
The Inhibitory Effect of Selected D2 Dopaminergic Receptor Agonists on VEGF-Dependent Neovascularization in Zebrafish Larvae: Potential New Therapy in Ophthalmic Diseases


Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Embryos Care and Maintenance
2.2. Drug Administration
2.3. Phenotype-Based Evaluation of Trunk and Retinal Vascularization
2.4. RNA Extraction, Reverse Transcription and qPCR Analysis
2.5. Statistical Analysis
3. Results
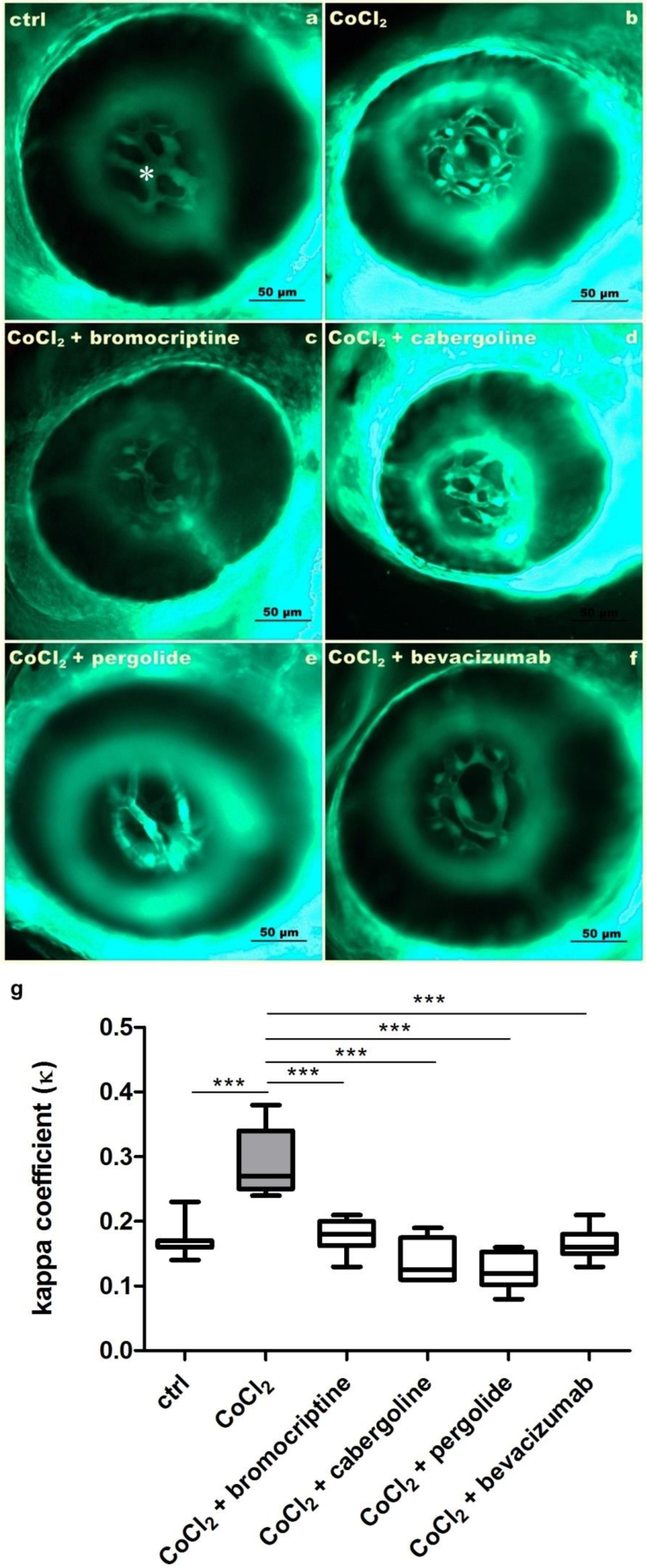
3.1. Inhibitory Effects of Selected D2 Dopaminergic Receptor Agonists towards CoCl2-Derived Increase of Hyaloid-Retinal Vessels
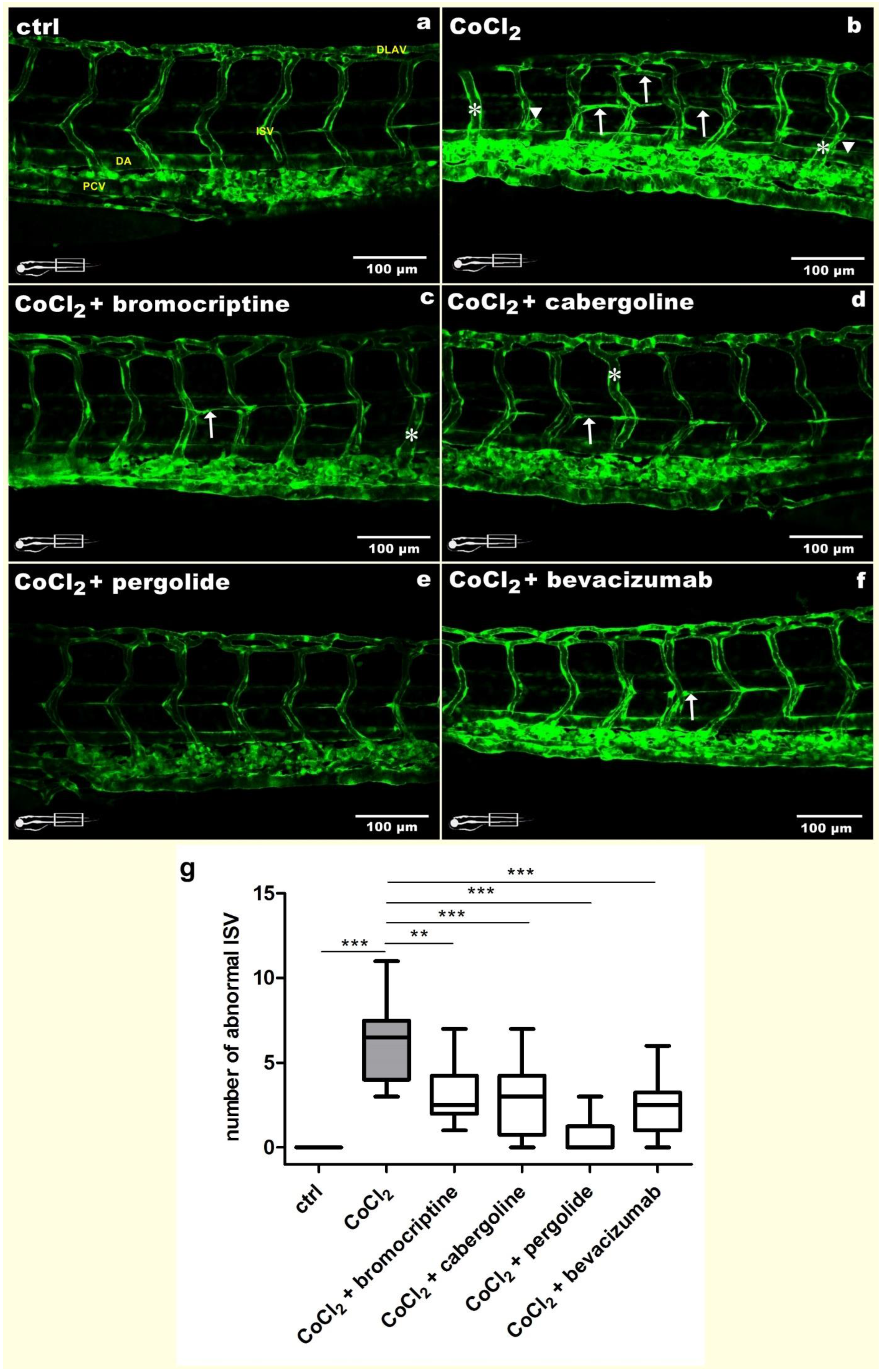
3.2. Ameliorative Effects of Selected D2 Dopaminergic Receptor Agonists towards CoCl2-Derived Trunk Blood Vessels Disruption
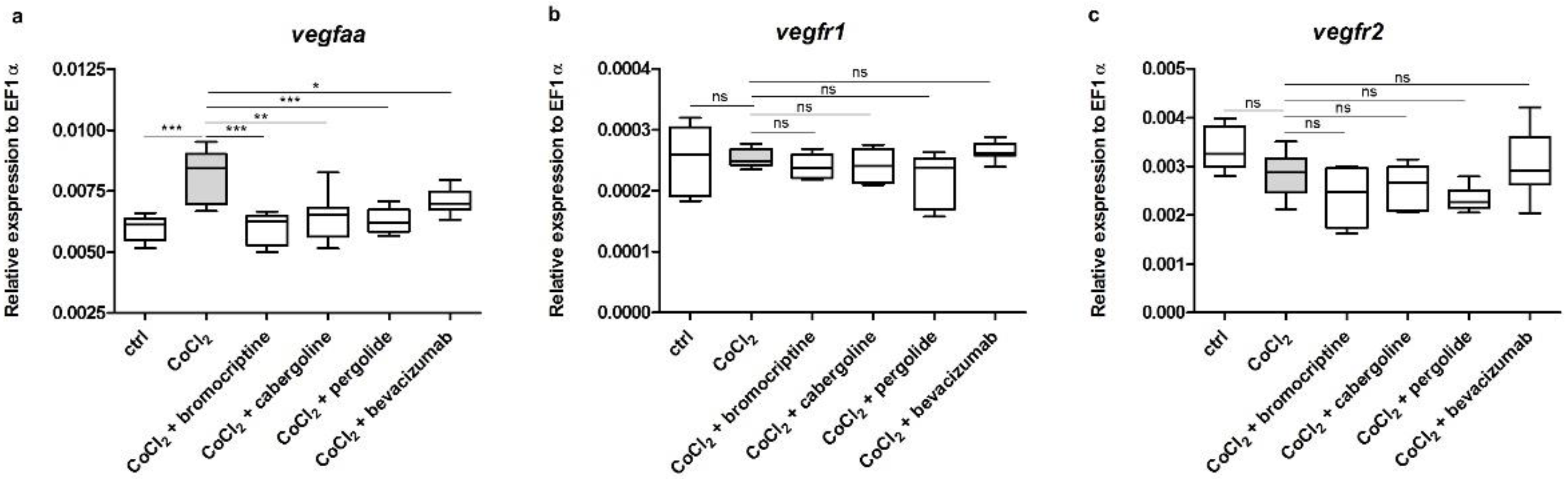
3.3. Effects of CoCl2 and Selected D2 Dopaminergic Agonists on mRNA Expression Level of vegfaa, vegfr1 and vegfr2
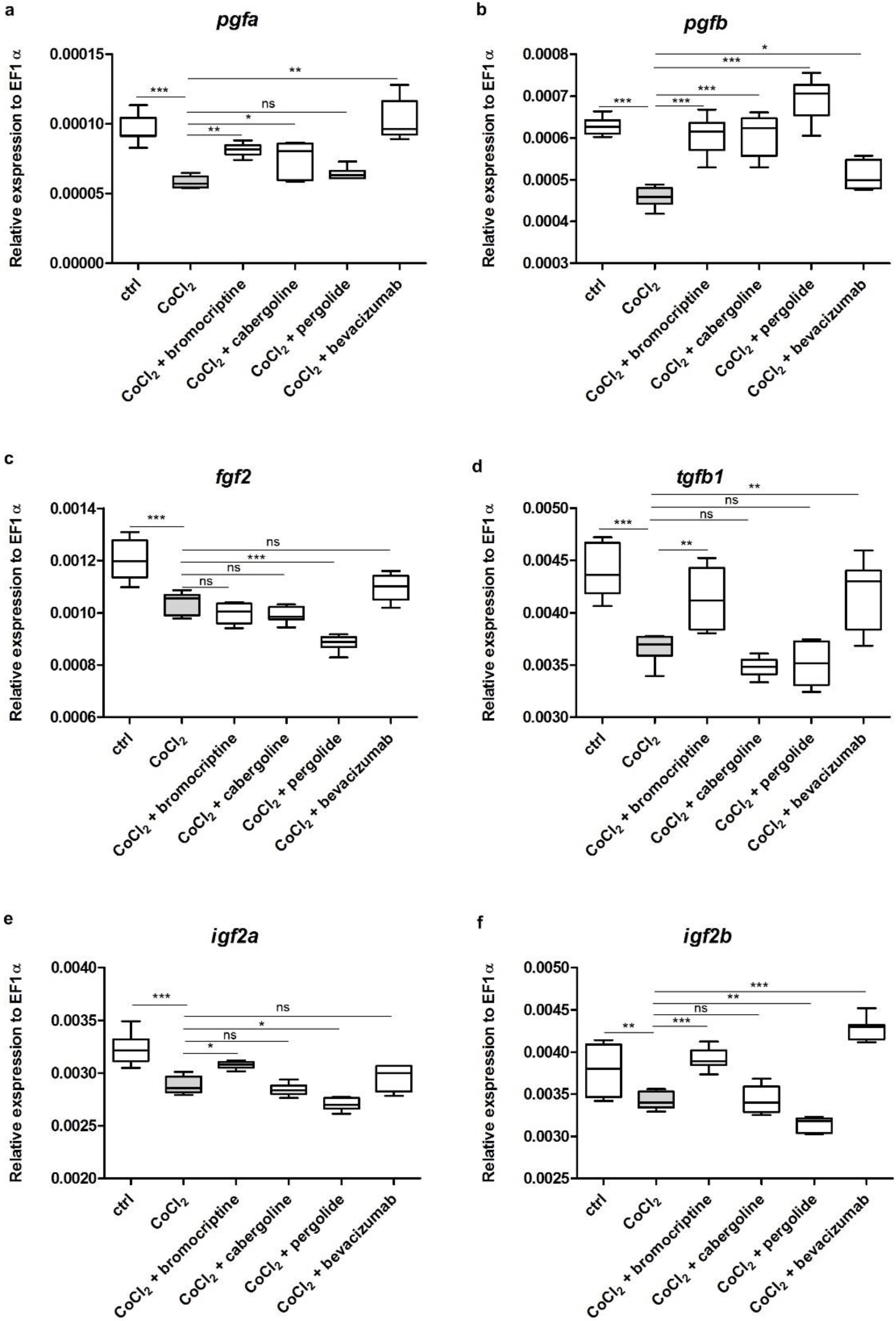
3.4. Effects of CoCl2 and Selected D2 Dopaminergic Agonists on mRNA Expression Level of Chosen Growth Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maniscalco, P.W.M.; D’Angio, C.T. Vascular endothelial growth factor. In Encyclopedia of Respiratory Medicine; Elsevier: Amsterdam, The Netherlands, 2006; pp. 413–418. [Google Scholar]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campochiaro, P.A. Molecular pathogenesis of retinal and choroidal vascular diseases. Prog. Retin. Eye Res. 2015, 49, 67–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartnett, M.E. Retinopathy of Prematurity: Evolving Treatment With Anti-Vascular Endothelial Growth Factor. Am. J. Ophthalmol. 2020, 218, 208–213, Erratum in Am. J. Ophthalmol. 2021, 231, 213. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.J.; Wan, L.; Lai, J.N.; Chen, C.S.; Chen, J.J.; Yen, W.M.; Chiu, L.T.; Hu, K.C.; Tien, P.T.; Lin, H.J. Increased risk of Parkinson’s disease among patients with age-related macular degeneration. BMC Ophthalmol. 2021, 21, 426. [Google Scholar] [CrossRef]
