
The Molecular Signature of Human Testicular Peritubular Cells Revealed by Single-Cell Analysis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. HTPCs Culture
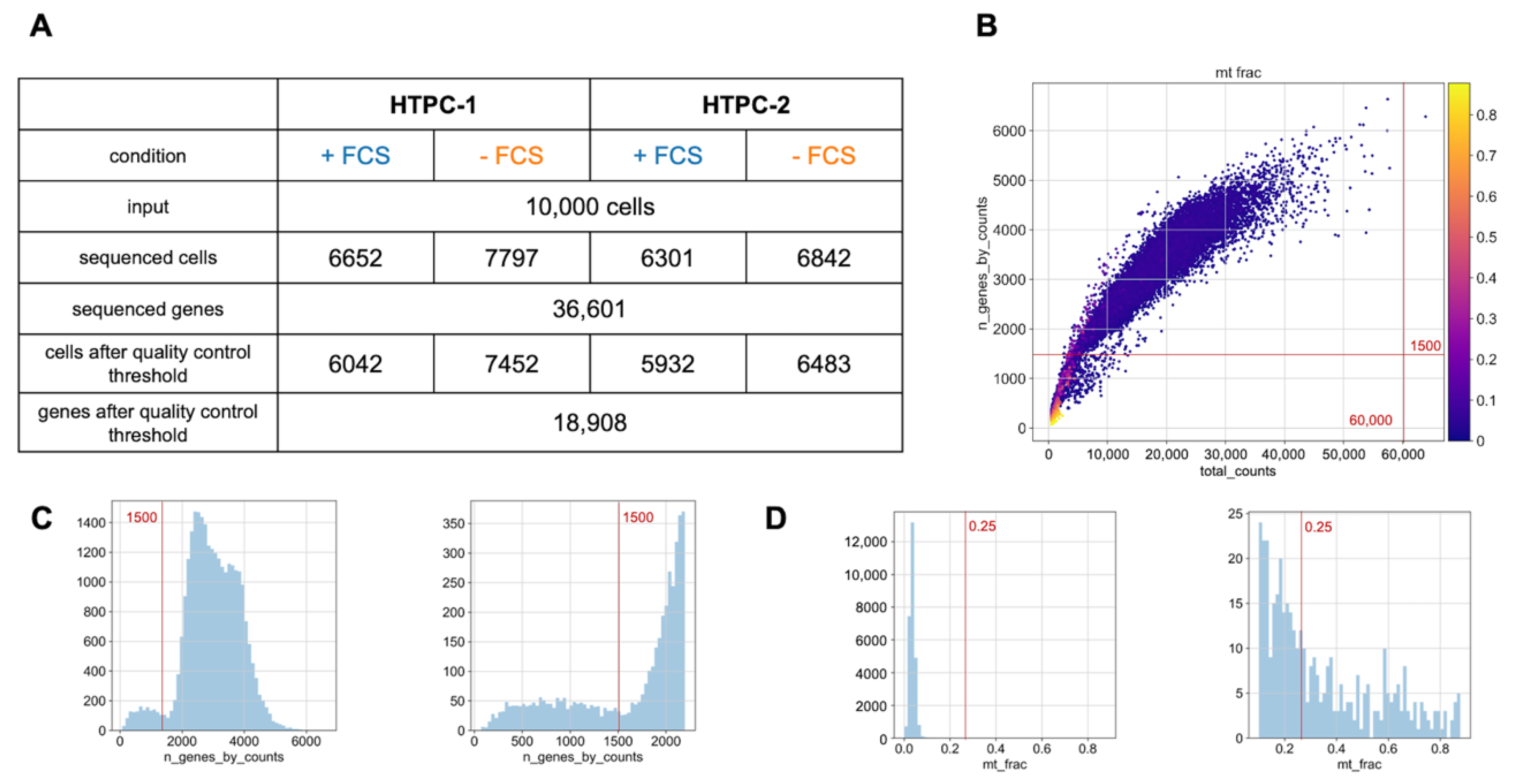
2.2. Single-Cell RNA Sequencing (scRNAseq)
2.3. Alignment and Data Analysis
2.4. RNA Velocity
2.5. Comparison with Existing Ex Vivo and In Situ Data
2.6. Comparison with Proteomic Data
2.7. Differentiation Studies
2.8. ELISA Test for Retinoic Acid
3. Results
3.1. General Remarks
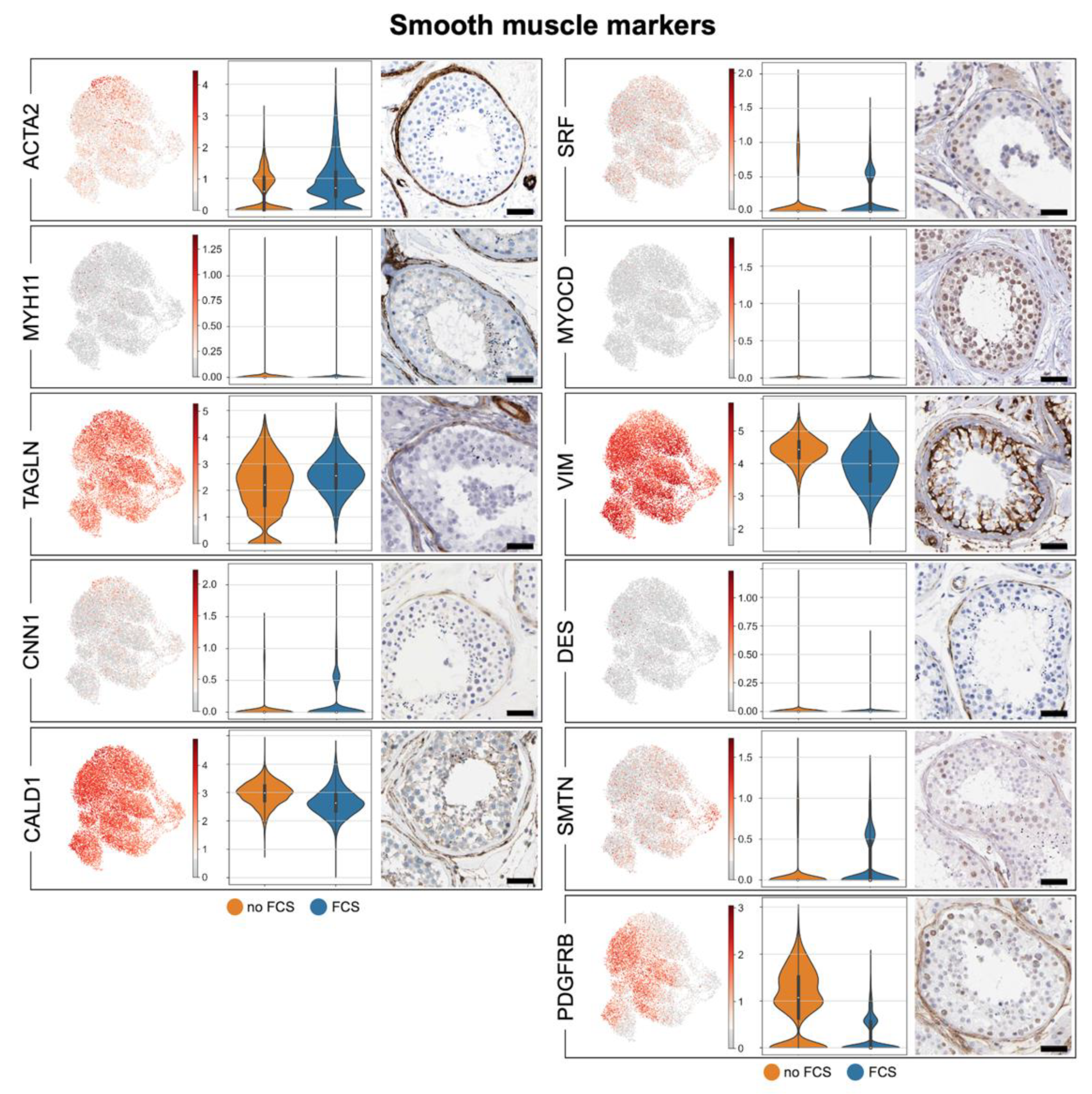
3.2. Human Testicular Peritubular Cells Are a Testicular Subtype of Smooth Muscle Cells
3.3. Human Testicular Peritubular Cells Are Producers of ECM, Which Is a Constituent of the Peritubular Wall
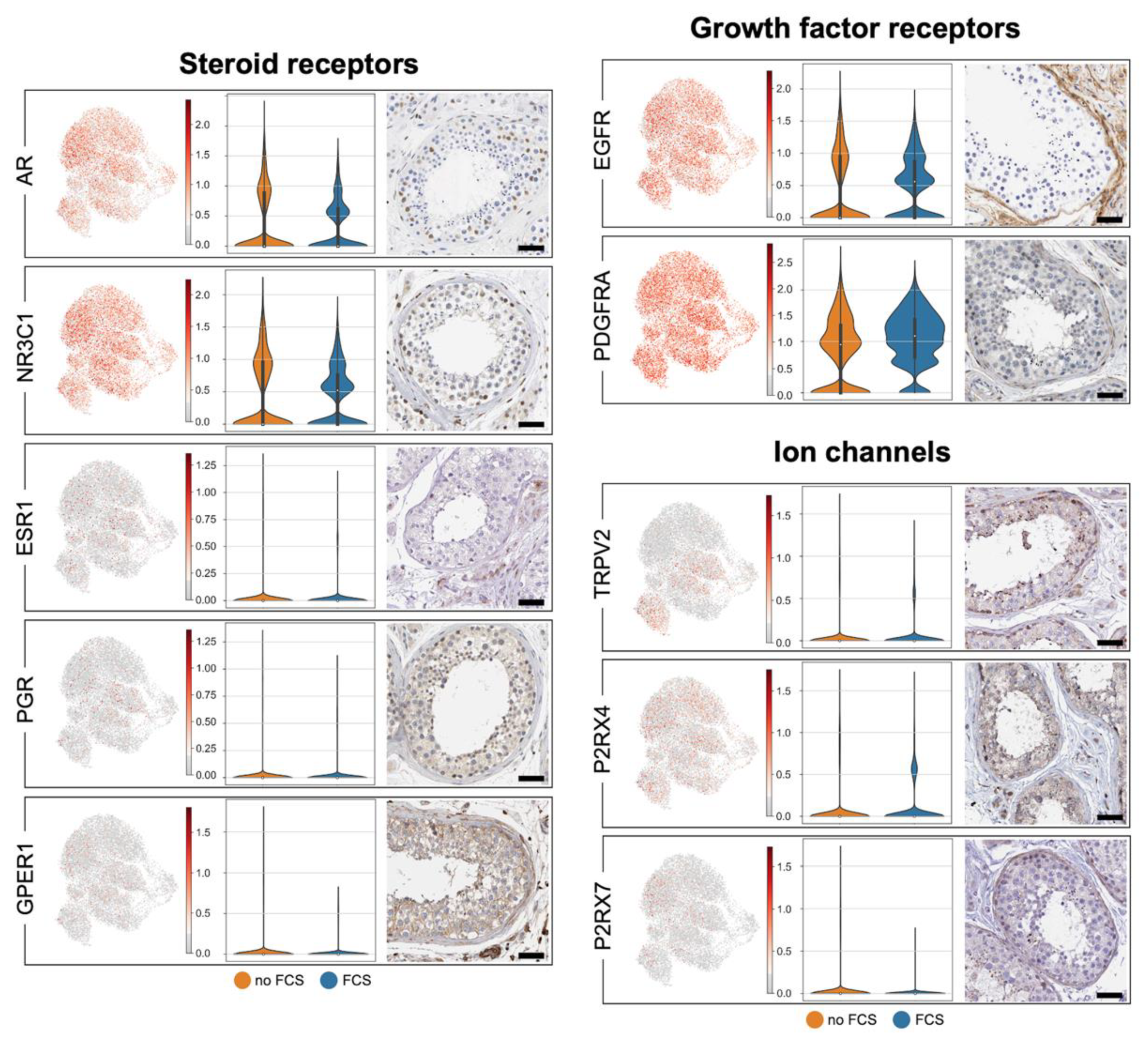
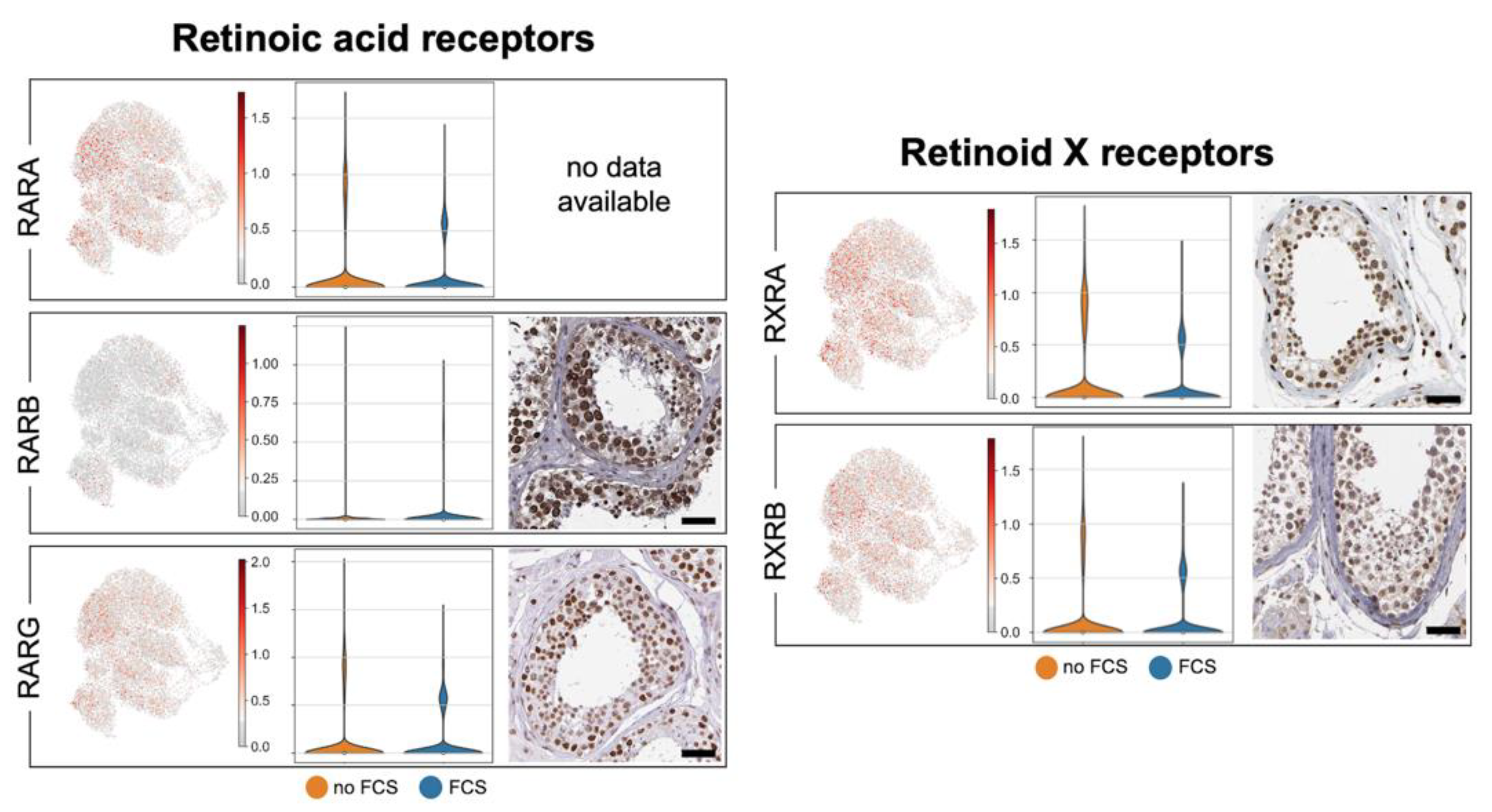
3.4. Human Testicular Peritubular Cells Are Targeted by Hormones and Local Factors: Receptors and Ion Channels
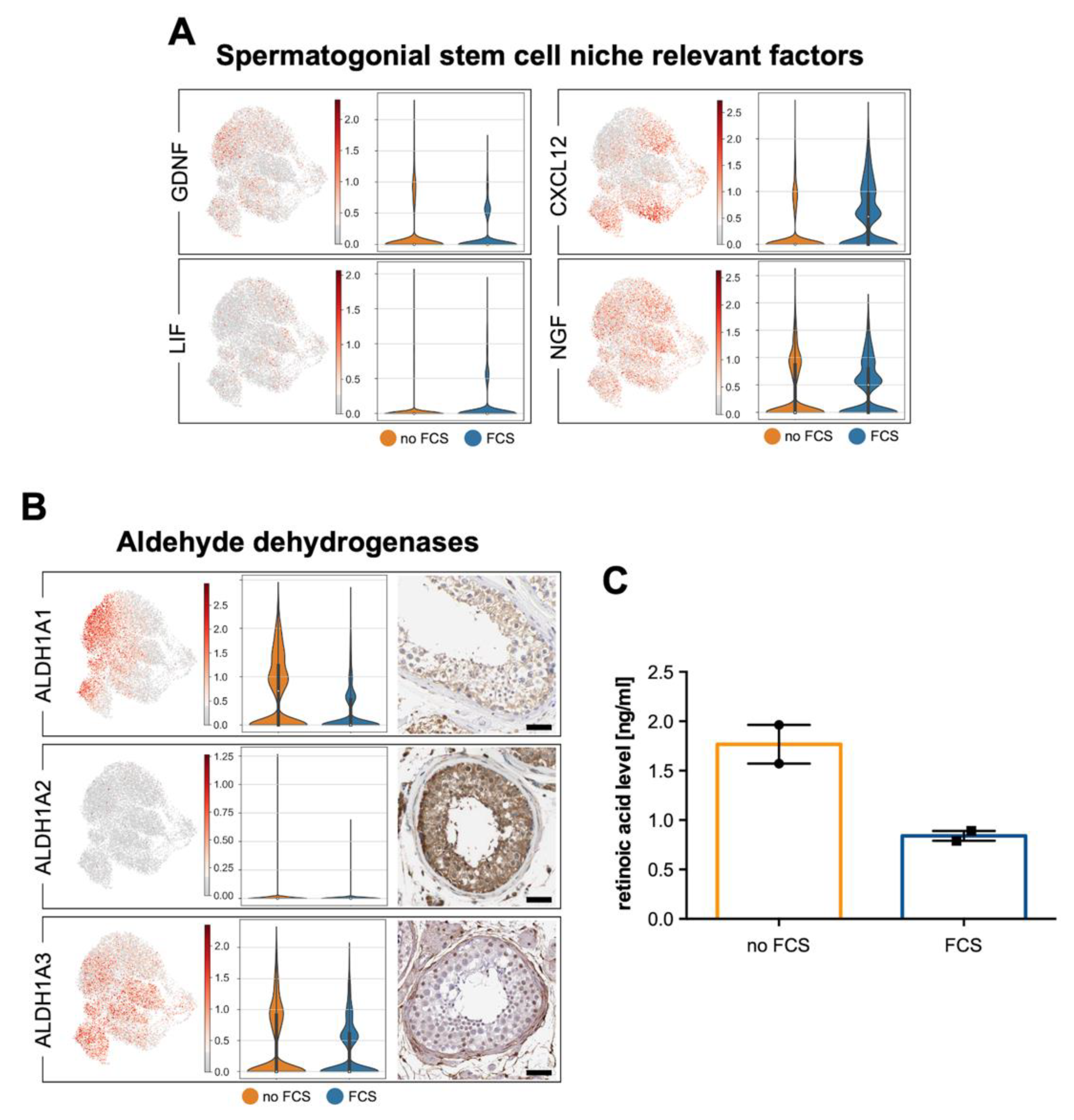
3.5. Human Testicular Peritubular Cells Are a Source of Factors Involved in the Regulation of Spermatogenesis
3.6. Human Testicular Peritubular Cells Express Mesenchymal Stromal Cell Markers and Leydig Stem Cell Markers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Link | Antibody |
|---|---|---|
| Smooth muscle markers | ||
| ACTA2 | https://images.proteinatlas.org/2/368_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB000002 |
| MYH11 | https://images.proteinatlas.org/15310/36933_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA015310 |
| TAGLN | https://images.proteinatlas.org/1447/4295_A_4_6.jpg (accessed on 15 October 2022) | Antibody CAB001447 |
| CNN1 | https://images.proteinatlas.org/7/1341_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB000007 |
| CALD1 | https://images.proteinatlas.org/8066/21805_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA008066 |
| SRF | https://images.proteinatlas.org/1819/7579_A_4_6.jpg (accessed on 15 October 2022) | Antibody HPA001819 |
| MYOCD | https://images.proteinatlas.org/46485/102910_A_5_6.jpg (accessed on 15 October 2022) | Antibody CAB046485 |
| VIM | https://images.proteinatlas.org/1762/7628_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA001762 |
| DES | https://images.proteinatlas.org/34/2106_A_4_6.jpg (accessed on 15 October 2022) | Antibody CAB000034 |
| SMTN | https://images.proteinatlas.org/51778/145653_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA051778 |
| PDGFRB | https://images.proteinatlas.org/3842/10254_A_4_6.jpg (accessed on 15 October 2022) | Antibody CAB003842 |
| Producers of ECM | ||
| DCN | https://images.proteinatlas.org/17118/38996_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB017118 |
| BGN | https://images.proteinatlas.org/3157/9472_A_4_6.jpg (accessed on 15 October 2022) | Antibody HPA003157 |
| COL1A1 | https://images.proteinatlas.org/11795/32976_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA011795 |
| Receptors | ||
| AR | https://images.proteinatlas.org/1/135_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB000001 |
| NR3C1 | https://images.proteinatlas.org/4248/13599_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA004248 |
| ESR1 | https://images.proteinatlas.org/55099/122099_A_4_6.jpg (accessed on 15 October 2022) | Antibody CAB055099 |
| PGR | https://images.proteinatlas.org/4751/15549_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA004751 |
| GPER1 | https://images.proteinatlas.org/27052/56949_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA027052 |
| Growth factor receptors | ||
| EGFR | https://images.proteinatlas.org/35/895_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB000035 |
| PDGFRA | https://images.proteinatlas.org/18143/43112_A_5_6.jpg (accessed on 15 October 2022) | Antibody CAB018143 |
