
Thymosin Beta 15 Alters the Spatial Development of Thymic Epithelial Cells
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cells
2.2. Cell Infection, Transfection, and Screening
2.3. In Vitro Mouse Thymus Organ Culture System
2.4. Murine Artificial Thymic Organoid Cultures
2.5. Subcutaneous Transplantation Experiments of Thymic Organoids
2.6. Immunofluorescence Imaging
2.7. Histology and Morphometric Analysis
2.8. Flow Cytometry
2.9. Cell Cycle Analysis
2.10. Apoptosis Detection
2.11. Proliferation Assay
2.12. RNA Isolation, Reverse Transcription and Quantitative Real-Time PCR
2.13. TREC Quantification
2.14. Cell Self-Renewal, Proliferation, and Viability Assays
2.15. Migration Assays
2.16. Statistical Analysis
3. Results
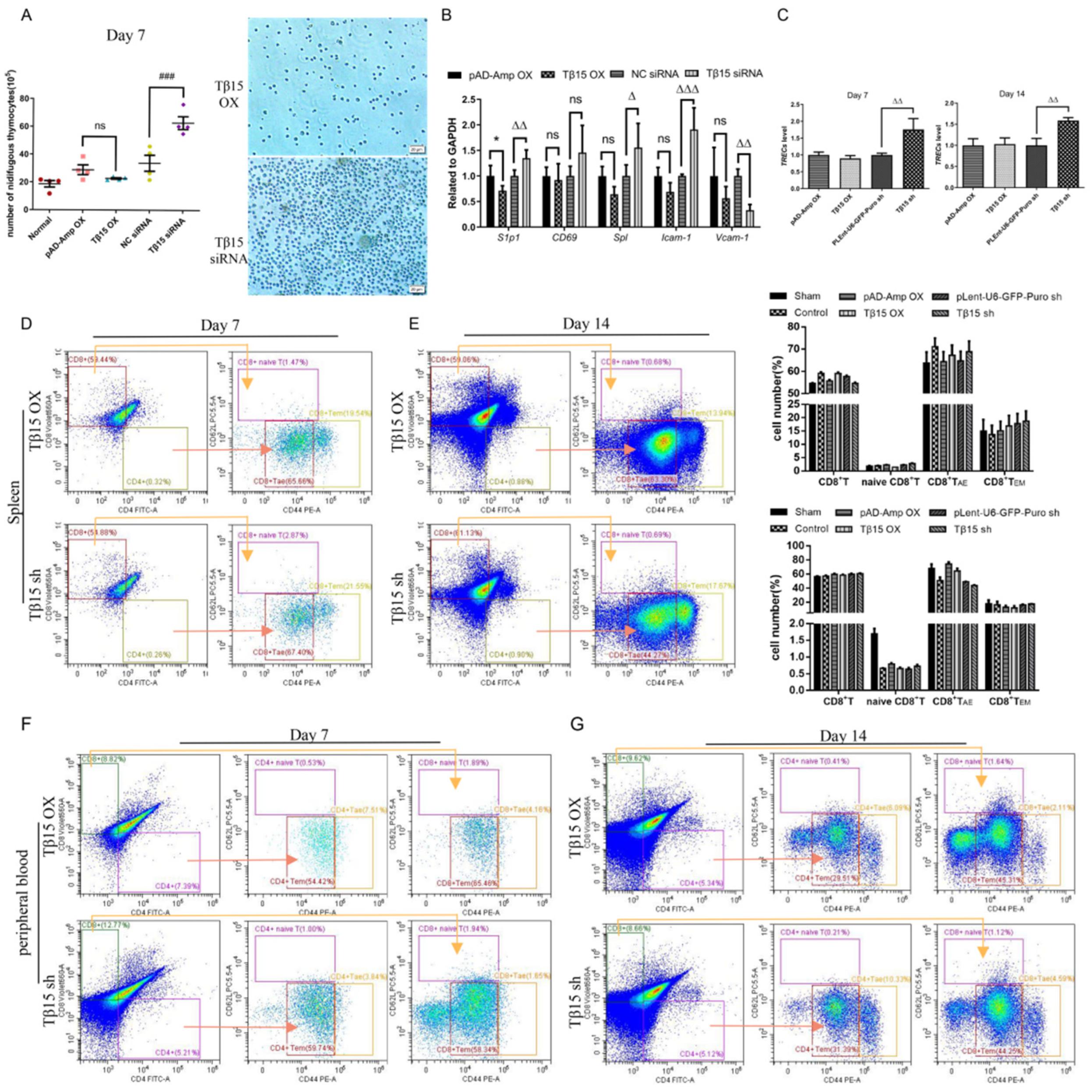
3.1. Tβ15 Influences the Maturation of Thymus by Acting on TECs
3.2. Tβ15 Inhibits the Development of mTECs and Suppresses the Formation of Reticulations in TECs
3.3. Tβ15 Delays the Development of CD4SP Thymocytes in Thymus
3.4. Tβ15 Level in iTECs Influences Thymic Output and Activation of T Cells
3.5. Tβ15 Regulates Thymocyte Fate via Inhibiting the Directed Reticular Differentiation of TECs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aw, D.; Taylor-Brown, F.; Cooper, K.; Palmer, D.B. Phenotypical and morphological changes in the thymic microenvironment from ageing mice. Biogerontology 2009, 10, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Barbouti, A.; Vasileiou, P.V.S.; Evangelou, K.; Vlasis, K.G.; Papoudou-Bai, A.; Gorgoulis, V.G.; Kanavaros, P. Implications of Oxidative Stress and Cellular Senescence in Age-Related Thymus Involution. Oxid Med. Cell Longev. 2020, 2020, 7986071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gameiro, J.; Nagib, P.; Verinaud, L. The thymus microenvironment in regulating thymocyte differentiation. Cell Adhes. Migr. 2010, 4, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Shui, Y.; Xu, X.; He, K.; Yang, F.; Gao, J. Thymic function affects breast cancer development and metastasis by regulating expression of thymus secretions PTMα and Tβ15b1. Transl. Oncol. 2021, 14, 100980. [Google Scholar] [CrossRef] [PubMed]
- Buus, T.B.; Ødum, N.; Geisler, C.; Lauritsen, J.P.H. Three distinct developmental pathways for adaptive and two IFN-γ-producing γδ T subsets in adult thymus. Nat. Commun. 2017, 8, 1911. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Liu, C.; Gong, Y.; Bai, Z.; Hou, S.; He, J.; Bian, Z.; Li, Z.; Ni, Y.; Yan, J.; et al. Single-Cell RNA Sequencing Resolves Spatiotemporal Development of Pre-thymic Lymphoid Progenitors and Thymus Organogenesis in Human Embryos. Immunity 2019, 51, 930–948.e6. [Google Scholar] [CrossRef]
- Rausch, M.P.; Meador, L.R.; Metzger, T.C.; Li, H.; Qiu, S.; Anderson, M.S.; Hastings, K.T. GILT in Thymic Epithelial Cells Facilitates Central CD4 T Cell Tolerance to a Tissue-Restricted, Melanoma-Associated Self-Antigen. J. Immunol. 2020, 204, 2877–2886. [Google Scholar] [CrossRef]
