
The Mysterious Actor—γδ T Lymphocytes in Chronic Lymphocytic Leukaemia (CLL)


{kind=link}
{kind=link}
Abstract
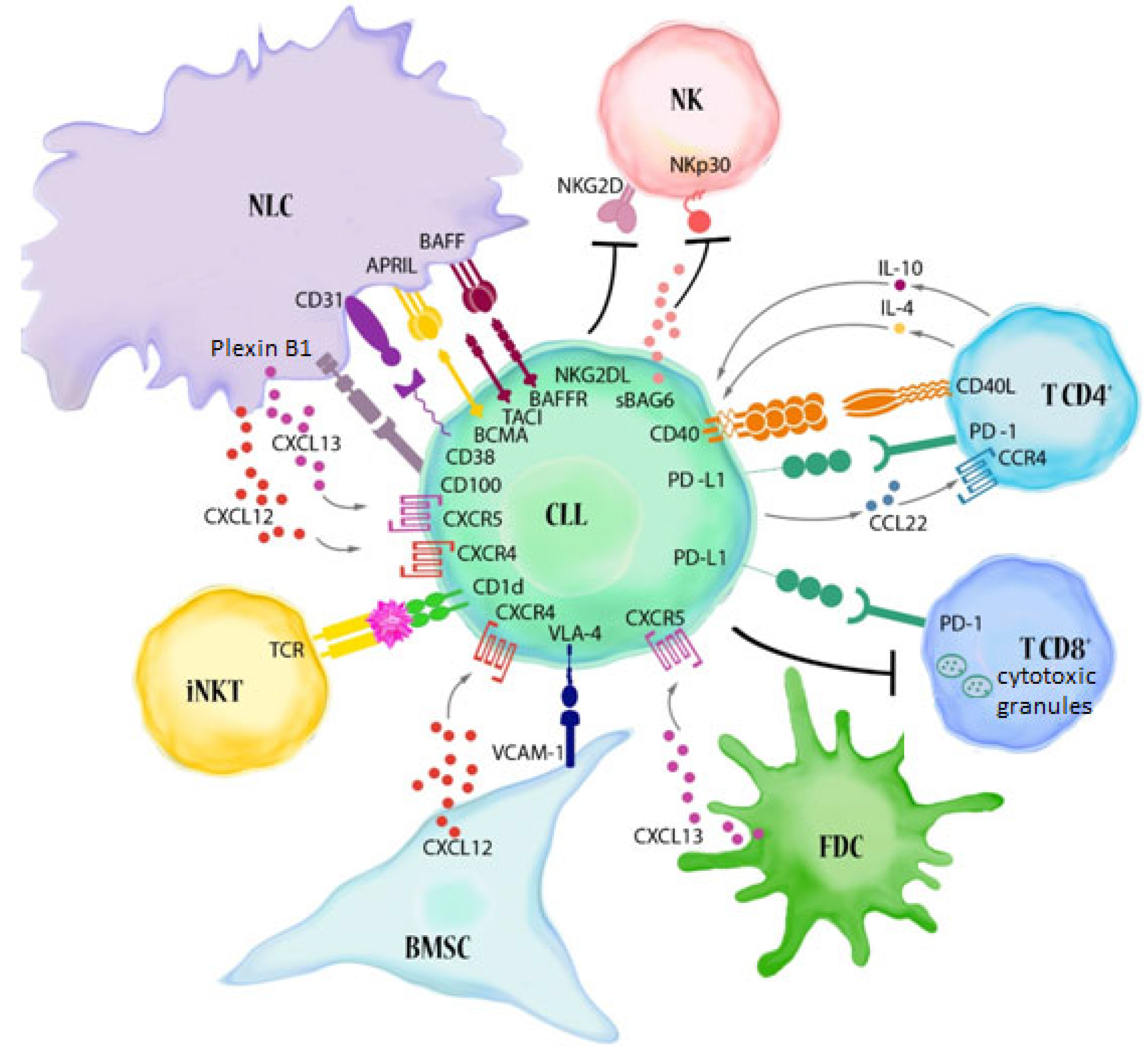
:1. Chronic Lymphocytic Leukaemia
2. γδ T Cells in Human
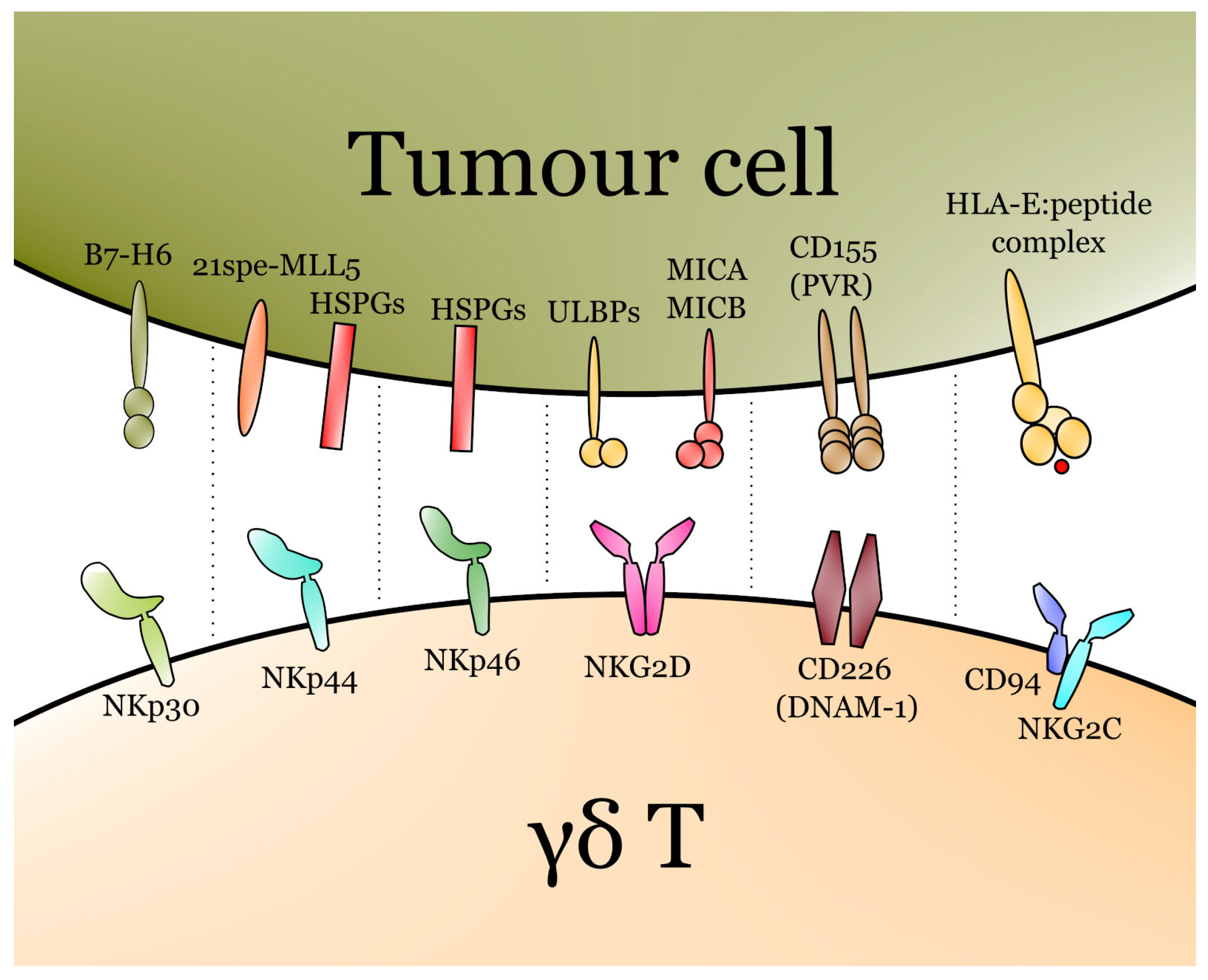
3. γδ T Recognition of Tumour Cells
4. Vδ1 γδ T Cells Are Expanded in the Peripheral Blood of CLL Patients
5. Vδ1 Cells Are More Cytotoxic towards CLL Clones Than Vδ2
6. Vδ1 Cells for the Cellular Immunotherapy of CLL
7. Vδ2 in CLL Patients Are Dysfunctional
8. Poor response of Vδ2 to Phosphoantigen Stimulation Has a Negative Prognostic Value in CLL
9. Effect of Ibrutinib on γδ T Cells
10. Perspectives for the Use of γδ T Cells in CLL Immunotherapy
11. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2B4 | natural killer cell receptor 2B4: CD244 |
| BCR | B cell receptor |
| CAR | chimeric antigen receptor |
| CTLA-4 | cytotoxic T lymphocyte-associated protein 4 |
| DNAM-1 | DNAX accessory molecule-1 |
| EBV | Epstein–Barr virus |
| FasL | Fas ligand |
| HMB-PP | (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate |
| KIR2DL | killer cell immunoglobulin-like receptor 2DL |
| LAG-3 | lymphocyte activation gene 3 |
| LDH | lactate dehydrogenase |
| MICA | MHC class I chain-related protein A |
| MICB | MHC class I chain-related protein B |
| MLL5 | mixed lineage leukaemia 5 |
| NCRs | natural cytotoxicity receptors |
| NKG2A | natural killer group 2A |
| NKG2C | natural killer group 2C |
| NKG2D | natural killer group 2D |
| NKp30 | natural cytotoxicity triggering receptor 3 |
| NKp44 | natural cytotoxicity triggering receptor 2 |
| PHA | phytohaemagglutinin |
| PD-1 | programmed cell death protein 1 |
| PD-L1 | programmed death-ligand 1 |
| PVR | polio virus receptor |
| TCR | T cell receptor |
| TIGIT | T cell immunoreceptor with Ig and ITIM domains |
| TIM-3 | T cell immunoglobulin and mucin domain-containing-3 |
| TRAIL | TNF-related apoptosis-inducing ligand |
| ULBPs | UL16-binding proteins |
| ZAP-70 | zeta-chain-associated protein kinase 70 |
References
- Borges, F.C.; Ramos, A.; Lourenço, A.; da Silva, M.G.; Miranda, A. Detailing the Epidemiological and Clinical Characteristics of Chronic Lymphocytic Leukaemia in Portugal—Results from a Population-Based Cancer Registry Cohort Study. PLoS ONE 2021, 16, e0258423. [Google Scholar] [CrossRef]
- Alshemmari, S.H.; Hamdah, A.; Pandita, R.; Kunhikrishnan, A. Chronic Lymphocytic Leukemia in a Young Population. Leuk. Res. 2021, 110, 106668. [Google Scholar] [CrossRef] [PubMed]
- Molica, S. Sex Differences in Incidence and Outcome of Chronic Lymphocytic Leukemia Patients. Leuk. Lymphoma 2006, 47, 1477–1480. [Google Scholar] [CrossRef] [PubMed]
- Morton, L.M.; Wang, S.S.; Devesa, S.S.; Hartge, P.; Weisenburger, D.D.; Linet, M.S. Lymphoma Incidence Patterns by WHO Subtype in the United States, 1992–2001. Blood 2006, 107, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Rozman, C.; Montserrat, E. Chronic Lymphocytic Leukemia. N. Engl. J. Med. 1995, 333, 1052–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weil, C.; Chodick, G.; Shalev, V.; Kan, I.; Afik, R.; Cohen, R.; Sail, K.; Herishanu, Y. Epidemiology, Longitudinal Treatment Patterns and Outcomes of Chronic Lymphocytic Leukemia in Israel. Leuk. Lymphoma 2021, 62, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Defrancesco, I.; Frigeni, M.; Arcaini, L. Low-Grade Lymphomas (Other than CLL/SLL) in Older Patients. In Geriatric Oncology; Springer: Cham, Switzerland, 2020; pp. 567–581. ISBN 9783319574158. [Google Scholar]
- Watson, L.; Wyld, P.; Catovsky, D. Disease Burden of Chronic Lymphocytic Leukaemia within the European. Union. Eur. J. Haematol. 2008, 81, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Pflug, N.; Bahlo, J.; Shanafelt, T.D.; Eichhorst, B.F.; Bergmann, M.A.; Elter, T.; Bauer, K.; Malchau, G.; Rabe, K.G.; Stilgenbauer, S.; et al. Development of a Comprehensive Prognostic Index for Patients with Chronic Lymphocytic Leukemia. Blood 2014, 124, 49–62. [Google Scholar] [CrossRef]
- Bewarder, M.; Stilgenbauer, S.; Thurner, L.; Kaddu-Mulindwa, D. Current Treatment Options in CLL. Cancers 2021, 13, 2468. [Google Scholar] [CrossRef]