| Ion channels | ||
| TRPV2 | https://images.proteinatlas.org/44993/102399_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA044993 |
| P2RX4 | https://images.proteinatlas.org/39494/85835_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA039494 |
| P2RX7 | https://images.proteinatlas.org/42013/103582_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA042013 |
| Aldehyde dehydrogenases | ||
| ALDH1A1 | https://images.proteinatlas.org/2123/8966_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA002123 |
| ALDH1A2 | https://images.proteinatlas.org/10022/24728_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA010022 |
| ALDH1A3 | https://images.proteinatlas.org/46271/105391_A_6_6.jpg (accessed on 15 October 2022) | Antibody HPA046271 |
| Mesenchymal stromal cell markers | ||
| ENG/CD105 | https://images.proteinatlas.org/67440/156475_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA067440 |
| CD73/NT5E | https://images.proteinatlas.org/17357/41086_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA017357 |
| CD90/THY1 | https://images.proteinatlas.org/68243/152840_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB068243 |
| CD34 | https://images.proteinatlas.org/36722/122320_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA036722 |
| CD14 | https://images.proteinatlas.org/72865/157660_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB072865 |
| CD79A | https://images.proteinatlas.org/19/247_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB000019 |
| CD19 | https://images.proteinatlas.org/16110/36120_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB016110 |
| Leydig cells factors | ||
| THY1 | https://images.proteinatlas.org/68243/152840_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB068243 |
| NES | https://images.proteinatlas.org/7007/20723_A_4_6.jpg (accessed on 15 October 2022) | Antibody HPA007007 |
| CD51/ITGAV | https://images.proteinatlas.org/2499/6125_A_4_6.jpg (accessed on 15 October 2022) | Antibody CAB002499 |
| PDGFRA | https://images.proteinatlas.org/18143/43112_A_5_6.jpg (accessed on 15 October 2022) | Antibody CAB018143 |
| TCF21 | https://images.proteinatlas.org/13189/60539_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA013189 |
| ENG/CD105 | https://images.proteinatlas.org/67440/156475_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA067440 |
| Steroidogenic markers | ||
| STAR | https://images.proteinatlas.org/27318/166039_A_4_6.jpg (accessed on 15 October 2022) | Antibody HPA027318 |
| CYP11A1 | https://images.proteinatlas.org/16436/142441_A_4_6.jpg (accessed on 15 October 2022) | Antibody HPA016436 |
| CYP19A1 | https://images.proteinatlas.org/355/2047_A_5_6.jpg (accessed on 15 October 2022) | Antibody CAB000355 |
| Retinoic acid & Retinoid X receptors | ||
| RARB | https://images.proteinatlas.org/4174/169826_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA004174 |
| RARG | https://images.proteinatlas.org/53883/121805_A_5_6.jpg (accessed on 15 October 2022) | Antibody HPA053883 |
| RXRA | https://images.proteinatlas.org/4565/13151_A_5_6.jpg (accessed on 15 October 2022) | Antibody CAB004565 |
| RXRB | https://images.proteinatlas.org/2003/5372_A_6_6.jpg (accessed on 15 October 2022) | Antibody CAB002003 |
| Gene Name | Nucleotide Sequence | Amplicon Size | Annealing Temperature |
|---|---|---|---|
| ALPL | 5′-GGA TGG GTG TCT CCA CAG TG-3′ 5′-GCT CTT CCA GGT GTC AAC GA-3′ | 593 bp | 60 °C |
| COMP | 5′-GGA TGC CTG TGA CAA CTG TC-3′ 5′-AAG GCC CTG AAG TCG GTG AG-3′ | 390 bp | 62 °C |
| INSL3 | 5′-GGG AGT CTC ACT CTG TTT CCC-3′ 5′-CAC TGT AGC AAC TCA CAT CGC-3′ | 108 bp | 60 °C |