- Rossi, S.W.; Jenkinson, W.E.; Anderson, G.; Jenkinson, E.J. Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 2006, 441, 988–991. [Google Scholar] [CrossRef]
- Takahama, Y.; Ohigashi, I.; Baik, S.; Anderson, G. Generation of diversity in thymic epithelial cells. Nat. Rev. Immunol. 2017, 17, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Abramson, J.; Anderson, G. Thymic Epithelial Cells. Annu. Rev. Immunol. 2017, 35, 85–118. [Google Scholar] [CrossRef]
- Wells, K.L.; Miller, C.N.; Gschwind, A.R.; Wei, W.; Phipps, J.D.; Anderson, M.S.; Steinmetz, L.M. Combined transient ablation and single-cell RNA-sequencing reveals the development of medullary thymic epithelial cells. eLife 2020, 9, e60188. [Google Scholar] [CrossRef] [PubMed]
- Eshima, K.; Misawa, K.; Ohashi, C.; Noma, H.; Iwabuchi, K. NF-κB-inducing kinase contributes to normal development of cortical thymic epithelial cells: Its possible role in shaping a proper T-cell repertoire. Immunology 2020, 160, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.L.; Slater, F.D.; White, A. Preparation, assay, and partial purification of a thymic lymphocytopoietic factor (thymosin). Proc. Natl. Acad. Sci. USA 1966, 56, 1010–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, S.S.; Wang, J.; Li, B.X.; Yang, G.W.; Sun, J.J.; Yang, H.T. Thymosin participates in antimicrobial immunity in zebrafish. Fish Shellfish Immunol. 2019, 87, 371–378. [Google Scholar] [CrossRef]
- Maar, K.; Hetenyi, R.; Maar, S.; Faskerti, G.; Hanna, D.; Lippai, B.; Takatsy, A.; Bock-Marquette, I. Utilizing Developmentally Essential Secreted Peptides Such as Thymosin Beta-4 to Remind the Adult Organs of Their Embryonic State-New Directions in Anti-Aging Regenerative Therapies. Cells 2021, 10, 1343. [Google Scholar] [CrossRef]
- Xue, B.; Robinson, R.C. Actin-Induced Structure in the Beta-Thymosin Family of Intrinsically Disordered Proteins. Vitam. Horm. 2016, 102, 55–71. [Google Scholar]
- Ninomiya, K.; Ohta, K.; Yamashita, K.; Mizuno, K.; Ohashi, K. PLEKHG4B enables actin cytoskeletal remodeling during epithelial cell-cell junction formation. J. Cell Sci. 2021, 134, jcs249078. [Google Scholar] [CrossRef]
- Bao, L.; Loda, M.; Janmey, P.A.; Stewart, R.; Anand-Apte, B.; Zetter, B.R. Thymosin beta 15: A novel regulator of tumor cell motility upregulated in metastatic prostate cancer. Nat. Med. 1996, 2, 1322–1328. [Google Scholar] [CrossRef]
- Koutrafouri, V.; Leondiadis, L.; Ferderigos, N.; Avgoustakis, K.; Livaniou, E.; Evangelatos, G.P.; Ithakissios, D.S. Synthesis and angiogenetic activity in the chick chorioallantoic membrane model of thymosin beta-15. Peptides 2003, 24, 107–115. [Google Scholar] [CrossRef]
- Shen, J.M.; Ma, L.; He, K.; Guo, W.Q.; Ding, C.; Hoffman, R.D.; He, B.Q.; Zheng, H.B.; Gao, J.L. Identification and functional study of immortalized mouse thymic epithelial cells. Biochem. Biophys. Res. Commun. 2020, 525, 440–446. [Google Scholar] [CrossRef]