- Brilliant, M.H.; Vaziri, K.; Connor, T.B., Jr.; Schwartz, S.G.; Carroll, J.J.; McCarty, C.A.; Schrodi, S.J.; Hebbring, S.J.; Kishor, K.S.; Flynn, H.W., Jr.; et al. Mining Retrospective Data for Virtual Prospective Drug Repurposing: L-DOPA and Age-related Macular Degeneration. Am. J. Med. 2016, 129, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Mourad, S.M.; Wang, A.; Zhai, S.D.; Hart, R.J. Dopamine agonists for preventing ovarian hyperstimulation syndrome. Cochrane Database Syst. Rev. 2021, 4, CD008605. [Google Scholar] [CrossRef]
- Herbert, R.C.; Thompson, D.L.; Mitcham, P.B.; Lestelle, J.D.; Gilley, R.M.; Burns, P.J. Inhibitory Effects of Pergolide and Cabergoline Formulations on Daily Plasma Prolactin Concentrations in Geldings and on the Daily Prolactin Responses to a Small Dose of Sulpiride in Mares. J. Equine Vet. Sci. 2013, 33, 773–778. [Google Scholar] [CrossRef]
- Bonuccelli, U.; Colzi, A.; Del Dotto, P. Pergolide in the treatment of patients with early and advanced Parkinson’s disease. Clin. Neuropharmacol. 2002, 25, 1–10. [Google Scholar] [CrossRef]
- Chávez, M.N.; Aedo, G.; Fierro, F.A.; Allende, M.L.; Egaña, J.T. Zebrafish as an Emerging Model Organism to Study Angiogenesis in Development and Regeneration. Front. Physiol. 2016, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.W. Molecular and developmental biology of the hemangioblast. Dev. Dyn. 2008, 237, 1218–1231. [Google Scholar] [CrossRef] [Green Version]
- Eberlein, J.; Herdt, L.; Malchow, J.; Rittershaus, A.; Baumeister, S.; Helker, C.S. Molecular and Cellular Mechanisms of Vascular Development in Zebrafish. Life 2021, 11, 1088. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Kasica, N.; Jakubowski, P.; Kaleczyc, J. P-Glycoprotein Inhibitor Tariquidar Plays an Important Regulatory Role in Pigmentation in Larval Zebrafish. Cells 2021, 10, 690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, B.; Zhang, W.; Qian, Z.; Xiang, Y. Monitoring antiangiogenesis of bevacizumab in zebrafish. Drug Des. Dev. Ther. 2018, 12, 2423–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.C.; Chang, C.Y.; Kao, A.; Hsi, B.; Lee, S.H.; Chen, Y.H.; Wang, I.J. Hypoxia-induced retinal neovascularization in zebrafish embryos: A potential model of retinopathy of prematurity. PLoS ONE 2015, 10, e0126750. [Google Scholar] [CrossRef] [Green Version]