| LHR | 5′-TGG AAA TGG ATT TGA AGA AGT ACA-3′ 5′-CAC GGA AGG CTC CAT TGT-3′ | 113 bp | 60 °C |
| PLIN1 | 5′-ACC TGC GAA TGC TTC CAG AA-3′ 5′-CAG GGG CTG ACT CTT CCT TG-3′ | 508 bp | 60 °C |
| STAR | 5′-ACG TGG ATT AAC CAG GTT CG-3′ 5′-CAG CCC TCT TGG TTG CTA AG-3′ | 149 bp | 58 °C |
| Gene Name | Description | HTPC-1 | HTPC-2 | TPCs of Nie et al. [17] | TPCs of Di Persio et al. [3] |
|---|---|---|---|---|---|
| ACTA2 | Smooth-Muscle Actin | √ | √ | √ | √ |
| ALDH1A1 | Aldehyde Dehydrogenase 1 Family Member A1 | √ | √ | √ | √ |
| ALDH1A2 | Aldehyde Dehydrogenase 1 Family Member A2 | (√) | ✕ | √ | (√) |
| ALDH1A3 | Aldehyde Dehydrogenase 1 Family Member A3 | √ | √ | √ | √ |
| AR | Androgen Receptor | √ | √ | √ | √ |
| ARX | Aristaless Related Homeobox | ✕ | (√) | √ | √ |
| BGN | Biglycan | √ | √ | √ | √ |
| CALD1 | Caldesmon 1 | √ | √ | √ | √ |
| CD14 | Monocyte Differentiation Antigen CD14 | (√) | ✕ | √ | ✕ |
| CD19 | B-Lymphocyte Surface Antigen B4 | ✕ | ✕ | ✕ | ✕ |
| CD34 | Hematopoietic Progenitor Cell Antigen CD34 | ✕ | ✕ | √ | √ |
| CD79A | B-Cell Antigen Receptor Complex-Associated Protein Alpha Chain | ✕ | ✕ | ✕ | ✕ |
| CNN1 | Calponin 1 | √ | √ | ✕ | √ |
| COL1A1 | Collagen Type I Alpha 1 Chain | √ | √ | √ | √ |
| CXCL12 | C-X-C Motif Chemokine Ligand 12 | √ | √ | √ | √ |
| CYP11A1 | Cholesterol Side-Chain Cleavage Enzyme | √ | √ | √ | √ |
| CYP19A1 | Aromatase | ✕ | ✕ | ✕ | ✕ |
| DCN | Decorin | √ | √ | √ | √ |
| DES | Desmin | √ | √ | √ | √ |
| EGFR | Epidermal Growth Factor Receptor | √ | √ | √ | √ |
| ENG | Endoglin | √ | √ | √ | √ |
| ESR1 | Estrogen Receptor | √ | √ | √ | (√) |
| GDNF | Glial Cell Derived Neurotrophic Factor | √ | (√) | ✕ | ✕ |
| GPER | G-Protein coupled Estrogen Receptor | √ | √ | ✕ | ✕ |
| ITGAV | Integrin Subunit Alpha V | √ | √ | √ | √ |
| LIF | Leukemia Inhibitory Factor | (√) | (√) | ✕ | ✕ |
| MYH11 | Myosin Heavy Chain 11 | √ | √ | √ | √ |
| MYOCD | Myocardin | (√) | ✕ | √ | √ |
| NES | NES | ✕ | ✕ | √ | (√) |
| NGF | Nerve Growth Factor | √ | √ | ✕ | ✕ |
| NR2F2 | NR2F2 | √ | √ | √ | √ |
| NR3C1 | Glucocorticoid Receptor | √ | √ | √ | √ |
| NT5E | Ecto-5’-Nucleotidase | √ | √ | √ | √ |
| P2RX4 | Purinergic Receptor P2X4 | √ | √ | √ | √ |
| P2RX7 | Purinergic Receptor P2X7 | √ | √ | √ | √ |
| PDGFRA | Platelet Derived Growth Factor Receptor Alpha | √ | √ | √ | √ |
| PDGFRB | Platelet Derived Growth Factor Receptor B | √ | √ | √ | √ |
| PGR | Progesterone Receptor | √ | √ | √ | √ |
| SMTN | Smoothelin | √ | √ | √ | √ |
| SRF | Serum Response Factor | √ | √ | √ | √ |
| STAR | Steroidogenic Acute Regulatory Protein | √ | √ | √ | √ |
| TAGLN | Transgelin | √ | √ | √ | √ |
| TCF21 | Transcription Factor 21 | √ | √ | √ | √ |
| THY1 | Thy-1 Cell Surface Antigen | √ | √ | √ | √ |
| TRPV2 | Transient Receptor Potential Cation Channel Subfamily V Member 2 | √ | √ | √ | √ |
| VIM | Vimentin | √ | √ | √ | √ |
| Leydig Cell Differentiation | ||||||
|---|---|---|---|---|---|---|
| Donor | qPCR analysis | |||||
| LHR | INSL3 | STAR | ||||
| 1 | Ctrl. | Diff. | Ctrl. | Diff. | Ctrl. | Diff. |
| 2 | n.d. | n.d. | n.d. | n.d. | + | ++ |
| 3 | n.d. | n.d. | n.d. | n.d. | + | ++ |
| 4 | n.d. | n.d. | n.d. | n.d. | + | ++ |
| Osteoblast Differentiation | ||||||
| Donor | qPCR analysis | Alc. ph. staining | ||||
| COMP | ALPL | |||||
| Ctrl. | Diff. | Ctrl. | Diff. | Ctrl. | Diff. | |
| 1 | + | +++ | + | + | n.d. | n.d. |
| 2 | + | +++ | + | + | n.d. | n.d. |
| Adipocyte Differentiation | ||||||
| Donor | qPCR analysis | Oil red staining | ||||
| PLIN1 | ||||||
| Ctrl. | Diff. | Ctrl. | Diff. | |||
| 1 | n.d. | n.d. | n.d. | n.d. | ||
| 2 | + | + | n.d. | n.d. | ||
| 3 | + | + | ||||
| 4 | + | + | ||||
References
