

ATP7B-Deficient Hepatocytes Reveal the Importance of Protein Misfolding Induced at Low Copper Concentration
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of ATP7B Knock-Out HepG2/C3a Cell Lines
2.2. Cell Culture
2.3. Immunofluorescence
2.4. Western Blot
2.5. RNA Extraction and Quantification
2.6. FXR Activity
2.7. ICP-MS
2.8. Statistical Analysis
3. Results
3.1. Generation of ATP7B-Deficient Cell Lines Deriving from the HepG2/C3a Cell Line
3.2. Atox1 Expression
3.3. Sensitivity to Cu and Effect of Cu Chelator
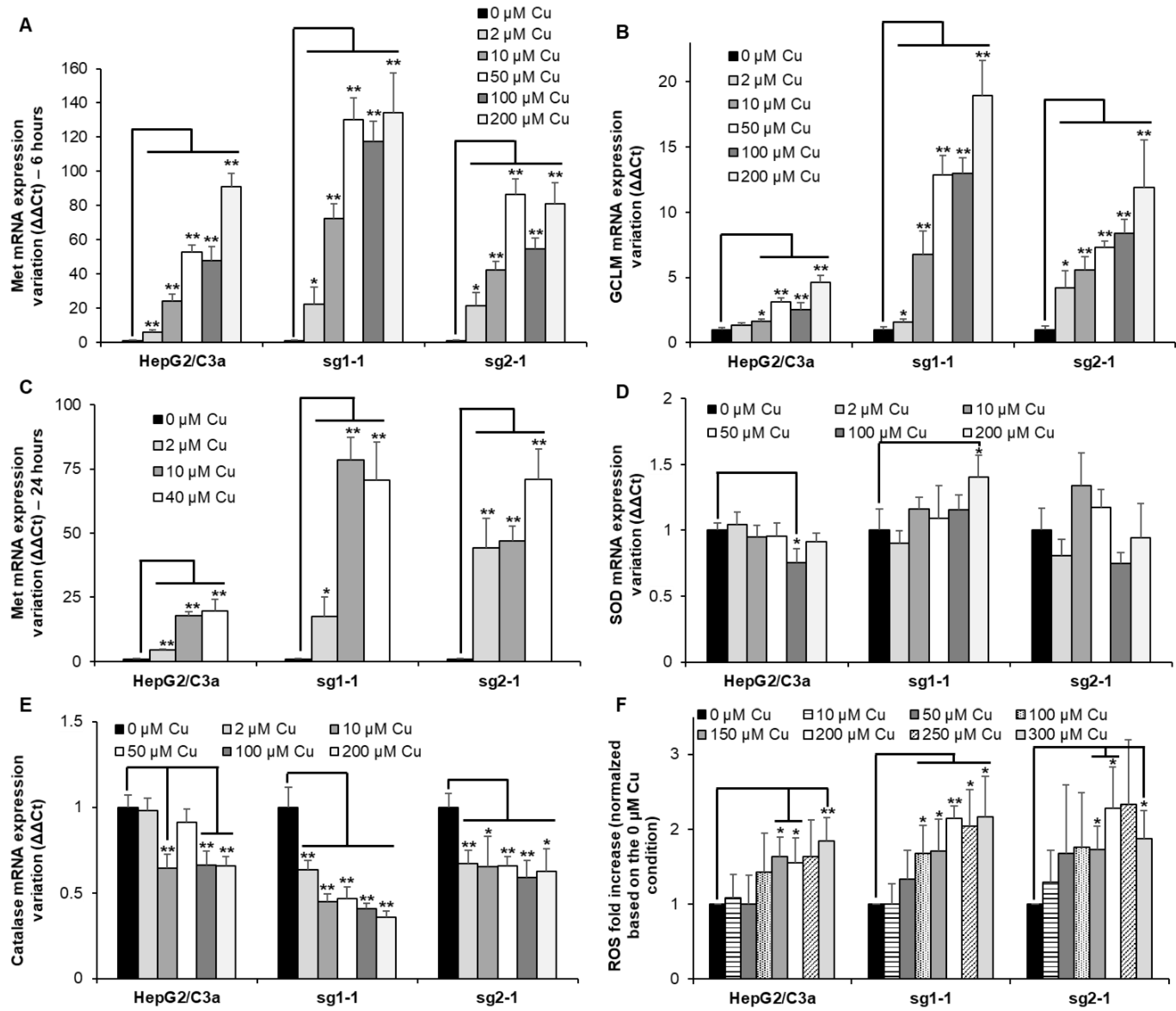
3.4. Metal Stress Response Started at Low Cu Concentration in Cell Line with ATP7B
3.5. Redox Stress Is not Central upon Cu Exposure
3.6. Protein Misfolding upon Cu Exposure Is Exacerbated in ATP7B-Deficient Cell Lines
3.7. Autophagy Is not Activated upon Cu Exposure in the Different Hepatocyte Cell Lines
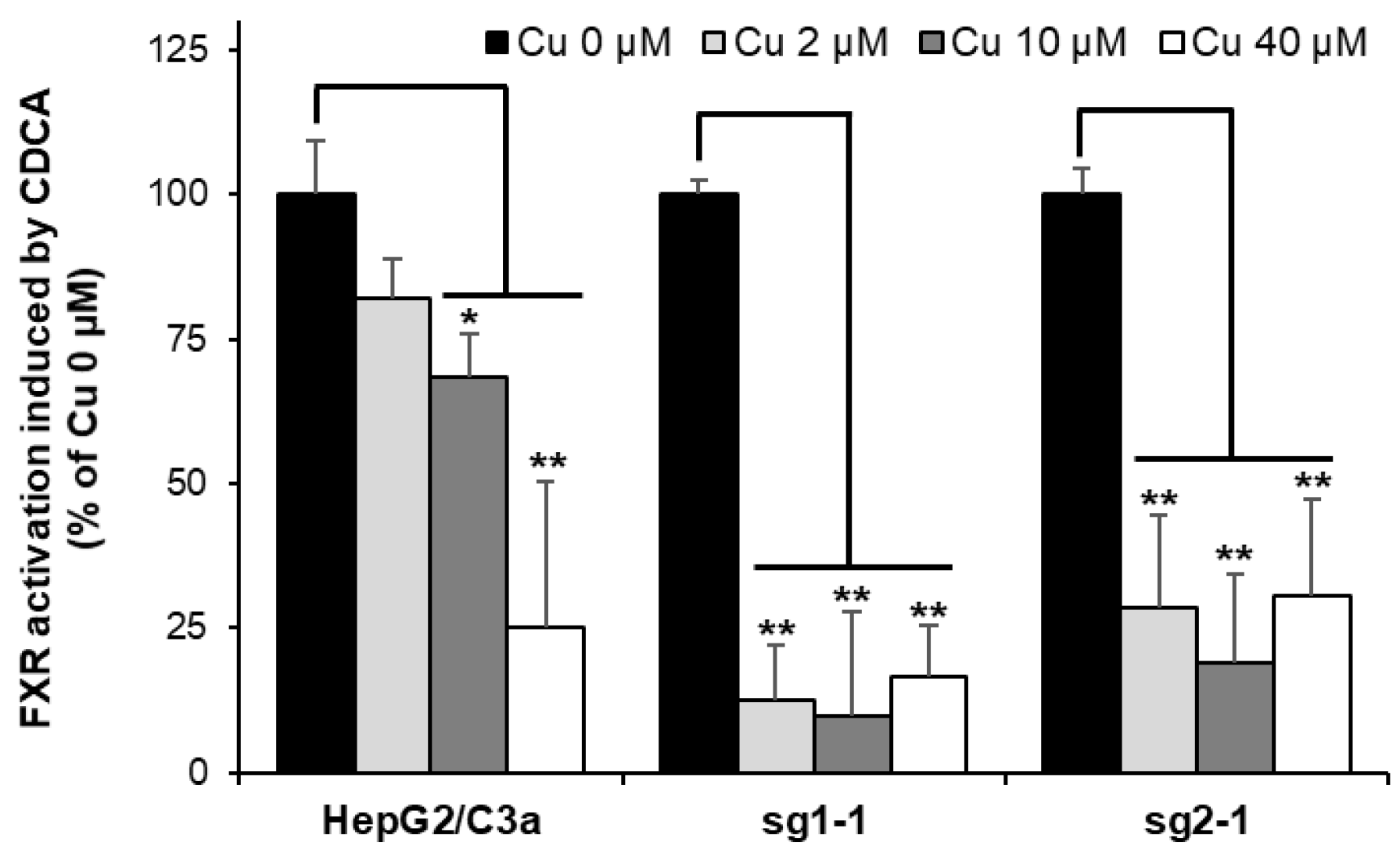
3.8. Nuclear Receptor Activity Is Dramatically Impaired in the Absence of ATP7B upon Cu Exposure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polishchuk, E.V.; Concilli, M.; Iacobacci, S.; Chesi, G.; Pastore, N.; Piccolo, P.; Paladino, S.; Baldantoni, D.; van IJzendoorn, S.C.D.; Chan, J.; et al. Wilson Disease Protein ATP7B Utilizes Lysosomal Exocytosis to Maintain Copper Homeostasis. Dev. Cell 2014, 29, 686–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, K.; Ralle, M.; Schaffer, T.; Jayakanthan, S.; Bari, B.; Muchenditsi, A.; Lutsenko, S. ATP7A and ATP7B Copper Transporters Have Distinct Functions in the Regulation of Neuronal Dopamine-β-Hydroxylase. J. Biol. Chem. 2018, 293, 20085–20098. [Google Scholar] [CrossRef] [Green Version]