- Epstein, A.C.; Gleadle, J.M.; McNeill, L.A.; Hewitson, K.S.; O’Rourke, J.; Mole, D.R.; Mukherji, M.; Metzen, E.; Wilson, M.I.; Dhanda, A.; et al. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Hilliard, G.; Ferguson, T.; Millhorn, D.E. Cobalt inhibits the interaction between hypoxia-inducible factor-alpha and von Hippel-Lindau protein by direct binding to hypoxia-inducible factor-alpha. J. Biol. Chem. 2003, 278, 15911–15916. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef]
- Wu, J.; Ke, X.; Wang, W.; Zhang, H.; Ma, N.; Fu, W.; Zhao, M.; Gao, X.; Hao, X.; Zhang, Z. Aloe-emodin suppresses hypoxia-induced retinal angiogenesis via inhibition of HIF-1α/VEGF pathway. Int. J. Biol. Sci. 2016, 12, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, Y.; Astudillo, O.; Jensen, L.; Reynolds, A.L.; Waghorne, N.; Brazil, D.P.; Cao, Y.; O’Connor, J.J.; Kennedy, B.N. Selective inhibition of retinal angiogenesis by targeting PI3 kinase. PLoS ONE 2009, 4, e7867. [Google Scholar] [CrossRef] [Green Version]
- Chimote, G.; Sreenivasan, J.; Pawar, N.; Subramanian, J.; Sivaramakrishnan, H.; Sharma, S. Comparison of effects of anti-angiogenic agents in the zebrafish efficacy-toxicity model for translational anti-angiogenic drug discovery. Drug Des. Dev. Ther. 2014, 8, 1107–11023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakroborty, D.; Sarkar, C.; Mitra, R.B.; Banerjee, S.; Dasgupta, P.S.; Basu, S. Depleted dopamine in gastric cancer tissues: Dopamine treatment retards growth of gastric cancer by inhibiting angiogenesis. Clin. Cancer Res. 2004, 10, 4349–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Dasgupta, P.S. Decreased dopamine receptor expression and its second-messenger cAMP in malignant human colon tissue. Dig. Dis. Sci. 1999, 44, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Wick, M.M.; Kramer, R.A.; Gorman, M. Enhancement of L-dopa incorporation into melanoma by dopa decarboxylase inhibition. J. Investig. Dermatol. 