
Epigenetic Changes within the Annulus Fibrosus by DNA Methylation in Rat Intervertebral Disc Degeneration Model
Abstract
:1. Introduction
2. Materials and Methods
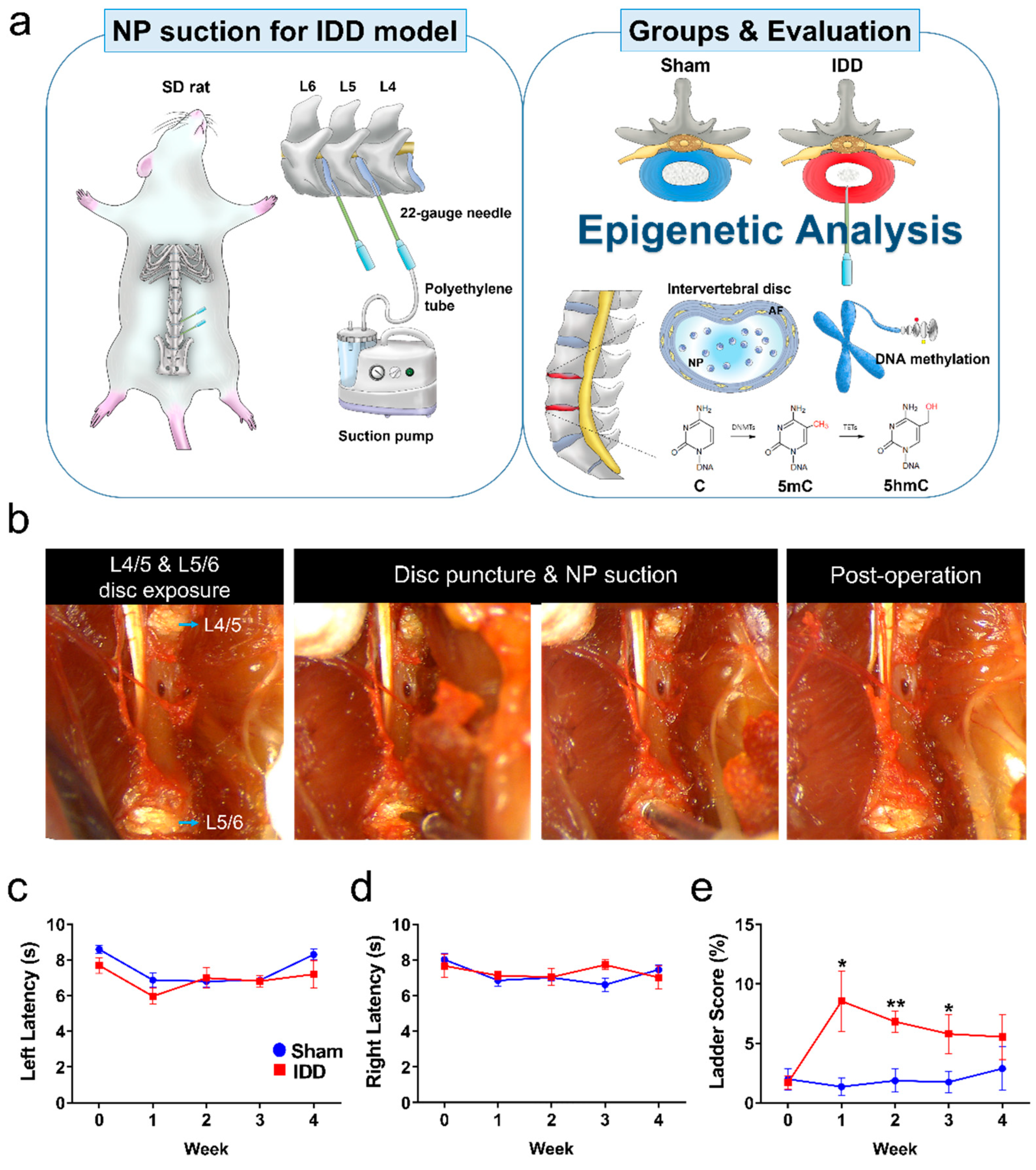
2.1. Intervertebral Disc Degeneration
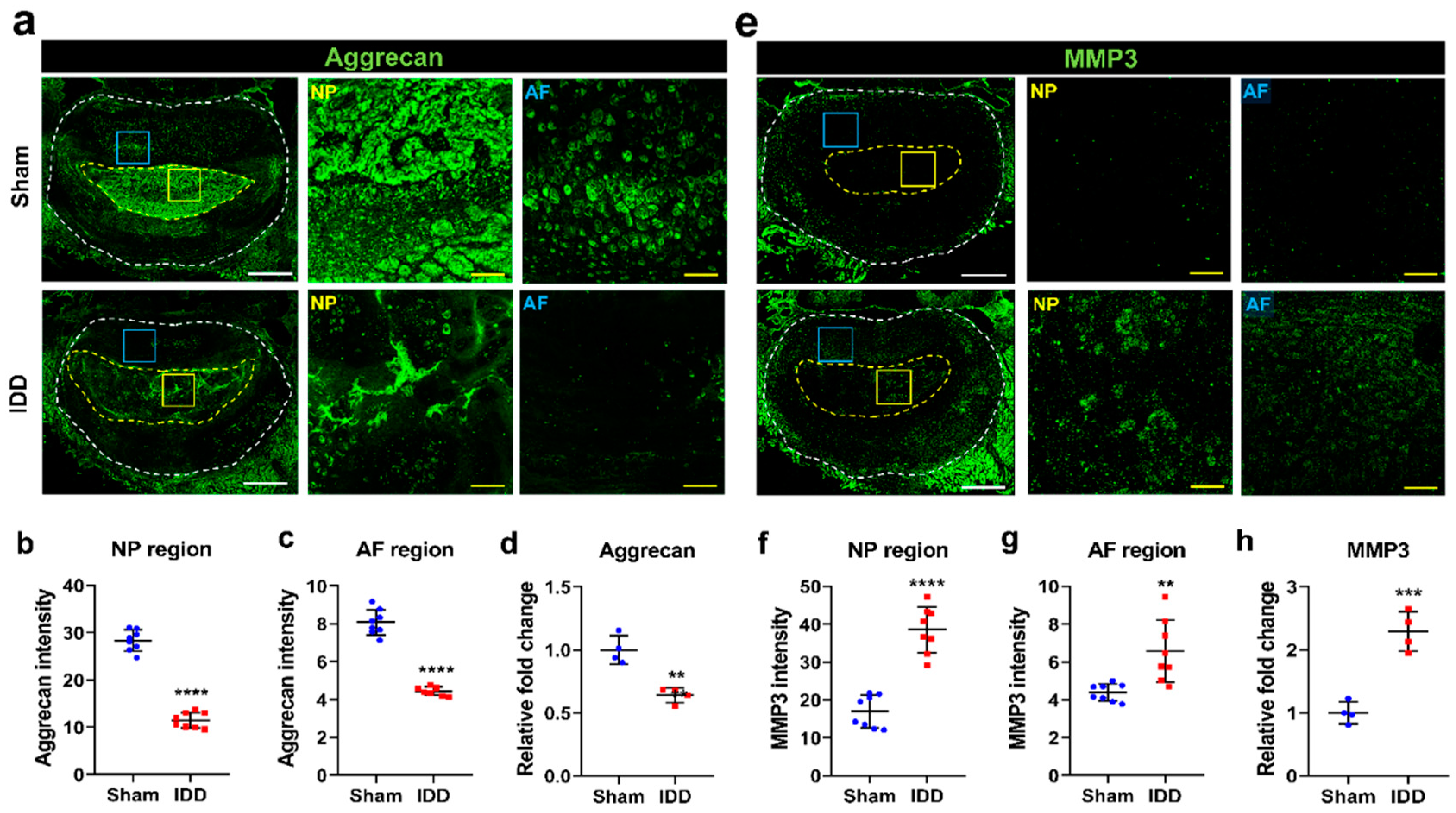
2.2. Immunohistochemistry
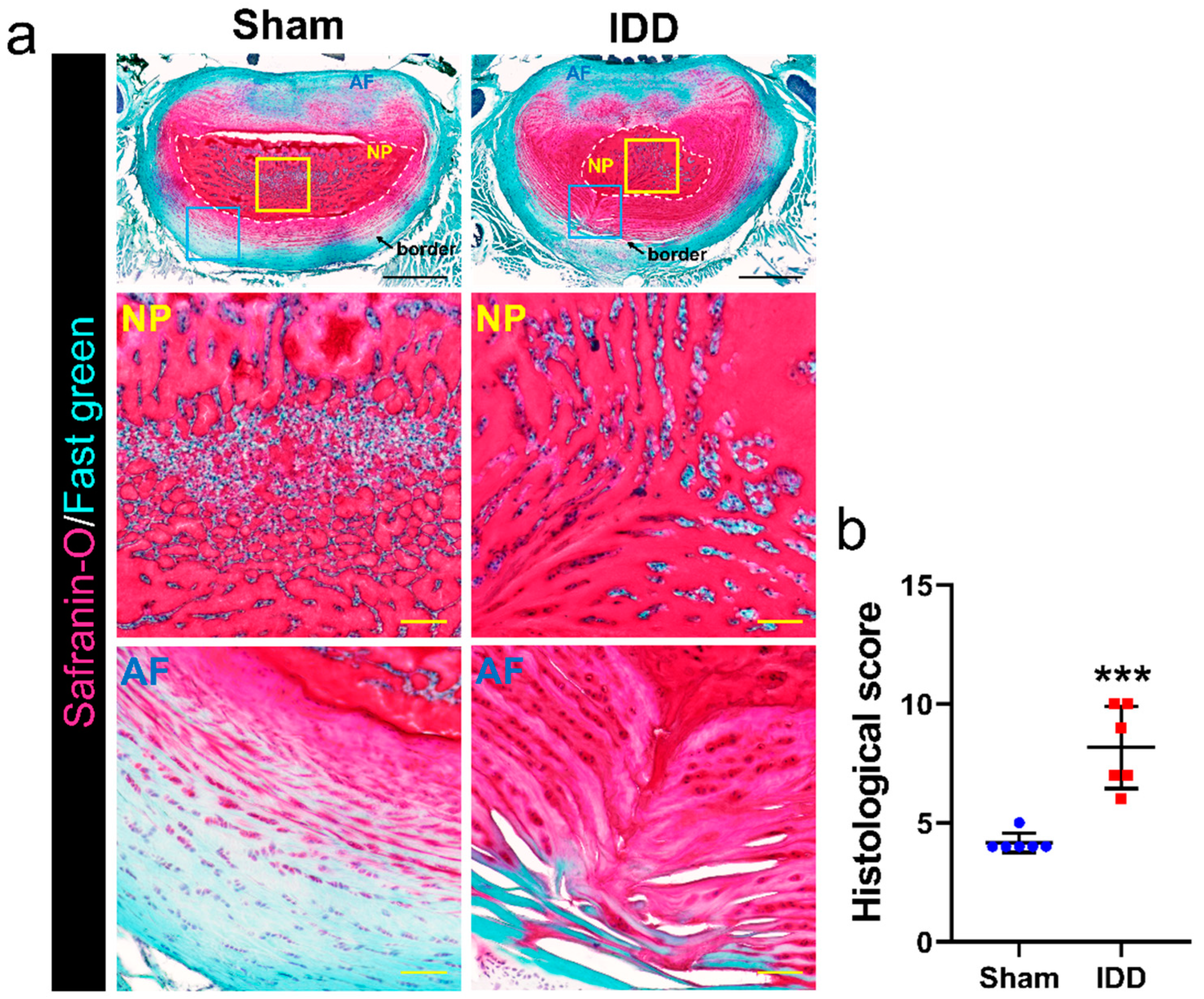
2.3. Safranin O/Fast Green Staining
2.4. Real-Time Polymerase Chain Reaction
2.5. DNA Dot-Blot Assay
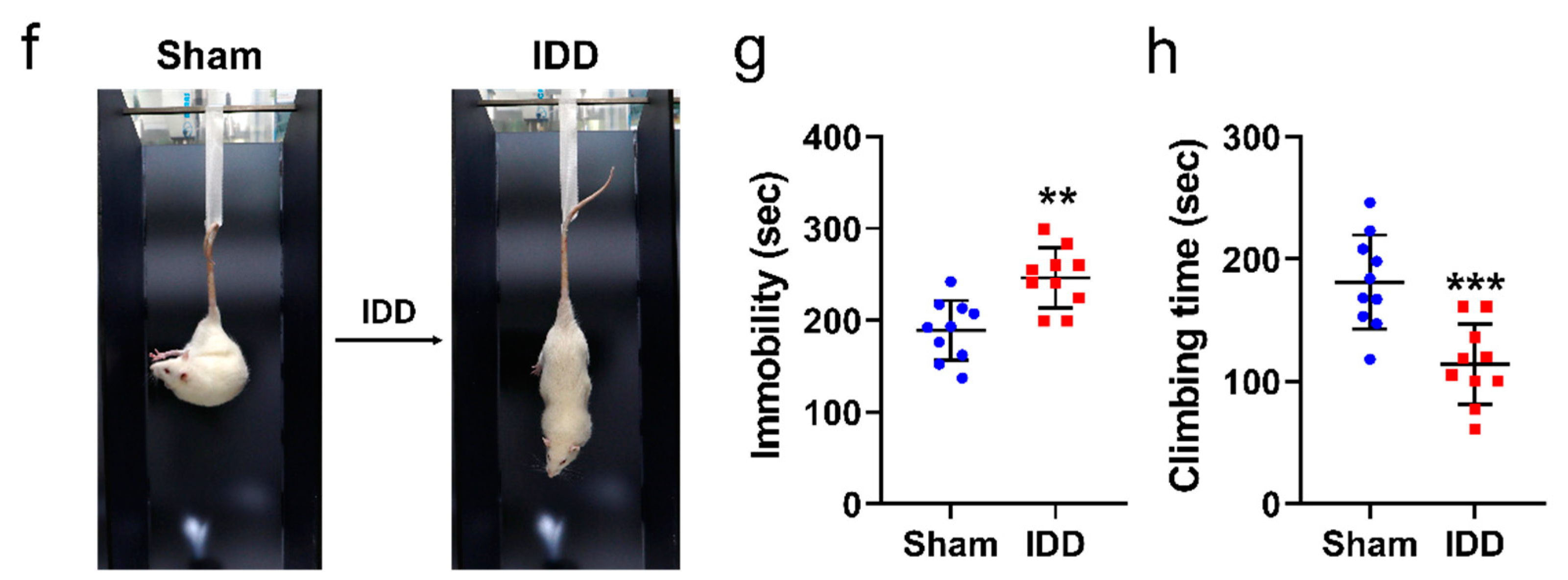
2.6. Functional Assessments
2.7. Statistics
3. Results
3.1. In Vivo Functional Assessment of IDD
3.2. Altered Histological Structure in the Disc after IDD
3.3. IDD Induces Decreased Aggrecan and Increased MMP Levels in the Disc
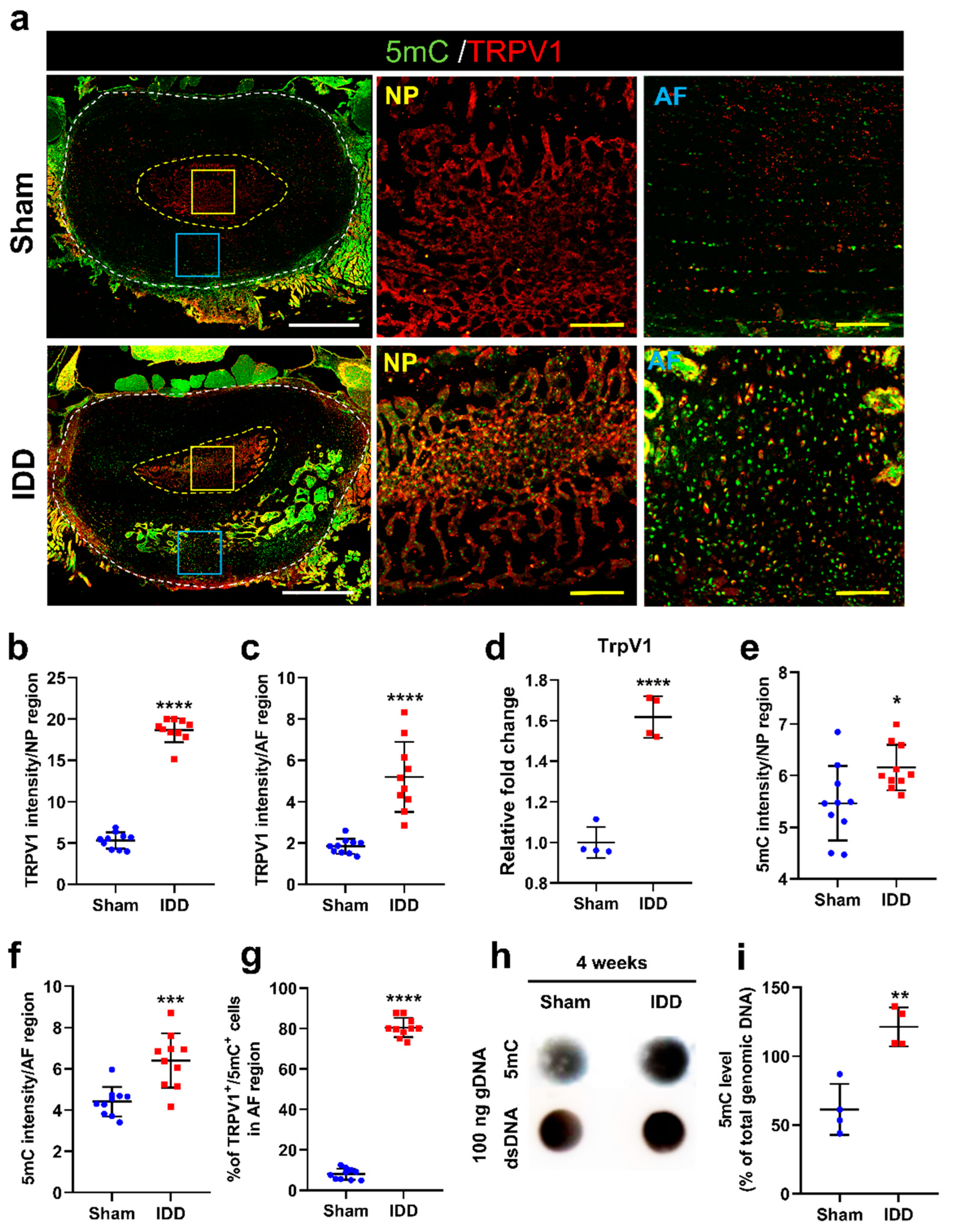
3.4. Increased TrpV1 Expression Was More Clearly Observed with Altered 5mC Levels in the AF Region than in the NP Region after IDD
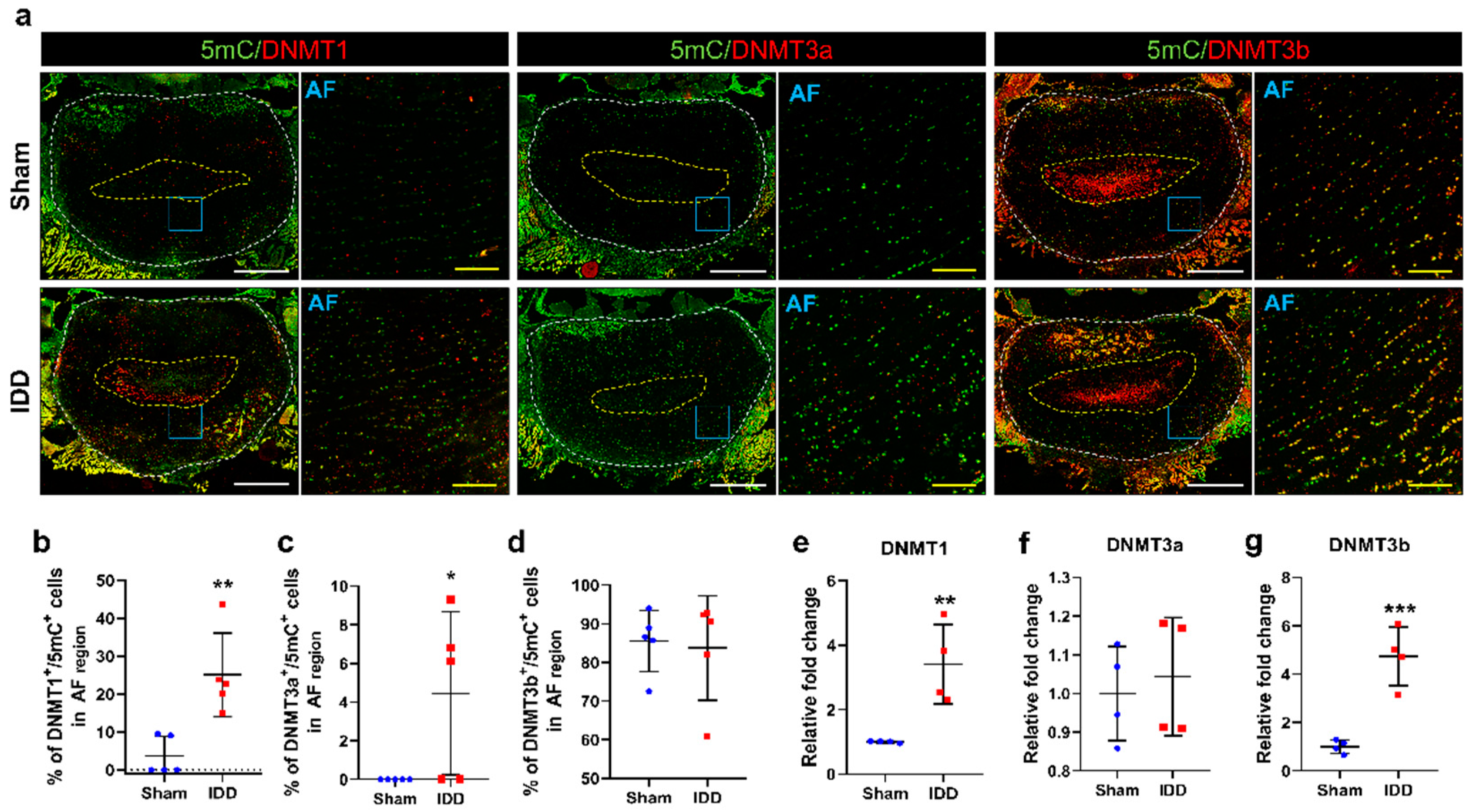
3.5. DNMT3b Was Mainly Expressed in Degenerative Discs and Was Double-Stained with 5mC after IDD
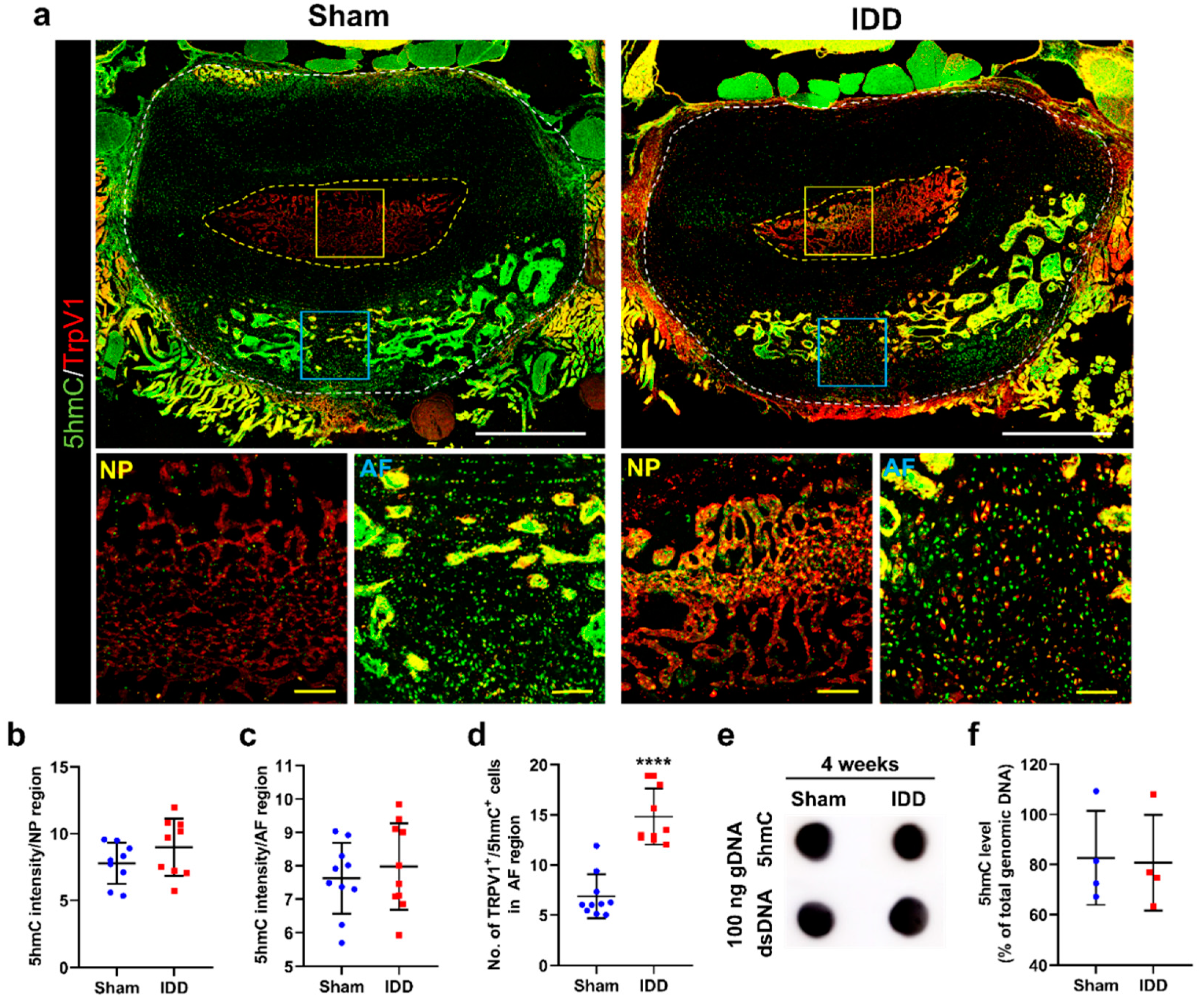
3.6. 5hmC Levels Were Not Altered in the Annulus Fibrosus after IDD
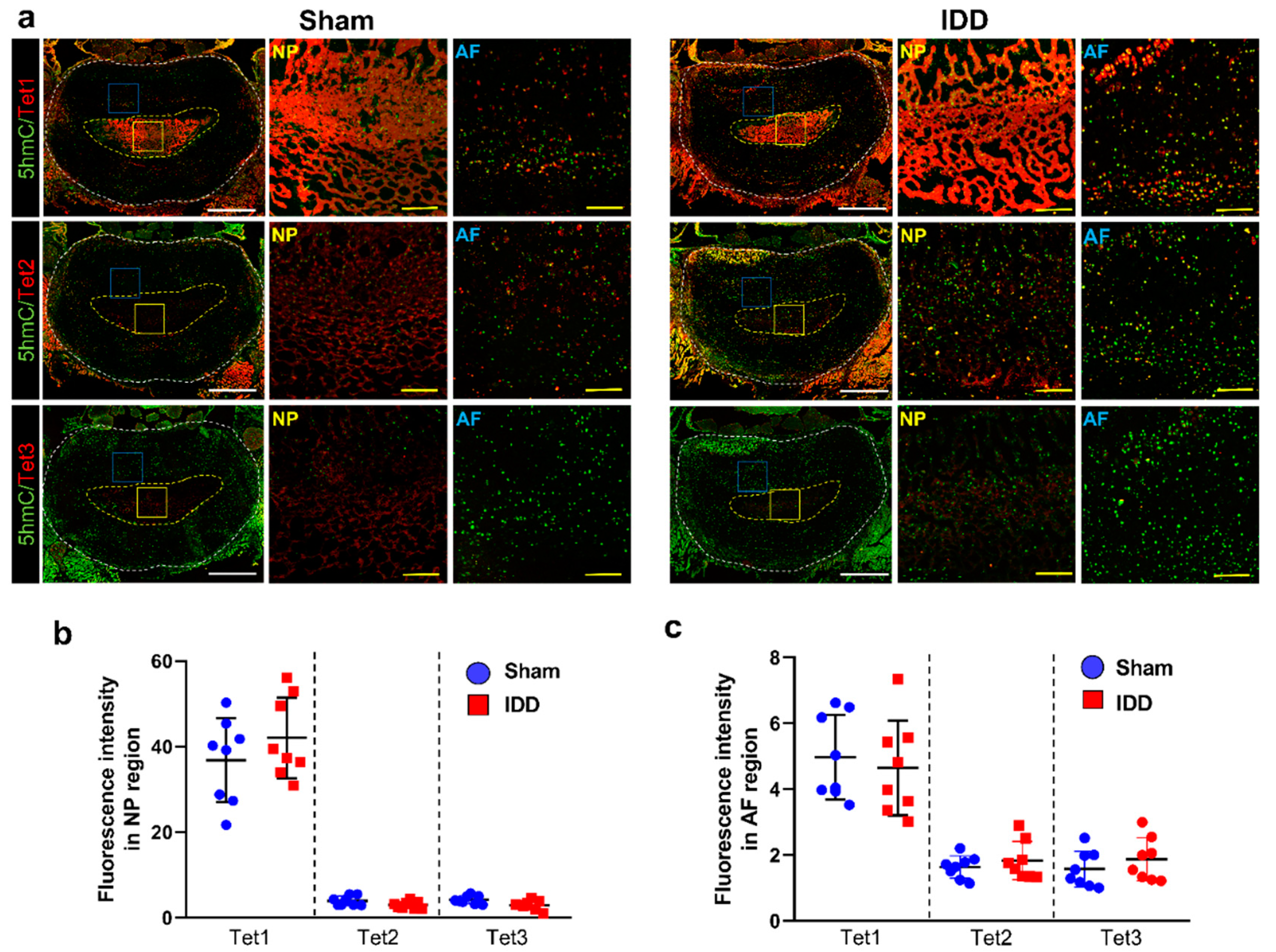