- Pérez-Carretero, C.; González-Gascón-y-Marín, I.; Rodríguez-Vicente, A.E.; Quijada-Álamo, M.; Hernández-Rivas, J.-Á.; Hernández-Sánchez, M.; Hernández-Rivas, J.M. The Evolving Landscape of Chronic Lymphocytic Leukemia on Diagnosis, Prognosis and Treatment. Diagnostics 2021, 11, 853. [Google Scholar] [CrossRef]
- González-Gascón-y-Marín, I.; Muñoz-Novas, C.; Rodríguez-Vicente, A.-E.; Quijada-Álamo, M.; Hernández-Sánchez, M.; Pérez-Carretero, C.; Ramos-Ascanio, V.; Hernández-Rivas, J.-Á. From Biomarkers to Models in the Changing Landscape of Chronic Lymphocytic Leukemia: Evolve or Become Extinct. Cancers 2021, 13, 1782. [Google Scholar] [CrossRef] [PubMed]
- Hotinski, A.K.; Best, O.G.; Kuss, B.J. The Future of Laboratory Testing in Chronic Lymphocytic Leukaemia. Pathology 2021, 53, 377–384. [Google Scholar] [CrossRef]
- Griggio, V.; Perutelli, F.; Salvetti, C.; Boccellato, E.; Boccadoro, M.; Vitale, C.; Coscia, M. Immune Dysfunctions and Immune-Based Therapeutic Interventions in Chronic Lymphocytic Leukemia. Front. Immunol. 2020, 11, 2313. [Google Scholar] [CrossRef] [PubMed]
- Ten Hacken, E.; Burger, J.A. Microenvironment Interactions and B-Cell Receptor Signaling in Chronic Lymphocytic Leukemia: Implications for Disease Pathogenesis and Treatment. Biochim. Biophys. Acta 2016, 1863, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.; Muñoz, C.; Terol, M.J.; Hernández-Rivas, J.-Á.; Villanueva, M. Restoration of the Immune Function as a Complementary Strategy to Treat Chronic Lymphocytic Leukemia Effectively. J. Exp. Clin. Cancer Res. 2021, 40, 321. [Google Scholar] [CrossRef]
- Zarobkiewicz, M.; Kowalska, W.; Chocholska, S.; Tomczak, W.; Szymańska, A.; Morawska, I.; Wojciechowska, A.; Bojarska-Junak, A. High M-MDSC Percentage as a Negative Prognostic Factor in Chronic Lymphocytic Leukaemia. Cancers 2020, 12, 2614. [Google Scholar] [CrossRef]
- Mohr, A.; Cumin, M.; Bagacean, C.; Pochard, P.; Le Dantec, C.; Hillion, S.; Renaudineau, Y.; Berthou, C.; Tempescul, A.; Saad, H.; et al. The Regulatory Capacity of B Cells Directs the Aggressiveness of CLL. OncoImmunology 2019, 8, 1554968. [Google Scholar] [CrossRef] [Green Version]
- Forconi, F.; Moss, P. Perturbation of the Normal Immune System in Patients with CLL. Blood 2015, 126, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Mhibik, M.; Wiestner, A.; Sun, C. Harnessing the Effects of BTKi on T Cells for Effective Immunotherapy against CLL. Int. J. Mol. Sci. 2020, 21, 68. [Google Scholar] [CrossRef] [Green Version]
- Peters, F.S.; Strefford, J.C.; Eldering, E.; Kater, A.P. T-Cell Dysfunction in Chronic Lymphocytic Leukemia from an Epigenetic Perspective. Haematologica 2021, 106, 1234–1243. [Google Scholar] [CrossRef]
- Burger, J.A.; Gribben, J.G. The Microenvironment in Chronic Lymphocytic Leukemia (CLL) and Other B Cell Malignancies: Insight into Disease Biology and New Targeted Therapies. Semin. Cancer Biol. 2014, 24, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Svanberg, R.; Janum, S.; Patten, P.E.M.; Ramsay, A.G.; Niemann, C.U. Targeting the Tumor Microenvironment in Chronic Lymphocytic Leukemia. Haematologica 2020, 106, 2312. [Google Scholar] [CrossRef] [PubMed]
- Vlachonikola, E.; Stamatopoulos, K.; Chatzidimitriou, A. T Cell Defects and Immunotherapy in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 3255. [Google Scholar] [CrossRef] [PubMed]
- Sportoletti, P.; De Falco, F.; Del Papa, B.; Baldoni, S.; Guarente, V.; Marra, A.; Dorillo, E.; Rompietti, C.; Adamo, F.M.; Ruggeri, L.; et al. NK Cells in Chronic Lymphocytic Leukemia and Their Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 6665. [Google Scholar] [CrossRef] [PubMed]
- Deaglio, S.; Vaisitti, T.; Bergui, L.; Bonello, L.; Horenstein, A.L.; Tamagnone, L.; Boumsell, L.; Malavasi, F. CD38 and CD100 Lead a Network of Surface Receptors Relaying Positive Signals for B-CLL Growth and Survival. Blood 2005, 105, 3042–3050. [Google Scholar] [CrossRef]
- Roessner, P.M.; Seiffert, M. T-Cells in Chronic Lymphocytic Leukemia: Guardians or Drivers of Disease? Leukemia 2020, 34, 2012–2024. [Google Scholar] [CrossRef]
- Bond, D.A.; Woyach, J.A. Targeting BTK in CLL: Beyond Ibrutinib. Curr. Hematol. Malig. Rep. 2019, 14, 197–205. [Google Scholar] [CrossRef]