1978, 70, 358–360. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, D.; Chowdhury, U.R.; Sarkar, C.; Baral, R.; Dasgupta, P.S.; Basu, S. Dopamine regulates endothelial progenitor cell mobilization from mouse bone marrow in tumor vascularization. J. Clin. Investig. 2008, 118, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Smith, M.; Lu, C.; Shahzad, M.M.; Pena, G.N.; Allen, J.K.; Stone, R.L.; Mangala, L.S.; Han, H.D.; Kim, H.S.; Farley, D.; et al. Dopamine blocks stress-mediated ovarian carcinoma growth. Clin. Cancer Res. 2011, 17, 3649–3659. [Google Scholar] [CrossRef] [Green Version]
- Senogles, S.E. D2 dopamine receptor-mediated antiproliferation in a small cell lung cancer cell line, NCI-H69. Anti-Cancer Drugs 2007, 18, 801–807. [Google Scholar] [CrossRef]
- Roy, S.; Lu, K.; Nayak, M.K.; Bhuniya, A.; Ghosh, T.; Kundu, S.; Ghosh, S.; Baral, R.; Dasgupta, P.S.; Basu, S. Activation of D2 Dopamine Receptors in CD133+ve Cancer Stem Cells in Non-small Cell Lung Carcinoma Inhibits Proliferation, Clonogenic Ability, and Invasiveness of These Cells. J. Biol. Chem. 2017, 292, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, D.; Sarkar, C.; Yu, H.; Wang, J.; Liu, Z.; Dasgupta, P.S.; Basu, S. Dopamine stabilizes tumor blood vessels by up-regulating angiopoietin 1 expression in pericytes and Kruppel-like factor-2 expression in tumor endothelial cells. Proc. Natl. Acad. Sci. USA 2011, 108, 20730–20735. [Google Scholar] [CrossRef] [Green Version]
- Chauvet, N.; Romanò, N.; Lafont, C.; Guillou, A.; Galibert, E.; Bonnefont, X.; Le Tissier, P.; Fedele, M.; Fusco, A.; Mollard, P.; et al. Complementary actions of dopamine D2 receptor agonist and anti-vegf therapy on tumoral vessel normalization in a transgenic mouse model. Int. J. Cancer 2017, 140, 2150–2161. [Google Scholar] [CrossRef] [Green Version]