3.7. Tet1 Was Greatly Enhanced in the Disc, although with Similar Induction of DNA Demethylation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Livshits, G.; Popham, M.; Malkin, I.; Sambrook, P.N.; Macgregor, A.J.; Spector, T.; Williams, F.M. Lumbar disc degeneration and genetic factors are the main risk factors for low back pain in women: The UK Twin Spine Study. Ann. Rheum. Dis. 2011, 70, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- Pennicooke, B.; Moriguchi, Y.; Hussain, I.; Bonssar, L.; Hartl, R. Biological Treatment Approaches for Degenerative Disc Disease: A Review of Clinical Trials and Future Directions. Cureus 2016, 8, e892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.G.; Zhang, F.; Sun, Z.; Guo, W.; Liu, J.; Liu, M.; Guo, X. A controlled case study of the relationship between environmental risk factors and apoptotic gene polymorphism and lumbar disc herniation. Am. J. Pathol. 2013, 182, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, N.; Podgorski, M.; Haladaj, R.; Polguj, M.; Olewnik, L. Risk Factors of Intervertebral Disc Pathology-A Point of View Formerly and Today-A Review. J. Clin. Med. 2021, 10, 409. [Google Scholar] [CrossRef]
- Jiang, S.; Guo, Y. Epigenetic Clock: DNA Methylation in Aging. Stem. Cells Int. 2020, 2020, 1047896. [Google Scholar] [CrossRef]
- Saul, D.; Kosinsky, R.L. Epigenetics of Aging and Aging-Associated Diseases. Int. J. Mol. Sci. 2021, 22, 401. [Google Scholar] [CrossRef]