- Yosifov, D.Y.; Wolf, C.; Stilgenbauer, S.; Mertens, D. From Biology to Therapy: The CLL Success Story. HemaSphere 2019, 3, e175. [Google Scholar] [CrossRef]
- Fraietta, J.A.; Lacey, S.F.; Orlando, E.J.; Pruteanu-Malinici, I.; Gohil, M.; Lundh, S.; Boesteanu, A.C.; Wang, Y.; O’Connor, R.S.; Hwang, W.-T.; et al. Determinants of Response and Resistance to CD19 Chimeric Antigen Receptor (CAR) T Cell Therapy of Chronic Lymphocytic Leukemia. Nat. Med. 2018, 24, 563–571. [Google Scholar] [CrossRef]
- Lorentzen, C.L.; Straten, P.T. CD19-Chimeric Antigen Receptor T Cells for Treatment of Chronic Lymphocytic Leukaemia and Acute Lymphoblastic Leukaemia. Scand. J. Immunol. 2015, 82, 307–319. [Google Scholar] [CrossRef]
- Xu-Monette, Z.Y.; Zhou, J.; Young, K.H. PD-1 Expression and Clinical PD-1 Blockade in B-Cell Lymphomas. Blood 2018, 131, 68–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachonikola, E.; Stamatopoulos, K.; Chatzidimitriou, A. T Cells in Chronic Lymphocytic Leukemia: A Two-Edged Sword. Front. Immunol. 2020, 11, 612244. [Google Scholar] [CrossRef] [PubMed]
- Shiromizu, C.M.; Jancic, C.C. Γδ T Lymphocytes: An Effector Cell in Autoimmunity and Infection. Front. Immunol. 2018, 9, 2389. [Google Scholar] [CrossRef]
- Kabelitz, D.; Wesch, D.; He, W. Perspectives of Γδ T Cells in Tumor Immunology. Cancer Res. 2007, 67, 5–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigau, M.; Ostrouska, S.; Fulford, T.S.; Johnson, D.N.; Woods, K.; Ruan, Z.; McWilliam, H.E.G.; Hudson, C.; Tutuka, C.; Wheatley, A.K.; et al. Butyrophilin 2A1 Is Essential for Phosphoantigen Reactivity by Γδ T Cells. Science 2020, 367, eaay5516. [Google Scholar] [CrossRef] [PubMed]
- Harly, C.; Guillaume, Y.; Nedellec, S.; Peigné, C.-M.; Mönkkönen, H.; Mönkkönen, J.; Li, J.; Kuball, J.; Adams, E.J.; Netzer, S.; et al. Key Implication of CD277/Butyrophilin-3 (BTN3A) in Cellular Stress Sensing by a Major Human Γδ T-Cell Subset. Blood 2012, 120, 2269–2279. [Google Scholar] [CrossRef] [Green Version]
- Kenna, T.; Golden-Mason, L.; Norris, S.; Hegarty, J.E.; O’Farrelly, C.; Doherty, D.G. Distinct Subpopulations of Gamma Delta T Cells Are Present in Normal and Tumor-Bearing Human Liver. Clin. Immunol. 2004, 113, 56–63. [Google Scholar] [CrossRef]
- Mangan, B.A.; Dunne, M.R.; O’Reilly, V.P.; Dunne, P.J.; Exley, M.A.; O’Shea, D.; Scotet, E.; Hogan, A.E.; Doherty, D.G. Cutting Edge: CD1d Restriction and Th1/Th2/Th17 Cytokine Secretion by Human Vδ3 T Cells. J. Immunol. 2013, 191, 30–34. [Google Scholar] [CrossRef]
- Robak, E.; Błoński, J.Z.; Bartkowiak, J.; Niewiadomska, H.; Sysa-Jȩdrzejowska, A.; Robak, T. Circulating TCR Γδ Cells in the Patients with Systemic Lupus Erythematosus. Mediators Inflamm. 1999, 8, 305–312. [Google Scholar] [CrossRef]
- Kabelitz, D.; Hinz, T.; Dobmeyer, T.; Mentzel, U.; Marx, S.; Böhme, A.; Arden, B.; Rosssio, R.; Hoelzer, D. Clonal Expansion of Vγ3/Vδ3-Expressing Γδ T Cells in a HIV-1/2-Negative Patient with CD4 T-Cell Deficiency. Br. J. Haematol. 1997, 96, 266–271. [Google Scholar] [CrossRef]
- Paul, S.; Shilpi, n.; Lal, G. Role of Gamma-Delta (Γδ) T Cells in Autoimmunity. J. Leukoc. Biol. 2015, 97, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Zarobkiewicz, M.K.; Wawryk-Gawda, E.; Kowalska, W.; Janiszewska, M.; Bojarska-Junak, A. Γδ T Lymphocytes in Asthma: A Complicated Picture. Arch. Immunol. Ther. Exp. 2021, 69, 4. [Google Scholar] [CrossRef]
- Caron, J.; Ridgley, L.A.; Bodman-Smith, M. How to Train Your Dragon: Harnessing Gamma Delta T Cells Antiviral Functions and Trained Immunity in a Pandemic Era. Front. Immunol. 2021, 12, 983. [Google Scholar] [CrossRef] [PubMed]
- Agerholm, R.; Bekiaris, V. Evolved to Protect, Designed to Destroy: IL-17-Producing Γδ T Cells in Infection, Inflammation, and Cancer. Eur. J. Immunol. 2021, 51, 2164–2177. [Google Scholar] [CrossRef] [PubMed]
- Pizzolato, G.; Kaminski, H.; Tosolini, M.; Franchini, D.-M.; Pont, F.; Martins, F.; Valle, C.; Labourdette, D.; Cadot, S.; Quillet-Mary, A.; et al. Single-Cell RNA Sequencing Unveils the Shared and the Distinct Cytotoxic Hallmarks of Human TCRVδ1 and TCRVδ2 Γδ T Lymphocytes. Proc. Natl. Acad. Sci. USA 2019, 116, 11906–11915. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, M.; Shimizu, T.; Ashihara, E.; Ukimura, O. Strategies to Improve the Antitumor Effect of Γδ T Cell Immunotherapy for Clinical Application. Int. J. Mol. Sci. 2021, 22, 8910. [Google Scholar] [CrossRef]