- Li, L.; Papadopoulos, V. Advances in stem cell research for the treatment of primary hypogonadism. Nat. Rev. Urol. 2021, 18, 487–507. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tian, E.; Wang, Y.; Wen, Z.; Lei, Z.; Zhong, Y.; Ge, R.S. Stem Leydig cells: Current research and future prospects of regenerative medicine of male reproductive health. Semin. Cell Dev. Biol. 2022, 121, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Di Persio, S.; Tekath, T.; Siebert-Kuss, L.M.; Cremers, J.F.; Wistuba, J.; Li, X.; Meyer Zu Horste, G.; Drexler, H.C.A.; Wyrwoll, M.J.; Tüttelmann, F.; et al. Single-cell RNA-seq unravels alterations of the human spermatogonial stem cell compartment in patients with impaired spermatogenesis. Cell Rep. Med. 2021, 2, 100395. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Guo, J.; Sosa, E.; Chitiashvili, T.; Nie, X.; Rojas, E.J.; Oliver, E.; DonorConnect; Plath, K.; Hotaling, J.M.; Stukenborg, J.B.; et al. Single-cell analysis of the developing human testis reveals somatic niche cell specification and fetal germline stem cell establishment. Cell Stem Cell 2021, 28, 764–778.e4. [Google Scholar] [CrossRef]
- Sohni, A.; Tan, K.; Song, H.W.; Burow, D.; de Rooij, D.G.; Laurent, L.; Hsieh, T.C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517.e4. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, M.; Kamimura, K.; Nagano, T. Peritubular myoid cells in the testis: Their structure and function. Arch. Histol. Cytol. 1996, 59, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayerhofer, A. Human testicular peritubular cells: More than meets the eye. Reproduction 2013, 145, R107–R116. [Google Scholar] [CrossRef] [Green Version]
- Ross, M.H.; Long, I.R. Contractile cells in human seminiferous tubules. Science 1966, 153, 1271–1273. [Google Scholar] [CrossRef]
- Adam, M.; Schwarzer, J.U.; Köhn, F.M.; Strauss, L.; Poutanen, M.; Mayerhofer, A. Mast cell tryptase stimulates production of decorin by human testicular peritubular cells: Possible role of decorin in male infertility by interfering with growth factor signaling. Hum. Reprod. 2011, 26, 2613–2625. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Calandra, R.S.; Lustig, L.; Meineke, V.; Kohn, F.M.; Vogt, H.J.; Mayerhofer, A. Number, distribution pattern, and identification of macrophages in the testes of infertile men. Fertil. Steril. 2002, 78, 298–306. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Weidinger, S.; Meineke, V.; Kohn, F.M.; Mayerhofer, A. Proliferative action of mast-cell tryptase is mediated by PAR2, COX2, prostaglandins, and PPARgamma: Possible relevance to human fibrotic disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 15072–15077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauerhof, A.C.; Nicolas, N.; Bhushan, S.; Wahle, E.; Loveland, K.A.; Fietz, D.; Bergmann, M.; Groome, N.P.; Kliesch, S.; Schuppe, H.C.; et al. Investigation of activin A in inflammatory responses of the testis and its role in the development of testicular fibrosis. Hum. Reprod. 2019, 34, 1536–1550. [Google Scholar] [CrossRef]
- Schell, C.; Albrecht, M.; Spillner, S.; Mayer, C.; Kunz, L.; Köhn, F.M.; Schwarzer, U.; Mayerhofer, A. 15-Deoxy-delta 12-14-prostaglandin-J2 induces hypertrophy and loss of contractility in human testicular peritubular cells: Implications for human male fertility. Endocrinology 2010, 151, 1257–1268. [Google Scholar] [CrossRef] [Green Version]