- Turner, B.M. Epigenetic responses to environmental change and their evolutionary implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 3403–3418. [Google Scholar] [CrossRef] [Green Version]
- Bollati, V.; Baccarelli, A. Environmental epigenetics. Heredity 2010, 105, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Kawarai, Y.; Jang, S.H.; Lee, S.; Millecamps, M.; Kang, H.; Gregoire, S.; Suzuki-Narita, M.; Ohtori, S.; Stone, L.S. Exercise attenuates low back pain and alters epigenetic regulation in intervertebral discs in a mouse model. Spine J. 2021, 21, 1938–1949. [Google Scholar] [CrossRef]
- Tajerian, M.; Alvarado, S.; Millecamps, M.; Dashwood, T.; Anderson, K.M.; Haglund, L.; Ouellet, J.; Szyf, M.; Stone, L.S. DNA methylation of SPARC and chronic low back pain. Mol. Pain 2011, 7, 65. [Google Scholar] [CrossRef]
- Hou, Y.; Shi, G.; Guo, Y.; Shi, J. Epigenetic modulation of macrophage polarization prevents lumbar disc degeneration. Aging 2020, 12, 6558–6569. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Zhang, Y. DNA methylation in mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a019133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Rao, A.; Ko, M. TET family dioxygenases and DNA demethylation in stem cells and cancers. Exp. Mol. Med. 2017, 49, e323. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.R.; Beyer, C.E.; Stahl, S.M. TRPV1 Antagonists and Chronic Pain: Beyond Thermal Perception. Pharmaceuticals 2012, 5, 114–132. [Google Scholar] [CrossRef]
- Lv, F.J.; Peng, Y.; Lim, F.L.; Sun, Y.; Lv, M.; Zhou, L.; Wang, H.; Zheng, Z.; Cheung, K.M.C.; Leung, V.Y.L. Matrix metalloproteinase 12 is an indicator of intervertebral disc degeneration co-expressed with fibrotic markers. Osteoarthr. Cartil. 2016, 24, 1826–1836. [Google Scholar] [CrossRef] [Green Version]
- Masuda, K.; Aota, Y.; Muehleman, C.; Imai, Y.; Okuma, M.; Thonar, E.J.; Andersson, G.B.; An, H.S. A novel rabbit model of mild, reproducible disc degeneration by an anulus needle puncture: Correlation between the degree of disc injury and radiological and histological appearances of disc degeneration. Spine 2005, 30, 5–14. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.B.; Hwang, E.S.; Kim, E.S.; Kim, S.S.; Jeon, T.D.; Song, M.C.; Lee, J.S.; Chung, M.C.; Maeng, S.; et al. Antidepressant-like Effects of p-Coumaric Acid on LPS-induced Depressive and Inflammatory Changes in Rats. Exp. Neurobiol. 2018, 27, 189–199. [Google Scholar] [CrossRef]