- Kabelitz, D.; Serrano, R.; Kouakanou, L.; Peters, C.; Kalyan, S. Cancer Immunotherapy with Γδ T Cells: Many Paths Ahead of Us. Cell. Mol. Immunol. 2020, 17, 925–939. [Google Scholar] [CrossRef]
- Barros, M.D.S.; de Araújo, N.D.; Magalhães-Gama, F.; Pereira Ribeiro, T.L.; Alves Hanna, F.S.; Tarragô, A.M.; Malheiro, A.; Costa, A.G. Γδ T Cells for Leukemia Immunotherapy: New and Expanding Trends. Front. Immunol. 2021, 12, 3770. [Google Scholar] [CrossRef]
- Correia, D.V.; Lopes, A.C.; Silva-Santos, B. Tumor Cell Recognition by Γδ T Lymphocytes. OncoImmunology 2013, 2, e22892. [Google Scholar] [CrossRef] [Green Version]
- Barrow, A.D.; Martin, C.J.; Colonna, M. The Natural Cytotoxicity Receptors in Health and Disease. Front. Immunol. 2019, 10, 909. [Google Scholar] [CrossRef] [Green Version]
- Parodi, M.; Favoreel, H.; Candiano, G.; Gaggero, S.; Sivori, S.; Mingari, M.C.; Moretta, L.; Vitale, M.; Cantoni, C. NKp44-NKp44 Ligand Interactions in the Regulation of Natural Killer Cells and Other Innate Lymphoid Cells in Humans. Front. Immunol. 2019, 10, 719. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; Shemesh, A.; Brusilovsky, M.; Porgador, A.; Campbell, K.S. Regulation of the Functions of Natural Cytotoxicity Receptors by Interactions with Diverse Ligands and Alterations in Splice Variant Expression. Front. Immunol. 2017, 8, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zingoni, A.; Ardolino, M.; Santoni, A.; Cerboni, C. NKG2D and DNAM-1 Activating Receptors and Their Ligands in NK-T Cell Interactions: Role in the NK Cell-Mediated Negative Regulation of T Cell Responses. Front. Immunol. 2013, 3, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, J.-L.; Benyamine, A.; Pasero, C.; Olive, D. New Insights into the Regulation of Γδ T Cells by BTN3A and Other BTN/BTNL in Tumor Immunity. Front. Immunol. 2018, 9, 1601. [Google Scholar] [CrossRef] [Green Version]
- Uldrich, A.P.; Rigau, M.; Godfrey, D.I. Immune Recognition of Phosphoantigen-Butyrophilin Molecular Complexes by Γδ T Cells. Immunol. Rev. 2020, 298, 74–83. [Google Scholar] [CrossRef]
- Tokuyama, H.; Hagi, T.; Mattarollo, S.R.; Morley, J.; Wang, Q.; Fai-So, H.; Moriyasu, F.; Nieda, M.; Nicol, A.J. Vγ9Vδ2 T Cell Cytotoxicity against Tumor Cells Is Enhanced by Monoclonal Antibody Drugs—Rituximab and Trastuzumab. Int. J. Cancer 2008, 122, 2526–2534. [Google Scholar] [CrossRef]
- Chen, Z.; Freedman, M.S. CD16+ Γδ T Cells Mediate Antibody Dependent Cellular Cytotoxicity: Potential Mechanism in the Pathogenesis of Multiple Sclerosis. Clin. Immunol. 2008, 128, 219–227. [Google Scholar] [CrossRef]
- Chang, K.-M.; Traum, D.; Park, J.-J.; Ho, S.; Ojiro, K.; Wong, D.K.; Wahed, A.S.; Terrault, N.A.; Khalili, M.; Sterling, R.K.; et al. Distinct Phenotype and Function of Circulating Vδ1+ and Vδ2+ ΓδT-Cells in Acute and Chronic Hepatitis B. PLoS Pathog. 2019, 15, e1007715. [Google Scholar] [CrossRef]
- Deseke, M.; Prinz, I. Ligand Recognition by the Γδ TCR and Discrimination between Homeostasis and Stress Conditions. Cell. Mol. Immunol. 2020, 17, 914–924. [Google Scholar] [CrossRef]
- Vermijlen, D.; Gatti, D.; Kouzeli, A.; Rus, T.; Eberl, M. Γδ T Cell Responses: How Many Ligands Will It Take till We Know? Semin. Cell Dev. Biol. 2018, 84, 75–86. [Google Scholar] [CrossRef]
- de Sousa, T.R.; Victor, J.R. Natural Self-Ligand Gamma Delta T Cell Receptors (ΓδTCRs) Insight: The Potential of Induced IgG. Vaccines 2020, 8, 436. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, H.K. Function of Γδ T Cells in Tumor Immunology and Their Application to Cancer Therapy. Exp. Mol. Med. 2021, 53, 318–327. [Google Scholar] [CrossRef]