- Welter, H.; Kampfer, C.; Lauf, S.; Feil, R.; Schwarzer, J.U.; Köhn, F.M.; Mayerhofer, A. Partial loss of contractile marker proteins in human testicular peritubular cells in infertility patients. Andrology 2013, 1, 318–324. [Google Scholar] [CrossRef]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Munyoki, S.K.; Sukhwani, M.; Schmid, N.; Missel, A.; Emery, B.R.; DonorConnect; Stukenborg, J.B.; Mayerhofer, A.; Orwig, K.E.; et al. Single-cell analysis of human testis aging and correlation with elevated body mass index. Dev. Cell 2022, 57, 1160–1176.e5. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Ramsch, R.; Kohn, F.M.; Schwarzer, J.U.; Mayerhofer, A. Isolation and cultivation of human testicular peritubular cells: A new model for the investigation of fibrotic processes in the human testis and male infertility. J. Clin. Endocrinol. Metab. 2006, 91, 1956–1960. [Google Scholar] [CrossRef] [Green Version]
- Fleck, D.; Kenzler, L.; Mundt, N.; Strauch, M.; Uesaka, N.; Moosmann, R.; Bruentgens, F.; Missel, A.; Mayerhofer, A.; Merhof, D.; et al. ATP activation of peritubular cells drives testicular sperm transport. elife 2021, 10, e62885. [Google Scholar] [CrossRef] [PubMed]
- Walenta, L.; Fleck, D.; Frohlich, T.; von Eysmondt, H.; Arnold, G.J.; Spehr, J.; Schwarzer, J.U.; Köhn, F.M.; Spehr, M.; Mayerhofer, A. ATP-mediated Events in Peritubular Cells Contribute to Sterile Testicular Inflammation. Sci. Rep. 2018, 8, 1431. [Google Scholar] [CrossRef]
- Chi, J.T.; Rodriguez, E.H.; Wang, Z.; Nuyten, D.S.; Mukherjee, S.; van de Rijn, M.; van de Vijver, M.J.; Hastie, T.; Brown, P.O. Gene expression programs of human smooth muscle cells: Tissue-specific differentiation and prognostic significance in breast cancers. PLoS Genet. 2007, 3, 1770–1784. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.; Flenkenthaler, F.; Stockl, J.B.; Dietrich, K.G.; Köhn, F.M.; Schwarzer, J.U.; Kunz, L.; Luckner, M.; Wanner, G.; Arnold, G.J.; et al. Insights into replicative senescence of human testicular peritubular cells. Sci. Rep. 2019, 9, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missel, A.; Walenta, L.; Eubler, K.; Mundt, N.; Heikelä, H.; Pickl, U.; Trottmann, M.; Popper, B.; Poutanen, M.; Strauss, L.; et al. Testicular adenosine acts as a pro-inflammatory molecule: Role of testicular peritubular cells. Mol Hum. Reprod. 2021, 27, gaab037. [Google Scholar] [CrossRef]
- Luecken, M.D.; Theis, F.J. Current best practices in single-cell RNA-seq analysis: A tutorial. Mol. Syst. Biol. 2019, 15, e8746. [Google Scholar] [CrossRef]
- Wolock, S.L.; Lopez, R.; Klein, A.M. Scrublet: Computational Identification of Cell Doublets in Single-Cell Transcriptomic Data. Cell Syst. 2019, 8, 281–291 e289. [Google Scholar] [CrossRef] [Green Version]
- Lun, A.T.; Bach, K.; Marioni, J.C. Pooling across cells to normalize single-cell RNA sequencing data with many zero counts. Genome Biol. 2016, 17, 75. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.X.; Terry, J.M.; Belgrader, P.; Ryvkin, P.; Bent, Z.W.; Wilson, R.; Ziraldo, S.B.; Wheeler, T.D.; McDermott, G.P.; Zhu, J.; et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 2017, 8, 14049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hie, B.; Bryson, B.; Berger, B. Efficient integration of heterogeneous single-cell transcriptomes using Scanorama. Nat. Biotechnol. 2019, 37, 685–691. [Google Scholar] [CrossRef]
- Traag, V.A.; Waltman, L.; van Eck, N.J. From Louvain to Leiden: Guaranteeing well-connected communities. Sci. Rep. 2019, 9, 5233. [Google Scholar] [CrossRef] [Green Version]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar]
- Tirosh, I.; Izar, B.; Prakadan, S.M.; Wadsworth, M.H.; Treacy, D.; Trombetta, J.J.; Rotem, A.; Rodman, C.; Lian, C.; Murphy, G.; et al. Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-seq. Science 2016, 352, 189–196. [Google Scholar] [CrossRef]
- La Manno, G.; Soldatov, R.; Zeisel, A.; Braun, E.; Hochgerner, H.; Petukhov, V.; Lidschreiber, K.; Kastriti, M.E.; Lonnerberg, P.; Furlan, A.; et al. RNA velocity of single cells. Nature 2018, 560, 494–498. [Google Scholar] [CrossRef] [Green Version]