- Fukuta, S.; Miyamoto, K.; Suzuki, K.; Maehara, H.; Inoue, T.; Hara, A.; Kikuike, K.; Taguchi, A.; Shimizu, K. Abundance of calpain and aggrecan-cleavage products of calpain in degenerated human intervertebral discs. Osteoarthr. Cartil. 2011, 19, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Hong, J.Y.; Lee, J.; Jeon, W.J.; Ha, I.H. IL-1beta promotes disc degeneration and inflammation through direct injection of intervertebral disc in a rat lumbar disc herniation model. Spine J. 2021, 21, 1031–1041. [Google Scholar] [CrossRef]
- Choi, S.I.; Lim, J.Y.; Yoo, S.; Kim, H.; Hwang, S.W. Emerging Role of Spinal Cord TRPV1 in Pain Exacerbation. Neural Plast. 2016, 2016, 5954890. [Google Scholar] [CrossRef]
- Sadowska, A.; Hitzl, W.; Karol, A.; Jaszczuk, P.; Cherif, H.; Haglund, L.; Hausmann, O.N.; Wuertz-Kozak, K. Differential regulation of TRP channel gene and protein expression by intervertebral disc degeneration and back pain. Sci. Rep. 2019, 9, 18889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.C.; Wang, K.Y.; Shen, C.K. DNA 5-methylcytosine demethylation activities of the mammalian DNA methyltransferases. J. Biol. Chem. 2013, 288, 9084–9091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, P.; Saadat, A. Neurodegeneration and epigenetics: A review. Neurologia, 2021; in press. [Google Scholar] [CrossRef]