- Ferrero, H.; García-Pascual, C.M.; Gómez, R.; Delgado-Rosas, F.; Cauli, O.; Simón, C.; Gaytán, F.; Pellicer, A. Dopamine receptor 2 activation inhibits ovarian vascular endothelial growth factor secretion in vitro: Implications for treatment of ovarian hyperstimulation syndrome with dopamine receptor 2 agonists. Fertil. Steril. 2014, 101, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Nagy, J.A.; Pal, S.; Vasile, E.; Eckelhoefer, I.A.; Bliss, V.S.; Manseau, E.J.; Dasgupta, P.S.; Dvorak, H.F.; Mukhopadhyay, D. The neurotransmitter dopamine inhibits angiogenesis induced by vascular permeability factor/vascular endothelial growth factor. Nat. Med. 2001, 7, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Chakroborty, D.; Mitra, R.B.; Banerjee, S.; Dasgupta, P.S.; Basu, S. Dopamine in vivo inhibits VEGF-induced phosphorylation of VEGFR-2, MAPK, and focal adhesion kinase in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1554–H1560. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Vohra, P.K.; Bhattacharya, R.; Dutta, S.; Sinha, S.; Mukhopadhyay, D. Dopamine regulates phosphorylation of VEGF receptor 2 by engaging Src-homology-2-domain-containing protein tyrosine phosphatase 2. J. Cell Sci. 2009, 122 Pt 18, 3385–3392. [Google Scholar] [CrossRef] [Green Version]
- Büchler, P.; Reber, H.A.; Büchler, M.W.; Friess, H.; Lavey, R.S.; Hines, O.J. Antiangiogenic activity of genistein in pancreatic carcinoma cells is mediated by the inhibition of hypoxia-inducible factor-1 and the down-regulation of VEGF gene expression. Cancer 2004, 100, 201–210. [Google Scholar] [CrossRef]
- Beckert, S.; Farrahi, F.; Perveen Ghani, Q.; Aslam, R.; Scheuenstuhl, H.; Coerper, S.; Königsrainer, A.; Hunt, T.K.; Hussain, M.Z. IGF-I-induced VEGF expression in HUVEC involves phosphorylation and inhibition of poly(ADP-ribose)polymerase. Biochem. Biophys. Res. Commun. 2006, 341, 67–72. [Google Scholar] [CrossRef]
- Narburgh, L.J.; Turner, J.; Freeman, S.J. Evaluation of the teratogenic potential of the dopamine agonist bromocriptine in rats. Toxicol. Lett. 1990, 50, 189–194. [Google Scholar] [CrossRef]
- Turkalj, I.; Braun, P.; Krupp, P. Surveillance of bromocriptine in pregnancy. JAMA 1982, 247, 1589–1591. [Google Scholar] [CrossRef]