- Rossi, C.; Gravelle, P.; Decaup, E.; Bordenave, J.; Poupot, M.; Tosolini, M.; Franchini, D.-M.; Laurent, C.; Morin, R.; Lagarde, J.-M.; et al. Boosting Γδ T Cell-Mediated Antibody-Dependent Cellular Cytotoxicity by PD-1 Blockade in Follicular Lymphoma. OncoImmunology 2019, 8, 1554175. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, M.; Tanaka, Y.; Kobayashi, H.; Murata-Hirai, K.; Miyabe, H.; Sugie, T.; Toi, M.; Minato, N. Expression and Function of PD-1 in Human Γδ T Cells That Recognize Phosphoantigens. Eur. J. Immunol. 2011, 41, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.J.; Lee, J.J.; Kang, S.H.; Suh, J.K.; Choi, E.S.; Jang, S.; Hwang, S.-H.; Koh, K.-N.; Im, H.J.; Kim, N. The BTLA and PD-1 Signaling Pathways Independently Regulate the Proliferation and Cytotoxicity of Human Peripheral Blood Γδ T Cells. Immun. Inflamm. Dis. 2021, 9, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Gertner-Dardenne, J.; Fauriat, C.; Orlanducci, F.; Thibult, M.-L.; Pastor, S.; Fitzgibbon, J.; Bouabdallah, R.; Xerri, L.; Olive, D. The Co-Receptor BTLA Negatively Regulates Human Vγ9Vδ2 T-Cell Proliferation: A Potential Way of Immune Escape for Lymphoma Cells. Blood 2013, 122, 922–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogoi, D.; Biswas, D.; Borkakoty, B.; Mahanta, J. Exposure to Plasmodium Vivax Is Associated with the Increased Expression of Exhaustion Markers on Γδ T Lymphocytes. Parasite Immunol. 2018, 40, e12594. [Google Scholar] [CrossRef]
- Jin, Z.; Lan, T.; Zhao, Y.; Du, J.; Chen, J.; Lai, J.; Xu, L.; Chen, S.; Zhong, X.; Wu, X.; et al. Higher TIGIT+CD226- Γδ T Cells in Patients with Acute Myeloid Leukemia. Immunol. Investig. 2020, 51, 1–11. [Google Scholar] [CrossRef]
- Li, X.; Lu, H.; Gu, Y.; Zhang, X.; Zhang, G.; Shi, T.; Chen, W. Tim-3 Suppresses the Killing Effect of Vγ9Vδ2 T Cells on Colon Cancer Cells by Reducing Perforin and Granzyme B Expression. Exp. Cell Res. 2020, 386, 111719. [Google Scholar] [CrossRef]
- Wu, K.; Feng, J.; Xiu, Y.; Li, Z.; Lin, Z.; Zhao, H.; Zeng, H.; Xia, W.; Yu, L.; Xu, B. Vδ2 T Cell Subsets, Defined by PD-1 and TIM-3 Expression, Present Varied Cytokine Responses in Acute Myeloid Leukemia Patients. Int. Immunopharmacol. 2020, 80, 106122. [Google Scholar] [CrossRef]
- Halary, F.; Peyrat, M.-A.; Champagne, E.; Lopez-Botet, M.; Moretta, A.; Moretta, L.; Vié, H.; Fournié, J.-J.; Bonneville, M. Control of Self-Reactive Cytotoxic T Lymphocytes Expressing Γδ T Cell Receptors by Natural Killer Inhibitory Receptors. Eur. J. Immunol. 1997, 27, 2812–2821. [Google Scholar] [CrossRef] [PubMed]
- Raverdeau, M.; Cunningham, S.P.; Harmon, C.; Lynch, L. Γδ T Cells in Cancer: A Small Population of Lymphocytes with Big Implications. Clin. Transl. Immunol. 2019, 8, e01080. [Google Scholar] [CrossRef] [PubMed]
- Bartkowiak, J.; Błoński, J.Z.; Niewiadomska, H.; Kulczycka, D.; Robak, T. Characterization of Γδ T Cells in Peripheral Blood from Patients with B-Cell Chronic Lymphocytic Leukaemia. Biomed. Lett. 1998, 58, 19–30. [Google Scholar]
- Bartkowiak, J.; Kulczycka-Wojdala, D.; Blonski, J.Z.; Robak, T. Molecular Diversity of Γδ T Cells in Peripheral Blood from Patients with B-Cell Chronic Lymphocytic Leukaemia. Neoplasma 2002, 49, 86–90. [Google Scholar]
- Poggi, A.; Venturino, C.; Catellani, S.; Clavio, M.; Miglino, M.; Gobbi, M.; Steinle, A.; Ghia, P.; Stella, S.; Caligaris-Cappio, F.; et al. Vδ1 T Lymphocytes from B-CLL Patients Recognize ULBP3 Expressed on Leukemic B Cells and up-Regulated by Trans-Retinoic Acid. Cancer Res. 2004, 64, 9172–9179. [Google Scholar] [CrossRef] [Green Version]
- Własiuk, P.; Giannopoulos, K.; Kowal, M.; Wa̧sik-Szczepanek, E.; Bojarska-Junak, A.; Chocholska, S.; Roliński, J.; Dmoszyńska, A. The Frequency of Lymphocytes Expressing Γδ T-Cell Receptor Is Decreased after Thalidomide and Fludarabine Therapy in Patients with B-Cell Lymphocytic Leukemia. Acta Haematol. Pol. 2007, 38, 133–140. [Google Scholar]
- Simões, C.; Silva, I.; Carvalho, A.; Silva, S.; Santos, S.; Marques, G.; Ribeiro, A.; Roque, A.; Carda, J.; Sarmento-Ribeiro, A.B.; et al. Quantification and Phenotypic Characterization of Peripheral Blood Vδ1 + T Cells in Chronic Lymphocytic Leukemia and Monoclonal B Cell Lymphocytosis. Cytom. Part B—Clin. Cytom. 2019, 96, 164–168. [Google Scholar] [CrossRef]
- Chabab, G.; Barjon, C.; Abdellaoui, N.; Salvador-Prince, L.; Dejou, C.; Michaud, H.; Boissière-Michot, F.; Lopez-Crapez, E.; Jacot, W.; Pourquier, D.; et al. Identification of a Regulatory Vδ1 Gamma Delta T Cell Subpopulation Expressing CD73 in Human Breast Cancer. J. Leukoc. Biol. 2020, 107, 1057–1067. [Google Scholar] [CrossRef]