- Montel-Hagen, A.; Sun, V.; Casero, D.; Tsai, S.; Zampieri, A.; Jackson, N.; Li, S.; Lopez, S.; Zhu, Y.; Chick, B.; et al. In Vitro Recapitulation of Murine Thymopoiesis from Single Hematopoietic Stem Cells. Cell Rep. 2020, 33, 108320. [Google Scholar] [CrossRef] [PubMed]
- Pignata, C.; D’Assante, R.; Sousa, A.E. Thymic stromal alterations and genetic disorders of immune system. Front. Immunol. 2015, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Abercrombie, M.; Heaysman, J.E.; Pegrum, S.M. The locomotion of fibroblasts in culture. I. Movements of the leading edge. Exp. Cell Res. 1970, 59, 393–398. [Google Scholar] [CrossRef]
- Lauffenburger, D.A.; Horwitz, A.F. Cell migration: A physically integrated molecular process. Cell 1996, 84, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Koch, U.; Radtke, F. Mechanisms of T cell development and transformation. Annu. Rev. Cell Dev. Biol. 2011, 27, 539–562. [Google Scholar] [CrossRef]
- Kretz-Rommel, A.; Rubin, R.L. Disruption of positive selection of thymocytes causes autoimmunity. Nat. Med. 2000, 6, 298–305. [Google Scholar] [CrossRef]
- Zou, L.; Mendez, F.; Martin-Orozco, N.; Peterson, E.J. Defective positive selection results in T cell lymphopenia and increased autoimmune diabetes in ADAP-deficient BDC2.5-C57BL/6 mice. Eur. J. Immunol. 2008, 38, 986–994. [Google Scholar] [CrossRef]
- Takaba, H.; Morishita, Y.; Tomofuji, Y.; Danks, L.; Nitta, T.; Komatsu, N.; Kodama, T.; Takayanagi, H. Fezf2 Orchestrates a Thymic Program of Self-Antigen Expression for Immune Tolerance. Cell 2015, 163, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Tomofuji, Y.; Takaba, H.; Suzuki, H.I.; Benlaribi, R.; Martinez, C.D.P.; Abe, Y.; Morishita, Y.; Okamura, T.; Taguchi, A.; Kodama, T.; et al. Chd4 choreographs self-antigen expression for central immune tolerance. Nat. Immunol. 2020, 21, 892–901. [Google Scholar] [CrossRef]
- Hiramine, C.; Hojo, K.; Koseto, M.; Nakagawa, T.; Mukasa, A. Establishment of a murine thymic epithelial cell line capable of inducing both thymic nurse cell formation and thymocyte apoptosis. Lab. Investig. 1990, 62, 41–54. [Google Scholar] [PubMed]
- Imachi, H.; Murao, K.; Hiramine, C.; Sayo, Y.; Sato, M.; Hosokawa, H.; Ishida, T.; Kodama, T.; Quehenberger, O.; Steinberg, D.; et al. Human scavenger receptor B1 is involved in recognition of apoptotic thymocytes by thymic nurse cells. Lab. Investig. 2000, 80, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, R.A.; Sousa, A.G.G.; Ramos, C.V.; Ávila, M.; Lilue, J.; Paixão, T.; Martins, V.C. Self-renewal of double-negative 3 early thymocytes enables thymus autonomy but compromises the β-selection checkpoint. Cell Rep. 2021, 35, 108967. [Google Scholar] [CrossRef]
- Ramos, C.V.; Ballesteros-Arias, L.; Silva, J.G.; Paiva, R.A.; Nogueira, M.F.; Carneiro, J.; Gjini, E.; Martins, V.C. Cell Competition, the Kinetics of Thymopoiesis, and Thymus Cellularity Are Regulated by Double-Negative 2 to 3 Early Thymocytes. Cell Rep. 2020, 32, 107910. [Google Scholar] [CrossRef] [PubMed]