- Buckwalter, J.A. Aging and degeneration of the human intervertebral disc. Spine 1995, 20, 1307–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crofford, L.J. Chronic Pain: Where the Body Meets the Brain. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 167–183. [Google Scholar]
- Bali, K.K.; Kuner, R. Noncoding RNAs: Key molecules in understanding and treating pain. Trends Mol. Med. 2014, 20, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Kissin, I. The development of new analgesics over the past 50 years: A lack of real breakthrough drugs. Anesth. Analg. 2010, 110, 780–789. [Google Scholar] [CrossRef]
- Riccio, A. Dynamic epigenetic regulation in neurons: Enzymes, stimuli and signaling pathways. Nat. Neurosci. 2010, 13, 1330–1337. [Google Scholar] [CrossRef]
- Su, R.C.; Becker, A.B.; Kozyrskyj, A.L.; Hayglass, K.T. Epigenetic regulation of established human type 1 versus type 2 cytokine responses. J. Allergy Clin. Immunol. 2008, 121, 57–63.e53. [Google Scholar] [CrossRef]
- Urban, J.P.; Roberts, S. Degeneration of the intervertebral disc. Arthritis Res. Ther. 2003, 5, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Pagiatakis, C.; Musolino, E.; Gornati, R.; Bernardini, G.; Papait, R. Epigenetics of aging and disease: A brief overview. Aging Clin. Exp. Res. 2021, 33, 737–745. [Google Scholar] [CrossRef]
