zhou, X. Research progress on the PI3K/AKT signaling pathway in gynecological cancer. Mol. Med. Rep. 2019, 19, 4529–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Cao, G.; Zhang, N.; Lou, T.; Wang, Q.; Zhang, Z.; Liu, C. RBP4 regulates trophoblastic cell proliferation and invasion via the PI3K/AKT signaling pathway. Mol. Med. Rep. 2018, 18, 2873–2879. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, X.-l.; Ma, C.-l.; Li, C.; Zhan, Y.; Li, W.-t.; Li, C.; Wang, Y.-h. STX2 Promotes Trophoblast Growth, Migration, and Invasion Through Activation of the PI3K-AKT Pathway in Preeclampsia. Front. Cell Dev. Biol. 2021, 9, 615973. [Google Scholar] [CrossRef]
- Highet, A.R.; Khoda, S.M.; Buckberry, S.; Leemaqz, S.; Bianco-Miotto, T.; Harrington, E.; Ricciardelli, C.; Roberts, C.T. Hypoxia induced HIF-1/HIF-2 activity alters trophoblast transcriptional regulation and promotes invasion. Eur. J. Cell Biol. 2015, 94, 589–602. [Google Scholar] [CrossRef]
- Albers, R.E.; Kaufman, M.R.; Natale, B.V.; Keoni, C.; Kulkarni-Datar, K.; Min, S.; Williams, C.R.; Natale, D.R.; Brown, T.L. Trophoblast-specific expression of Hif-1α results in preeclampsia-like symptoms and fetal growth restriction. Sci. Rep. 2019, 9, 2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boopathy, G.T.; Hong, W. Role of hippo pathway-YAP/TAZ signaling in angiogenesis. Front. Cell Dev. Biol. 2019, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Soncin, F.; Parast, M.M. Role of Hippo signaling pathway in early placental development. Proc. Natl. Acad. Sci. USA 2020, 117, 20354–20356. [Google Scholar] [CrossRef]
- Brett, K.E.; Ferraro, Z.M.; Yockell-Lelievre, J.; Gruslin, A.; Adamo, K.B. Maternal–fetal nutrient transport in pregnancy pathologies: The role of the placenta. Int. J. Mol. Sci. 2014, 15, 16153–16185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, W.W., Jr. Placental transport of nutrients to the fetus. Horm. Res. Paediatr. 1994, 42, 215–222. [Google Scholar]
- Hannan, N.J.; Stock, O.; Spencer, R.; Whitehead, C.; David, A.L.; Groom, K.; Petersen, S.; Henry, A.; Said, J.M.; Seeho, S. Circulating mRNAs are differentially expressed in pregnancies with severe placental insufficiency and at high risk of stillbirth. BMC Med. 2020, 18, 145. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.L.; Walker, S.P.; Ye, L.; Mendis, S.; Kaitu’u-Lino, T.u.J.; Lappas, M.; Tong, S. Placental specific mRNA in the maternal circulation are globally dysregulated in pregnancies complicated by fetal growth restriction. J. Clin. Endocrinol. Metab. 2013, 98, E429–E436. [Google Scholar] [CrossRef] [Green Version]
- Ng, E.K.; Tsui, N.B.; Lau, T.K.; Leung, T.N.; Chiu, R.W.; Panesar, N.S.; Lit, L.C.; Chan, K.-W.; Lo, Y.D. mRNA of placental origin is readily detectable in maternal plasma. Proc. Natl. Acad. Sci. USA 2003, 100, 4748–4753. [Google Scholar] [CrossRef] [Green Version]
- Paquette, A.G.; Chu, T.; Wu, X.; Wang, K.; Price, N.D.; Sadovsky, Y. Distinct communication patterns of trophoblastic miRNA among the maternal-placental-fetal compartments. Placenta 2018, 72, 28–35. [Google Scholar] [CrossRef]
- Chang, G.; Mouillet, J.F.; Mishima, T.; Chu, T.; Sadovsky, E.; Coyne, C.B.; Parks, W.T.; Surti, U.; Sadovsky, Y. Expression and trafficking of placental microRNAs at the feto-maternal interface. FASEB J. 2017, 31, 2760–2770. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Lu, J.; Wang, S.; Li, H.; Ge, Q.; Lu, Z. Application of next-generation sequencing technology to profile the circulating microRNAs in the serum of preeclampsia versus normal pregnant women. Clin. Chim. Acta 2011, 412, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Cai, Y. Down-regulation of microRNA let-7d inhibits the proliferation and invasion of trophoblast cells in preeclampsia. J. Cell. Biochem. 2018, 119, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.V.; Gusar, V.A.; Kan, N.E.; Prozorovskaya, K.N.; Karapetyan, A.O.; Bayev, O.R.; Chagovets, V.V.; Kliver, S.F.; Iakovishina, D.Y.; Frankevich, V.E. Identification of potential early biomarkers of preeclampsia. Placenta 2018, 61, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Hromadnikova, I.; Kotlabova, K.; Ivankova, K.; Krofta, L. First trimester screening of circulating C19MC microRNAs and the evaluation of their potential to predict the onset of preeclampsia and IUGR. PLoS ONE 2017, 12, e0171756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.L.; Jiang, X.; Rom, S. Let-7 microRNAs: Their Role in Cerebral and Cardiovascular Diseases, Inflammation, Cancer, and Their Regulation. Biomedicines 2021, 9, 606. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, W.; Yuan, Y.; Bai, Y.; Sun, Y.; Zhu, W.; Du, Z. MicroRNAs tend to synergistically control expression of genes encoding extensively-expressed proteins in humans. PeerJ 2017, 5, e3682. [Google Scholar] [CrossRef] [Green Version]
- Jens, M.; Rajewsky, N. Competition between target sites of regulators shapes post-transcriptional gene regulation. Nat. Rev. Genet. 2015, 16, 113–126. [Google Scholar] [CrossRef]
- Suter, D.M. Transcription factors and DNA play hide and seek. Trends Cell Biol. 2020, 30, 491–500. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Iqbal, M.A.; Abbas, M.W.; Bouma, G.J.; Anthony, R.V.; Spencer, T.E.; Winger, Q.A. Trophectoderm Transcriptome Analysis in LIN28 Knockdown Ovine Conceptuses Suggests Diverse Roles of the LIN28-let-7 Axis in Placental and Fetal Development. Cells 2022, 11, 1234. https://doi.org/10.3390/cells11071234
Ali A, Iqbal MA, Abbas MW, Bouma GJ, Anthony RV, Spencer TE, Winger QA. Trophectoderm Transcriptome Analysis in LIN28 Knockdown Ovine Conceptuses Suggests Diverse Roles of the LIN28-let-7 Axis in Placental and Fetal Development. Cells. 2022; 11(7):1234. https://doi.org/10.3390/cells11071234
Chicago/Turabian StyleAli, Asghar, Muhammad A. Iqbal, Muhammad W. Abbas, Gerrit J. Bouma, Russell V. Anthony, Thomas E. Spencer, and Quinton A. Winger. 2022. "Trophectoderm Transcriptome Analysis in LIN28 Knockdown Ovine Conceptuses Suggests Diverse Roles of the LIN28-let-7 Axis in Placental and Fetal Development" Cells 11, no. 7: 1234. https://doi.org/10.3390/cells11071234