- Paiva, R.A.; Ramos, C.V.; Martins, V.C. Thymus autonomy as a prelude to leukemia. FEBS J. 2018, 285, 4565–4574. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, K.; Grobe, H.; Cohen, J.; Soffer, A.; Mahly, A.; Adir, O.; Zaidel-Bar, R.; Luxenburg, C. Thymosin β4 is essential for adherens junction stability and epidermal planar cell polarity. Development 2020, 147, dev193425. [Google Scholar] [CrossRef]
- Ying, Y.Y.; Lin, C.; Tao, N.N.; Robert, D.H.; Shi, D.L.; Chen, Z.J.; Gao, J.L. Thymosin β4 and actin: Binding modes, biological functions, and clinical applications. Curr. Protein. Pept. Sci. 2022, in press. [Google Scholar]
- Zhang, J.; Wong, C.C.; Leung, K.T.; Wu, F.; Zhou, Y.; Tong, J.H.M.; Chan, R.C.K.; Li, H.; Wang, Y.; Yan, H.; et al. FGF18-FGFR2 signaling triggers the activation of c-Jun-YAP1 axis to promote carcinogenesis in a subgroup of gastric cancer patients and indicates translational potential. Oncogene 2020, 39, 6647–6663. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Chu, Q.; Jiang, K.; Li, J.; Tang, N. The Strength of Mechanical Forces Determines the Differentiation of Alveolar Epithelial Cells. Dev. Cell 2018, 44, 297–312.e5. [Google Scholar] [CrossRef] [Green Version]
- Brent, A.E.; Rajan, A. Insulin and Leptin/Upd2 Exert Opposing Influences on Synapse Number in Fat-Sensing Neurons. Cell Metab. 2020, 32, 786–800.e7. [Google Scholar] [CrossRef]
- Oswald, J.; Büttner, M.; Jasinski-Bergner, S.; Jacobs, R.; Rosenstock, P.; Kielstein, H. Leptin affects filopodia and cofilin in NK-92 cells in a dose- and time-dependent manner. Eur. J. Histochem. 2018, 62, 2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Kononenko, N.L.; Brüel, A.; Thomsen, J.S.; Poy, M.N. Neuronal Cell Adhesion Molecule 1 Regulates Leptin Sensitivity and Bone Mass. Calcif. Tissue Int. 2018, 102, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Jeong, Y.T.; Jeong, S.R.; Park, M.; Go, H.S.; Kim, M.Y.; Seong, J.K.; Kim, K.W.; Seo, J.T.; Kim, C.H.; et al. Neural regulation of energy and bone homeostasis by the synaptic adhesion molecule Calsyntenin-3. Exp. Mol. Med. 2020, 52, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Pállinger, É.; Kiss, G.A.; Csaba, G. Hormone (ACTH, T3) content of immunophenotyped lymphocyte subpopulations. Acta Microbiol. Immunol. Hung. 2016, 63, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Dittel, L.J.; Dittel, B.N.; Brod, S.A. Ingested ACTH blocks Th17 production by inhibiting GALT IL-6. J. Neurol. Sci. 2020, 409, 116602. [Google Scholar] [CrossRef]
- Nikolouli, E.; Elfaki, Y.; Herppich, S.; Schelmbauer, C.; Delacher, M.; Falk, C.; Mufazalov, I.A.; Waisman, A.; Feuerer, M.; Huehn, J. Recirculating IL-1R2(+) Tregs fine-tune intrathymic Treg development under inflammatory conditions. Cell Mol. Immunol. 2021, 18, 182–193. [Google Scholar] [CrossRef]
- Creighton, B.A.; Afriyie, S.; Ajit, D.; Casingal, C.R.; Voos, K.M.; Reger, J.; Burch, A.M.; Dyne, E.; Bay, J.; Huang, J.K.; et al. Giant ankyrin-B mediates transduction of axon guidance and collateral branch pruning factor sema 3A. eLife 2021, 10, e69815. [Google Scholar] [CrossRef]