- Gruber, H.E.; Ingram, J.A.; Leslie, K.; Hanley, E.N., Jr. Cellular, but not matrix, immunolocalization of SPARC in the human intervertebral disc: Decreasing localization with aging and disc degeneration. Spine 2004, 29, 2223–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroke, E.N.; Overstreet, D.S.; Penn, T.M.; Crossman, D.K.; Jackson, P.; Tollefsbol, T.O.; Quinn, T.L.; Yi, N.; Goodin, B.R. Identification of DNA methylation associated enrichment pathways in adults with non-specific chronic low back pain. Mol. Pain 2020, 16, 1744806920972889. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Fu, Y.; Yan, B.; Shen, Z.; Lan, T. Analysis of key genes and pathways associated with the pathogenesis of intervertebral disc degeneration. J. Orthop. Surg. Res. 2020, 15, 371. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kong, J.; Xi, Y.M.; Yu, T.; Wu, X.L.; Hu, Y.G. Effects of AAV2-mediated co-transfection of CTGF and TIMP1 genes on degenerative lumbar intervertebral discs in rhesus monkeys in vivo. Am. J. Transl. Res. 2018, 10, 1085–1096. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-3′ | Primer Sequence |
|---|---|---|
| Aggrecan | Forward | GCCTCTCAAGCCCTTGTCTG |
| Reverse | GATCTCACACAGGTCCCCTC | |
| MMP3 | Forward | ATGATGAACGATGGACAGATGA |