- Caccavelli, L.; Cussac, D.; Pellegrini, I.; Audinot, V.; Jaquet, P.; Enjalbert, A. D2 dopaminergic receptors: Normal and abnormal transduction mechanisms. Horm. Res. 1992, 38, 78–83. [Google Scholar] [CrossRef]
- Stuttfeld, E.; Ballmer-Hofer, K. Structure and function of VEGF receptors. IUBMB Life 2009, 61, 915–922. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ema, M. Roles of VEGF-A signalling in development, regeneration, and tumours. J. Biochem. 2014, 156, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ober, E.A.; Olofsson, B.; Mäkinen, T.; Jin, S.W.; Shoji, W.; Koh, G.Y.; Alitalo, K.; Stainier, D.Y. Vegfc is required for vascular development and endoderm morphogenesis in zebrafish. EMBO Rep. 2004, 5, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzola, S.; Belleri, M.; Gariano, G.; Ribatti, D.; Costagliola, C.; Semeraro, F.; Presta, M. In vitro and ex vivo retina angiogenesis assays. Angiogenesis 2014, 17, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, A.; Dunk, C.; Jiang, J.; Shams, M.; Li, X.F.; Acevedo, C.; Weich, H.; Whittle, M.; Ahmed, A. Hypoxia down-regulates placenta growth factor, whereas fetal growth restriction up-regulates placenta growth factor expression: Molecular evidence for “placental hyperoxia” in intrauterine growth restriction. Lab. Investig. 1999, 79, 151–170. [Google Scholar]
- Poole, T.J.; Finkelstein, E.B.; Cox, C.M. The role of FGF and VEGF in angioblast induction and migration during vascular development. Dev. Dyn. 2001, 220, 1–17. [Google Scholar] [CrossRef]
- Jia, T.; Jacquet, T.; Dalonneau, F.; Coudert, P.; Vaganay, E.; Exbrayat-Héritier, C.; Vollaire, J.; Josserand, V.; Ruggiero, F.; Coll, J.L.; et al. FGF-2 promotes angiogenesis through a SRSF1/SRSF3/SRPK1-dependent axis that controls VEGFR1 splicing in endothelial cells. BMC Biol. 2021, 19, 173. [Google Scholar] [CrossRef]
- Javerzat, S.; Auguste, P.; Bikfalvi, A. The role of fibroblast growth factors in vascular development. Trends Mol. Med. 2002, 8, 483–489. [Google Scholar] [CrossRef]