- Huster, D.; Kühne, A.; Bhattacharjee, A.; Raines, L.; Jantsch, V.; Noe, J.; Schirrmeister, W.; Sommerer, I.; Sabri, O.; Berr, F.; et al. Diverse Functional Properties of Wilson Disease ATP7B Variants. Gastroenterology 2012, 142, 947–956.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Członkowska, A.; Litwin, T.; Dusek, P.; Ferenci, P.; Lutsenko, S.; Medici, V.; Rybakowski, J.K.; Weiss, K.H.; Schilsky, M.L. Wilson Disease. Nat. Rev. Dis. Primer 2018, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, S. Modifying Factors and Phenotypic Diversity in Wilson’s Disease. Ann. N. Y. Acad. Sci. 2014, 1315, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.P.; Koganti, L.; Muchenditsi, A.; Pendyala, V.S.; Huso, D.; Hankin, J.; Murphy, R.C.; Huster, D.; Merle, U.; Mangels, C.; et al. Activation of Liver X Receptor/Retinoid X Receptor Pathway Ameliorates Liver Disease in Atp7B(-/-) (Wilson Disease) Mice. Hepatology 2016, 63, 1828–1841. [Google Scholar] [CrossRef] [Green Version]
- Wilmarth, P.A.; Short, K.K.; Fiehn, O.; Lutsenko, S.; David, L.L.; Burkhead, J.L. A Systems Approach Implicates Nuclear Receptor Targeting in the Atp7b(-/-) Mouse Model of Wilson’s Disease. Met. Integr. Biometal Sci. 2012, 4, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Wooton-Kee, C.R.; Jain, A.K.; Wagner, M.; Grusak, M.A.; Finegold, M.J.; Lutsenko, S.; Moore, D.D. Elevated Copper Impairs Hepatic Nuclear Receptor Function in Wilson’s Disease. J. Clin. Investig. 2015, 125, 3449–3460. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, A.; Dev, S.; DeVine, L.; Gabrielson, K.L.; Cole, R.N.; Hamilton, J.P.; Lutsenko, S. Hepatic Steatosis in the Mouse Model of Wilson Disease Coincides with a Muted Inflammatory Response. Am. J. Pathol. 2022, 192, 146–159. [Google Scholar] [CrossRef]