| Reverse | CATTGGCTGAGTGAAAGAGACC | |
| TrpV1 | Forward | TTCACCGAATGGGCCTATGG |
| Reverse | TCACTGCTGCTGTAAGCGAT | |
| DNMT1 | Forward | GTGTGCGGGAATGTGCTCGCT |
| Reverse | CAGTGGTGGTGGCACAGCGT | |
| DNMT3a | Forward | AGCAAAGTGAGGACCATTACCACCA |
| Reverse | TGTGTAGTGGACAGGGAAGCCA | |
| DNMT3b | Forward | TGGCAAGGATGACGTTCTGTGGT |
| Reverse | CTGGCACACTCCAGGACCTTCC | |
| GAPDH | Forward | CCCCCAATGTATCCGTTGTG |
| Reverse | TAGCCCAGGATGCCCTTTAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.Y.; Kim, H.; Jeon, W.-J.; Lee, J.; Yeo, C.; Lee, Y.J.; Ha, I.-H. Epigenetic Changes within the Annulus Fibrosus by DNA Methylation in Rat Intervertebral Disc Degeneration Model. Cells 2022, 11, 3547. https://doi.org/10.3390/cells11223547
Hong JY, Kim H, Jeon W-J, Lee J, Yeo C, Lee YJ, Ha I-H. Epigenetic Changes within the Annulus Fibrosus by DNA Methylation in Rat Intervertebral Disc Degeneration Model. Cells. 2022; 11(22):3547. https://doi.org/10.3390/cells11223547
Chicago/Turabian StyleHong, Jin Young, Hyunseong Kim, Wan-Jin Jeon, Junseon Lee, Changhwan Yeo, Yoon Jae Lee, and In-Hyuk Ha. 2022. "Epigenetic Changes within the Annulus Fibrosus by DNA Methylation in Rat Intervertebral Disc Degeneration Model" Cells 11, no. 22: 3547. https://doi.org/10.3390/cells11223547