- Bergen, V.; Lange, M.; Peidli, S.; Wolf, F.A.; Theis, F.J. Generalizing RNA velocity to transient cell states through dynamical modeling. Nat. Biotechnol. 2020, 38, 1408–1414. [Google Scholar] [CrossRef]
- Shen, Y.C.; Shami, A.N.; Moritz, L.; Larose, H.; Manske, G.L.; Ma, Q.; Zheng, X.; Sukhwani, M.; Czerwinski, M.; Sultan, C.; et al. TCF21(+) mesenchymal cells contribute to testis somatic cell development, homeostasis, and regeneration in mice. Nat. Commun. 2021, 12, 3876. [Google Scholar] [CrossRef] [PubMed]
- Welter, H.; Herrmann, C.; Dellweg, N.; Missel, A.; Thanisch, C.; Urbanski, H.F.; Köhn, F.M.; Schwarzer, J.U.; Müller-Taubenberger, A.; Mayerhofer, A. The Glucocorticoid Receptor NR3C1 in Testicular Peritubular Cells is Developmentally Regulated and Linked to the Smooth Muscle-Like Cellular Phenotype. J. Clin. Med. 2020, 9, 961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadon, M.; Santoro, M.M. The origin and mechanisms of smooth muscle cell development in vertebrates. Development 2021, 148, dev197384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, X.; Xia, Y.; Mao, L. An update on the phenotypic switching of vascular smooth muscle cells in the pathogenesis of atherosclerosis. Cell Mol. Life Sci. 2021, 79, 6. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Adam, M.; Glashauser, L.; Dietrich, K.; Schwarzer, J.U.; Köhn, F.M.; Strauss, L.; Welter, H.; Poutanen, M.; Mayerhofer, A. Sterile inflammation as a factor in human male infertility: Involvement of Toll like receptor 2, biglycan and peritubular cells. Sci. Rep. 2016, 6, 37128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flenkenthaler, F.; Windschüttl, S.; Fröhlich, T.; Schwarzer, J.U.; Mayerhofer, A.; Arnold, G.J. Secretome analysis of testicular peritubular cells: A window into the human testicular microenvironment and the spermatogonial stem cell niche in man. J. Proteome Res. 2014, 13, 1259–1269. [Google Scholar] [CrossRef]
- Mayer, C.; Adam, M.; Walenta, L.; Schmid, N.; Heikelä, H.; Schubert, K.; Flenkenthaler, F.; Dietrich, K.G.; Gruschka, S.; Arnold, G.J.; et al. Insights into the role of androgen receptor in human testicular peritubular cells. Andrology 2018, 6, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Landreh, L.; Spinnler, K.; Schubert, K.; Hakkinen, M.R.; Auriola, S.; Poutanen, M.; Soder, O.; Svechnikov, K.; Mayerhofer, A. Human testicular peritubular cells host putative stem Leydig cells with steroidogenic capacity. J. Clin. Endocrinol. Metab. 2014, 99, E1227–E1235. [Google Scholar] [CrossRef]
- Eubler, K.; Herrmann, C.; Tiefenbacher, A.; Köhn, F.M.; Schwarzer, J.U.; Kunz, L.; Mayerhofer, A. Ca(2+) Signaling and IL-8 Secretion in Human Testicular Peritubular Cells Involve the Cation Channel TRPV2. Int. J. Mol. Sci. 2018, 19, 2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Y.; Willis, W.D.; Eddy, E.M. Targeting the Gdnf Gene in peritubular myoid cells disrupts undifferentiated spermatogonial cell development. Proc. Natl. Acad. Sci. USA 2016, 113, 1829–1834. [Google Scholar] [CrossRef] [Green Version]
- Spinnler, K.; Kohn, F.M.; Schwarzer, U.; Mayerhofer, A. Glial cell line-derived neurotrophic factor is constitutively produced by human testicular peritubular cells and may contribute to the spermatogonial stem cell niche in man. Hum. Reprod. 2010, 25, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Ma, Y.; Feng, X.; Deng, R.; Ke, Q.; Xiang, A.P.; Deng, C. Endosialin defines human stem Leydig cells with regenerative potential. Hum. Reprod. 2020, 35, 2197–2212. [Google Scholar] [CrossRef]