- Rong, L.; Li, K.; Li, R.; Liu, H.-M.; Sun, R.; Liu, X.-Y. Analysis of Tumor-Infiltrating Gamma Delta T Cells in Rectal Cancer. World J. Gastroenterol. 2016, 22, 3573–3580. [Google Scholar] [CrossRef]
- Morrow, E.S.; Roseweir, A.; Edwards, J. The Role of Gamma Delta T Lymphocytes in Breast Cancer: A Review. Transl. Res. 2019, 203, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Peters, C.; Kabelitz, D.; Wesch, D. Regulatory Functions of Γδ T Cells. Cell. Mol. Life Sci. CMLS 2018, 75, 2125–2135. [Google Scholar] [CrossRef]
- Andreu-Ballester, J.C.; García-Ballesteros, C.; Benet-Campos, C.; Amigó, V.; Almela-Quilis, A.; Mayans, J.; Ballester, F. Values for Aβ and Γδ T-Lymphocytes and CD4+, CD8+, and CD56+ Subsets in Healthy Adult Subjects: Assessment by Age and Gender. Cytometry B Clin. Cytom. 2012, 82B, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Gaballa, A.; Arruda, L.C.M.; Rådestad, E.; Uhlin, M. CD8+γδ T Cells Are More Frequent in CMV Seropositive Bone Marrow Grafts and Display Phenotype of an Adaptive Immune Response. Stem Cells Int. 2019, 2019, e6348060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherardin, N.A.; Waldeck, K.; Caneborg, A.; Martelotto, L.G.; Balachander, S.; Zethoven, M.; Petrone, P.M.; Pattison, A.; Wilmott, J.S.; Quiñones-Parra, S.M.; et al. Γδ T Cells in Merkel Cell Carcinomas Have a Proinflammatory Profile Prognostic of Patient Survival. Cancer Immunol. Res. 2021, 9, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Girard, P.; Charles, J.; Cluzel, C.; Degeorges, E.; Manches, O.; Plumas, J.; De Fraipont, F.; Leccia, M.-T.; Mouret, S.; Chaperot, L.; et al. The Features of Circulating and Tumor-Infiltrating Γδ T Cells in Melanoma Patients Display Critical Perturbations with Prognostic Impact on Clinical Outcome. OncoImmunology 2019, 8, 1601483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odaira, K.; Kimura, S.-N.; Fujieda, N.; Kobayashi, Y.; Kambara, K.; Takahashi, T.; Izumi, T.; Matsushita, H.; Kakimi, K. CD27(−)CD45(+) Γδ T Cells Can Be Divided into Two Populations, CD27(-)CD45(Int) and CD27(-)CD45(Hi) with Little Proliferation Potential. Biochem. Biophys. Res. Commun. 2016, 478, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- De Weerdt, I.; Hofland, T.; Lameris, R.; Endstra, S.; Jongejan, A.; Moerland, P.D.; De Bruin, R.C.G.; Remmerswaal, E.B.M.; Ten Berge, I.J.M.; Liu, N.; et al. Improving CLL Vγ9Vδ2-T-Cell Fitness for Cellular Therapy by Ex Vivo Activation and Ibrutinib. Blood 2018, 132, 2260–2272. [Google Scholar] [CrossRef] [Green Version]
- Siegers, G.M.; Lamb, L.S. Cytotoxic and Regulatory Properties of Circulating Vδ1+ Γδ T Cells: A New Player on the Cell Therapy Field? Mol. Ther. 2014, 22, 1416–1422. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Wu, P.; Wu, X.; Ye, J.; Wang, Z.; Zhao, S.; Ni, C.; Hu, G.; Xu, J.; Han, Y.; et al. Ex Vivo Expanded Human Circulating Vδ1 ΓδT Cells Exhibit Favorable Therapeutic Potential for Colon Cancer. OncoImmunology 2015, 4, e992749. [Google Scholar] [CrossRef] [Green Version]
- Knight, A.; MacKinnon, S.; Lowdell, M.W. Human Vdelta1 Gamma-Delta T Cells Exert Potent Specific Cytotoxicity against Primary Multiple Myeloma Cells. Cytotherapy 2012, 14, 1110–1118. [Google Scholar] [CrossRef]
- Correia, D.V.; Fogli, M.; Hudspeth, K.; Gomes Da Silva, M.; Mavilio, D.; Silva-Santos, B. Differentiation of Human Peripheral Blood Vδ1+ T Cells Expressing the Natural Cytotoxicity Receptor NKp30 for Recognition of Lymphoid Leukemia Cells. Blood 2011, 118, 992–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, A.R.; Correia, D.V.; Fernandes-Platzgummer, A.; Da Silva, C.L.; Da Silva, M.G.; Anjos, D.R.; Silva-Santos, B. Delta One T Cells for Immunotherapy of Chronic Lymphocytic Leukemia: Clinical-Grade Expansion/Differentiation and Preclinical Proof of Concept. Clin. Cancer Res. 2016, 22, 5795–5804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polito, V.A.; Cristantielli, R.; Weber, G.; Del Bufalo, F.; Belardinilli, T.; Arnone, C.M.; Petretto, A.; Antonucci, L.; Giorda, E.; Tumino, N.; et al. Universal Ready-to-Use Immunotherapeutic Approach for the Treatment of Cancer: Expanded and Activated Polyclonal Γδ Memory T Cells. Front. Immunol. 2019, 10, 2717. [Google Scholar] [CrossRef] [Green Version]