- Lins, M.P.; Medeiros, N.C.; Carmo, J.; Porto, F.L.; Dos Santos Reis, M.D.; Smaniotto, S. The Responsiveness of Thymic Stromal Cells to semaphorin-3A. Immunol. Investig. 2022, 51, 395–410. [Google Scholar] [CrossRef]
- Francelin, C.; Geniseli, I.; Nagib, P.; Gameiro, J.; Savino, W.; Verinaud, L. Semaphorin-3A-Related Reduction of Thymocyte Migration in Chemically Induced Diabetic Mice. Neuroimmunomodulation 2020, 27, 28–37. [Google Scholar] [CrossRef]
- Dobeš, J.; Edenhofer, F.; Vobořil, M.; Brabec, T.; Dobešová, M.; Čepková, A.; Klein, L.; Rajewsky, K.; Filipp, D. A novel conditional Aire allele enables cell-specific ablation of the immune tolerance regulator Aire. Eur. J. Immunol. 2018, 48, 546–548. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Nasreen, M.; Seike, T.; Goji, A.; Ohigashi, I.; Miyazaki, T.; Ohta, T.; Kanno, M.; Takahama, Y. IAN family critically regulates survival and development of T lymphocytes. PLoS Biol. 2006, 4, e103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, A.R.; Endale, M.; Lampe, K.; Aksoylar, H.I.; Flagg, A.; Woodgett, J.R.; Hildeman, D.; Jordan, M.B.; Singh, H.; Kucuk, Z.; et al. Gimap5-dependent inactivation of GSK3β is required for CD4(+) T cell homeostasis and prevention of immune pathology. Nat. Commun. 2018, 9, 430. [Google Scholar] [CrossRef]
- Benavides, F.; Venables, A.; Poetschke Klug, H.; Glasscock, E.; Rudensky, A.; Gómez, M.; Martin Palenzuela, N.; Guénet, J.L.; Richie, E.R.; Conti, C.J. The CD4 T cell-deficient mouse mutation nackt (nkt) involves a deletion in the cathepsin L (CtsI) gene. Immunogenetics 2001, 53, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.R.; Katsioudi, G.; Degn, M.; Ejlerskov, P.; Issazadeh-Navikas, S.; Kornum, B.R. DNMT1 regulates expression of MHC class I in post-mitotic neurons. Mol. Brain 2018, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkal, A.A.; Weiskopf, K.; Kao, K.S.; Gordon, S.R.; Rosental, B.; Yiu, Y.Y.; George, B.M.; Markovic, M.; Ring, N.G.; Tsai, J.M.; et al. Engagement of MHC class I by the inhibitory receptor LILRB1 suppresses macrophages and is a target of cancer immunotherapy. Nat. Immunol. 2018, 19, 76–84. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; He, K.; Hoffman, R.D.; Ying, Y.; Tao, N.; Guo, W.; Shen, J.; Liu, X.; Li, M.; Yan, M.; et al. Thymosin Beta 15 Alters the Spatial Development of Thymic Epithelial Cells. Cells 2022, 11, 3679. https://doi.org/10.3390/cells11223679
Xu X, He K, Hoffman RD, Ying Y, Tao N, Guo W, Shen J, Liu X, Li M, Yan M, et al. Thymosin Beta 15 Alters the Spatial Development of Thymic Epithelial Cells. Cells. 2022; 11(22):3679. https://doi.org/10.3390/cells11223679
Chicago/Turabian StyleXu, Xie, Kai He, Robert D. Hoffman, Yuyuan Ying, Nana Tao, Wenqin Guo, Jiaman Shen, Xi Liu, Meiya Li, Meiqiu Yan, and et al. 2022. "Thymosin Beta 15 Alters the Spatial Development of Thymic Epithelial Cells" Cells 11, no. 22: 3679. https://doi.org/10.3390/cells11223679