- Zhao, X.; Wen, X.; Ji, M.; Guan, X.; Chen, P.; Hao, X.; Chen, F.; Hu, Y.; Duan, P.; Ge, R.S.; et al. Differentiation of seminiferous tubule-associated stem cells into leydig cell and myoid cell lineages. Mol. Cell Endocrinol. 2021, 525, 111179. [Google Scholar] [CrossRef]
- Dobnikar, L.; Taylor, A.L.; Chappell, J.; Oldach, P.; Harman, J.L.; Oerton, E.; Dzierzak, E.; Bennett, M.R.; Spivakov, M.; Jorgensen, H.F. Disease-relevant transcriptional signatures identified in individual smooth muscle cells from healthy mouse vessels. Nat. Commun. 2018, 9, 4567. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Gomez, D. Smooth Muscle Cell Phenotypic Diversity. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1715–1723. [Google Scholar] [CrossRef]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef]
- Stöckl, J.B.; Schmid, N.; Flenkenthaler, F.; Drummer, C.; Behr, R.; Mayerhofer, A.; Arnold, G.J.; Fröhlich, T. Proteomic Insights into Senescence of Testicular Peritubular Cells from a Nonhuman Primate Model. Cells 2020, 9, 2498. [Google Scholar] [CrossRef]
- Davidoff, M.S.; Breucker, H.; Holstein, A.F.; Seidl, K. Cellular architecture of the lamina propria of human seminiferous tubules. Cell Tissue Res. 1990, 262, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Holstein, A.F.; Maekawa, M.; Nagano, T.; Davidoff, M.S. Myofibroblasts in the lamina propria of human semi-niferous tubules are dynamic structures of heterogeneous phenotype. Arch. Histol. Cytol. 1996, 59, 109–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B. The myofibroblast: Paradigm for a mechanically active cell. J. Biomech. 2010, 43, 146–155. [Google Scholar] [CrossRef]
- Pakshir, P.; Noskovicova, N.; Lodyga, M.; Son, D.O.; Schuster, R.; Goodwin, A.; Karvonen, H.; Hinz, B. The myofibroblast at a glance. J. Cell Sci. 2020, 133, jcs227900. [Google Scholar] [CrossRef]
- Zhu, D.; Hadoke, P.W.; Wu, J.; Vesey, A.T.; Lerman, D.A.; Dweck, M.R.; Newby, D.E.; Smith, L.B.; MacRae, V.E. Ablation of the androgen receptor from vascular smooth muscle cells demonstrates a role for testosterone in vascular calcification. Sci. Rep. 2016, 6, 24807. [Google Scholar] [CrossRef] [Green Version]
- Abe, S.I. Behavior and Functional Roles of CD34(+) Mesenchymal Cells in Mammalian Testes. Int. J. Mol. Sci. 2022, 23, 9585. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Guasti, D.; Gacci, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Reappraising the microscopic anatomy of human testis: Identification of telocyte networks in the peritubular and intertubular stromal space. Sci. Rep. 2018, 8, 14780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliveld, J.; van Daalen, S.K.M.; de Winter-Korver, C.M.; van der Veen, F.; Repping, S.; Teerds, K.; van Pelt, A.M.M. A comparative analysis of human adult testicular cells expressing stem Leydig cell markers in the interstitium, vasculature, and peritubular layer. Andrology 2020, 8, 1265–1276. [Google Scholar] [CrossRef]
- Huang, H.; Zhong, L.; Zhou, J.; Hou, Y.; Zhang, Z.; Xing, X.; Sun, J. Leydig-like cells derived from reprogrammed human foreskin fibroblasts by CRISPR/dCas9 increase the level of serum testosterone in castrated male rats. J. Cell Mol. Med. 2020, 24, 3971–3981. [Google Scholar] [CrossRef]
- Liu, Z.J.; Liu, Y.H.; Huang, S.Y.; Zang, Z.J. Insights into the Regulation on Proliferation and Differentiation of Stem Leydig Cells. Stem Cell Rev. Rep. 2021, 17, 1521–1533. [Google Scholar] [CrossRef] [PubMed]