- Simoes, A.; Kawalkowska, J.; Recaldin, T.; Foxler, D.; Nussbaumer, O.; Koslowski, M. Vδ1+ T Cells: Adoptive Cell Therapy for the Treatment of Haematological Malignancies in Allogeneic Settings. Blood 2019, 134, 3221. [Google Scholar] [CrossRef]
- Siegers, G.M.; Dhamko, H.; Wang, X.-H.; Mathieson, A.M.; Kosaka, Y.; Felizardo, T.C.; Medin, J.A.; Tohda, S.; Schueler, J.; Fisch, P.; et al. Human Vδ1 Γδ T Cells Expanded from Peripheral Blood Exhibit Specific Cytotoxicity against B-Cell Chronic Lymphocytic Leukemia-Derived Cells. Cytotherapy 2011, 13, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Coscia, M.; Vitale, C.; Peola, S.; Foglietta, M.; Rigoni, M.; Griggio, V.; Castella, B.; Angelini, D.; Chiaretti, S.; Riganti, C.; et al. Dysfunctional Vγ9Vδ2 T Cells Are Negative Prognosticators and Markers of Dysregulated Mevalonate Pathway Activity in Chronic Lymphocytic Leukemia Cells. Blood 2012, 120, 3271–3279. [Google Scholar] [CrossRef] [Green Version]
- Bruni, E.; Cazzetta, V.; Donadon, M.; Cimino, M.; Torzilli, G.; Spata, G.; Leonardi, G.; Dieli, F.; Mikulak, J.; Mavilio, D. Chemotherapy Accelerates Immune-Senescence and Functional Impairments of Vδ2pos T Cells in Elderly Patients Affected by Liver Metastatic Colorectal Cancer. J. Immunother. Cancer 2019, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Farrington, L.A.; Jagannathan, P.; McIntyre, T.I.; Vance, H.M.; Bowen, K.; Boyle, M.J.; Nankya, F.; Wamala, S.; Auma, A.; Nalubega, M.; et al. Frequent Malaria Drives Progressive Vδ2 T-Cell Loss, Dysfunction, and CD16 Up-Regulation During Early Childhood. J. Infect. Dis. 2016, 213, 1483–1490. [Google Scholar] [CrossRef]
- Ghosh, A.; Mondal, R.K.; Romani, S.; Bagchi, S.; Cairo, C.; Pauza, C.D.; Kottilil, S.; Poonia, B. Persistent Gamma Delta T-Cell Dysfunction in Chronic HCV Infection despite Direct-Acting Antiviral Therapy Induced Cure. J. Viral Hepat. 2019, 26, 1105–1116. [Google Scholar] [CrossRef]
- Li, H.; Chaudry, S.; Poonia, B.; Shao, Y.; Pauza, C.D. Depletion and Dysfunction of Vγ2Vδ2 T Cells in HIV Disease: Mechanisms, Impacts and Therapeutic Implications. Cell. Mol. Immunol. 2013, 10, 42–49. [Google Scholar] [CrossRef]
- Argentati, K.; Re, F.; Donnini, A.; Tucci, M.G.; Franceschi, C.; Bartozzi, B.; Bernardini, G.; Provinciali, M. Numerical and Functional Alterations of Circulating Γδ T Lymphocytes in Aged People and Centenarians. J. Leukoc. Biol. 2002, 72, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Colonna-Romano, G.; Aquino, A.; Bulati, M.; Lio, D.; Candore, G.; Oddo, G.; Scialabba, G.; Vitello, S.; Caruso, C. Impairment of Gamma/Delta T Lymphocytes in Elderly: Implications for Immunosenescence. Exp. Gerontol. 2004, 39, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Colonna-Romano, G.; Potestio, M.; Aquino, A.; Candore, G.; Lio, D.; Caruso, C. Gamma/Delta T Lymphocytes Are Affected in the Elderly. Exp. Gerontol. 2002, 37, 205–211. [Google Scholar] [CrossRef]
- Wistuba-Hamprecht, K.; Frasca, D.; Blomberg, B.; Pawelec, G.; Derhovanessian, E. Age-Associated Alterations in Γδ T-Cells Are Present Predominantly in Individuals Infected with Cytomegalovirus. Immun. Ageing 2013, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wistuba-Hamprecht, K.; Haehnel, K.; Janssen, N.; Demuth, I.; Pawelec, G. Peripheral Blood T-Cell Signatures from High-Resolution Immune Phenotyping of Γδ and Aβ T-Cells in Younger and Older Subjects in the Berlin Aging Study II. Immun. Ageing 2015, 12, 25. [Google Scholar] [CrossRef] [Green Version]
- Pourgheysari, B.; Bruton, R.; Parry, H.; Billingham, L.; Fegan, C.; Murray, J.; Moss, P. The Number of Cytomegalovirus-Specific CD4+ T Cells Is Markedly Expanded in Patients with B-Cell Chronic Lymphocytic Leukemia and Determines the Total CD4+ T-Cell Repertoire. Blood 2010, 116, 2968–2974. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.; Sakuta, K.; Noguchi, A.; Ariyoshi, N.; Sato, K.; Sato, S.; Sato, K.; Hosoi, A.; Nakajima, J.; Yoshida, Y.; et al. Zoledronate Facilitates Large-Scale Ex Vivo Expansion of Functional Γδ T Cells from Cancer Patients for Use in Adoptive Immunotherapy. Cytotherapy 2008, 10, 842–856. [Google Scholar] [CrossRef]