- Conte, C.; Riant, E.; Toutain, C.; Pujol, F.; Arnal, J.F.; Lenfant, F.; Prats, A.C. FGF2 translationally induced by hypoxia is involved in negative and positive feedback loops with HIF-1alpha. PLoS ONE 2008, 3, e3078. [Google Scholar] [CrossRef]
- Kakudo, N.; Morimoto, N.; Ogawa, T.; Taketani, S.; Kusumoto, K. Hypoxia Enhances Proliferation of Human Adipose-Derived Stem Cells via HIF-1ɑ Activation. PLoS ONE 2015, 10, e0139890. [Google Scholar] [CrossRef] [Green Version]
- Mingyuan, X.; Qianqian, P.; Shengquan, X.; Chenyi, Y.; Rui, L.; Yichen, S.; Jinghong, X. Hypoxia-inducible factor-1α activates transforming growth factor-β1/Smad signaling and increases collagen deposition in dermal fibroblasts. Oncotarget 2017, 9, 3188–3197. [Google Scholar] [CrossRef] [Green Version]
- Marques, I.J.; Leito, J.T.; Spaink, H.P.; Testerink, J.; Jaspers, R.T.; Witte, F.; van den Berg, S.; Bagowski, C.P. Transcriptome analysis of the response to chronic constant hypoxia in zebrafish hearts. J. Comp. Physiol. B 2008, 178, 77–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheid, A.; Wenger, R.H.; Christina, H.; Camenisch, I.; Ferenc, A.; Stauffer, U.G.; Gassmann, M.; Meuli, M. Hypoxia-regulated gene expression in fetal wound regeneration and adult wound repair. Pediatr. Surg. Int. 2000, 16, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Khaidakov, M.; Szwedo, J.; Mitra, S.; Mehta, J.L. Angiostatic effects of aspirin in hypoxia-reoxygenation are linked to modulation of TGFβ1 signaling. J. Cardiovasc. Pharmacol. Ther. 2011, 16, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Eme, J.; Rhen, T.; Tate, K.B.; Gruchalla, K.; Kohl, Z.F.; Slay, C.E.; Crossley, D.A., 2nd. Plasticity of cardiovascular function in snapping turtle embryos (Chelydra serpentina): Chronic hypoxia alters autonomic regulation and gene expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R966–R979. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Kuno, S.; Mizuta, I.; Fujinami, A.; Matsui, H.; Ohta, M. Effects of dopamine agonists bromocriptine, pergolide, cabergoline, and SKF-38393 on GDNF, NGF, and BDNF synthesis in cultured mouse astrocytes. Life Sci. 2003, 73, 617–626. [Google Scholar] [CrossRef]