- Schulze, C.; Holstein, A.F. Leydig cells within the lamina propria of seminiferous tubules in four patients with azoospermia. Andrologia 1978, 10, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wu, J.; Liu, B.; Jiang, Y.; Chen, W.; Li, J.; He, Q.; He, Z. The roles and mechanisms of Leydig cells and myoid cells in regulating spermatogenesis. Cell Mol. Life Sci. 2019, 76, 2681–2695. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Park, G.; Hong, H.S. Age affects the paracrine activity and differentiation potential of human adiposederived stem cells. Mol. Med. Rep. 2021, 23, 160. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Duan, Y.; Li, Y.; Sun, Y.; Sun, H.; Yu, X.; Gao, X.; Zhang, C.; Zhang, H.; et al. Metabolic Regulation: A Potential Strategy for Rescuing Stem Cell Senescence. Stem Cell Rev. Rep. 2022, 18, 1728–1742. [Google Scholar] [CrossRef]
- Mularoni, V.; Esposito, V.; Di Persio, S.; Vicini, E.; Spadetta, G.; Berloco, P.; Fanelli, F.; Mezzullo, M.; Pagotto, U.; Pelusi, C.; et al. Age-related changes in human Leydig cell status. Hum. Reprod. 2020, 35, 2663–2676. [Google Scholar] [CrossRef]
- Chen, L.Y.; Brown, P.R.; Willis, W.B.; Eddy, E.M. Peritubular myoid cells participate in male mouse spermatogonial stem cell maintenance. Endocrinology 2014, 155, 4964–4974. [Google Scholar] [CrossRef] [Green Version]
- Rey-Ares, V.; Rossi, S.P.; Dietrich, K.G.; Kohn, F.M.; Schwarzer, J.U.; Welter, H.; Frungieri, M.B.; Mayerhofer, A. Prostaglandin E2 (PGE2) is a testicular peritubular cell-derived factor involved in human testicular homeostasis. Mol. Cell Endocrinol. 2018, 473, 217–224. [Google Scholar] [CrossRef]
- Endo, T.; Mikedis, M.M.; Nicholls, P.K.; Page, D.C.; de Rooij, D.G. Retinoic Acid and Germ Cell Development in the Ovary and Testis. Biomolecules 2019, 9, 775. [Google Scholar] [CrossRef] [Green Version]
- Griswold, M.D. Spermatogenesis: The Commitment to Meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, E.; Sato, S.; Honda, S.; Nakazawa, A. All trans retinoic acid alleviates coronary stenosis by regulating smooth muscle cell function in a mouse model of Kawasaki disease. Sci. Rep. 2021, 11, 13856. [Google Scholar] [CrossRef] [PubMed]
- Saracino, R.; Capponi, C.; Di Persio, S.; Boitani, C.; Masciarelli, S.; Fazi, F.; Fera, S.; Vicini, E. Regulation of Gdnf expression by retinoic acid in Sertoli cells. Mol. Reprod. Dev. 2020, 87, 419–429. [Google Scholar] [CrossRef] [PubMed]







| Gene Name | Description | HTPC-1 | HTPC-2 | TPCs of Nie et al. [17] | TPCs of Di Persio et al. [3] |
|---|---|---|---|---|---|
| ACTA2 | Smooth-muscle actin | √ | √ | √ | √ |
| MYH11 | Myosin heavy chain 11 | √ | √ | √ | √ |
| TAGLN | Transgelin | √ | √ | √ | √ |
| CNN1 | Calponin 1 | √ | √ | ✕ | √ |
| CALD1 | Caldesmon 1 | √ | √ | √ | √ |
| SRF | Serum response factor | √ | √ | √ | √ |
| MYOCD | Myocardin | (√) | ✕ | √ | √ |
| VIM | Vimentin | √ | √ | √ | √ |
| DES | Desmin | √ | √ | √ | √ |
| SMTN | Smoothelin | √ | √ | √ | √ |
| PDGFRB | Platelet derived growth factor receptor B | √ | √ | √ | √ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebich, A.; Schmid, N.; Koupourtidou, C.; Herrmann, C.; Dietrich, K.-G.; Welter, H.; Ninkovic, J.; Mayerhofer, A. The Molecular Signature of Human Testicular Peritubular Cells Revealed by Single-Cell Analysis. Cells 2022, 11, 3685. https://doi.org/10.3390/cells11223685
Liebich A, Schmid N, Koupourtidou C, Herrmann C, Dietrich K-G, Welter H, Ninkovic J, Mayerhofer A. The Molecular Signature of Human Testicular Peritubular Cells Revealed by Single-Cell Analysis. Cells. 2022; 11(22):3685. https://doi.org/10.3390/cells11223685
Chicago/Turabian StyleLiebich, Annika, Nina Schmid, Christina Koupourtidou, Carola Herrmann, Kim-Gwendolyn Dietrich, Harald Welter, Jovica Ninkovic, and Artur Mayerhofer. 2022. "The Molecular Signature of Human Testicular Peritubular Cells Revealed by Single-Cell Analysis" Cells 11, no. 22: 3685. https://doi.org/10.3390/cells11223685