- Fournié, J.-J.; Sicard, H.; Poupot, M.; Bezombes, C.; Blanc, A.; Romagné, F.; Ysebaert, L.; Laurent, G. What Lessons Can Be Learned from Γδ T Cell-Based Cancer Immunotherapy Trials? Cell. Mol. Immunol. 2013, 10, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Singer, N.G.; Fox, D.A.; Haqqi, T.M.; Beretta, L.; Endres, J.S.; Prohaska, S.; Parnes, J.R.; Bromberg, J.; Sramkoski, R.M. CD6: Expression during Development, Apoptosis and Selection of Human and Mouse Thymocytes. Int. Immunol. 2002, 14, 585–597. [Google Scholar] [CrossRef] [Green Version]
- Rossini, M.; Adami, S.; Viapiana, O.; Fracassi, E.; Ortolani, R.; Vella, A.; Zanotti, R.; Tripi, G.; Idolazzi, L.; Gatti, D. Long-Term Effects of Amino-Bisphosphonates on Circulating Γδ T Cells. Calcif. Tissue Int. 2012, 91, 395–399. [Google Scholar] [CrossRef]
- Lo Presti, E.; Pizzolato, G.; Gulotta, E.; Cocorullo, G.; Gulotta, G.; Dieli, F.; Meraviglia, S. Current Advances in Γδ T Cell-Based Tumor Immunotherapy. Front. Immunol. 2017, 8, 1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Gokhale, S.; Jung, J.; Spirollari, E.; Tsai, J.; Arceo, J.; Wu, B.W.; Victor, E.; Xie, P. Multifaceted Immunomodulatory Effects of the BTK Inhibitors Ibrutinib and Acalabrutinib on Different Immune Cell Subsets—Beyond B Lymphocytes. Front. Cell Dev. Biol. 2021, 9, 2250. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Barr, P.M.; Robak, T.; Owen, C.; Ghia, P.; Tedeschi, A.; Bairey, O.; Hillmen, P.; Coutre, S.E.; Devereux, S.; et al. Long-Term Efficacy and Safety of First-Line Ibrutinib Treatment for Patients with CLL/SLL: 5 Years of Follow-up from the Phase 3 RESONATE-2 Study. Leukemia 2020, 34, 787–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, M.; Mulder, T.A.; Österborg, A. BTK Inhibitors in Chronic Lymphocytic Leukemia: Biological Activity and Immune Effects. Front. Immunol. 2021, 12, 2617. [Google Scholar] [CrossRef]
- Xia, S.; Liu, X.; Cao, X.; Xu, S. T-Cell Expression of Bruton’s Tyrosine Kinase Promotes Autoreactive T-Cell Activation and Exacerbates Aplastic Anemia. Cell. Mol. Immunol. 2020, 17, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Risnik, D.; Elías, E.E.; Keitelman, I.; Colado, A.; Podaza, E.; Cordini, G.; Vergara Rubio, M.; Fernández Grecco, H.; Bezares, R.F.; Borge, M.; et al. The Effect of Ibrutinib on Neutrophil and Γδ T Cell Functions. Leuk. Lymphoma 2020, 61, 2409–2418. [Google Scholar] [CrossRef]
- Colado, A.; Genoula, M.; Cougoule, C.; Marín Franco, J.L.; Almejún, M.B.; Risnik, D.; Kviatcovsky, D.; Podaza, E.; Elías, E.E.; Fuentes, F.; et al. Effect of the BTK Inhibitor Ibrutinib on Macrophage- and Γδ T Cell-Mediated Response against Mycobacterium Tuberculosis. Blood Cancer J. 2018, 8, 1–6. [Google Scholar] [CrossRef]
- Ran, F.; Liu, Y.; Wang, C.; Xu, Z.; Zhang, Y.; Liu, Y.; Zhao, G.; Ling, Y. Review of the Development of BTK Inhibitors in Overcoming the Clinical Limitations of Ibrutinib. Eur. J. Med. Chem. 2021, 229, 114009. [Google Scholar] [CrossRef]
- Xu, Y.; Xiang, Z.; Alnaggar, M.; Kouakanou, L.; Li, J.; He, J.; Yang, J.; Hu, Y.; Chen, Y.; Lin, L.; et al. Allogeneic Vγ9Vδ2 T-Cell Immunotherapy Exhibits Promising Clinical Safety and Prolongs the Survival of Patients with Late-Stage Lung or Liver Cancer. Cell. Mol. Immunol. 2021, 18, 427–439. [Google Scholar] [CrossRef]
- Rozenbaum, M.; Meir, A.; Aharony, Y.; Itzhaki, O.; Schachter, J.; Bank, I.; Jacoby, E.; Besser, M.J. Gamma-Delta CAR-T Cells Show CAR-Directed and Independent Activity Against Leukemia. Front. Immunol. 2020, 11, 1347. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarobkiewicz, M.K.; Bojarska-Junak, A.A. The Mysterious Actor—γδ T Lymphocytes in Chronic Lymphocytic Leukaemia (CLL). Cells 2022, 11, 661. https://doi.org/10.3390/cells11040661
Zarobkiewicz MK, Bojarska-Junak AA. The Mysterious Actor—γδ T Lymphocytes in Chronic Lymphocytic Leukaemia (CLL). Cells. 2022; 11(4):661. https://doi.org/10.3390/cells11040661
Chicago/Turabian StyleZarobkiewicz, Michał K., and Agnieszka A. Bojarska-Junak. 2022. "The Mysterious Actor—γδ T Lymphocytes in Chronic Lymphocytic Leukaemia (CLL)" Cells 11, no. 4: 661. https://doi.org/10.3390/cells11040661