- Rezzola, S.; Paganini, G.; Semeraro, F.; Presta, M.; Tobia, C. Zebrafish (Danio rerio) embryo as a platform for the identification of novel angiogenesis inhibitors of retinal vascular diseases. Biochim. Biophys. Acta 2016, 1862, 1291–1296. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward 5′ -3′ | Reverse 5′-3′ | Accession No. |
|---|---|---|---|
| ef-1α | CTGGAGGCCAGCTCAAACAT | ATCAAGAAGAGTAGTACCGCTAGCATTAC | NM_131263.1 |
| vegfaa | CTGCTGGTAGACATCATC | TTTCGTGTCTCTGTCGGG | XM_009292018.3, NM_131408.3, NM_001110349.2 |
| vegfr1 | AGCCACAGACAGGAAGTGTT | GACACAGCGATAGATGCCAGA | NM_001014829.3 |
| vegfr2 | CTGCTGGTAGACATCATC | TTTCGTGTCTCTGTCGGG | XM_009291142.3 |
| pgfa | GCTGCTGCAACGACGAAAAA | CATAATCTCTGCGCCGCTCT | XM_021468740.1 |
| pgfb | ACCTACAACAAAACAAGACAGATGG | GAGACAGCGTTTACCTGCGG | XM_017351845.2 |
| fgf2 | GCATCTGTACCAACCGTTTCC | ATCTGTGGTCCTTTTCGTCCC | NM_212823.2 |
| tgfb1 | TGTACCCGCAATCCTTGACC | GGACAATTGCTCCACCTTGTG | NM_182873.1 |
| igf2a | ACAGGCTCTTCACAAGGACAC | TTCGGGCCAACAGAATGGAT | NM_131433.1 |
| igf2b | CGCATTAAAACAGGAGGTCCC | CTGAGCAGCCTTTCTTTGCC | NM_001001815.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasica, N.; Święch, A.; Saładziak, K.; Mackiewicz, J.; Osęka, M. The Inhibitory Effect of Selected D2 Dopaminergic Receptor Agonists on VEGF-Dependent Neovascularization in Zebrafish Larvae: Potential New Therapy in Ophthalmic Diseases. Cells 2022, 11, 1202. https://doi.org/10.3390/cells11071202
Kasica N, Święch A, Saładziak K, Mackiewicz J, Osęka M. The Inhibitory Effect of Selected D2 Dopaminergic Receptor Agonists on VEGF-Dependent Neovascularization in Zebrafish Larvae: Potential New Therapy in Ophthalmic Diseases. Cells. 2022; 11(7):1202. https://doi.org/10.3390/cells11071202
Chicago/Turabian StyleKasica, Natalia, Anna Święch, Katarzyna Saładziak, Jerzy Mackiewicz, and Maciej Osęka. 2022. "The Inhibitory Effect of Selected D2 Dopaminergic Receptor Agonists on VEGF-Dependent Neovascularization in Zebrafish Larvae: Potential New Therapy in Ophthalmic Diseases" Cells 11, no. 7: 1202. https://doi.org/10.3390/cells11071202