- Kłysz, B.; Bembenek, J.; Skowrońska, M.; Członkowska, A.; Kurkowska-Jastrzębska, I. Autonomic Nervous System Dysfunction in Wilson’s Disease—A Systematic Literature Review. Auton. Neurosci. 2021, 236, 102890. [Google Scholar] [CrossRef]
- Weiss, K.H.; Askari, F.K.; Czlonkowska, A.; Ferenci, P.; Bronstein, J.M.; Bega, D.; Ala, A.; Nicholl, D.; Flint, S.; Olsson, L.; et al. Bis-Choline Tetrathiomolybdate in Patients with Wilson’s Disease: An Open-Label, Multicentre, Phase 2 Study. Lancet Gastroenterol. Hepatol. 2017, 2, 869–876. [Google Scholar] [CrossRef]
- Medici, V.; Huster, D. Animal Models of Wilson Disease. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 142, pp. 57–70. ISBN 978-0-444-63625-6. [Google Scholar]
- Sharma, V.R.; Shrivastava, A.; Gallet, B.; Karepina, E.; Charbonnier, P.; Chevallet, M.; Jouneau, P.-H.; Deniaud, A. Canalicular Domain Structure and Function in Matrix-Free Hepatic Spheroids. Biomater. Sci. 2020, 8, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Rekik, Y.; Tardillo Suárez, V.; Sharma, V.R.; Chevallet, M.; Gallet, B.; Falconet, D.; Charbonnier, P.; Kieffer, I.; Tucoulou, R.; Jouneau, P.-H.; et al. Deciphering Silver Nanoparticle Fate in Liver up to Biliary Excretion Using Spheroids in Scenarios Mimicking Different Exposure Pathways. ACS Nano, 2022; in revision. [Google Scholar]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelsen, T.S.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-Scale CRISPR-Cas9 Knockout Screening in Human Cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjana, N.E.; Shalem, O.; Zhang, F. Improved Vectors and Genome-Wide Libraries for CRISPR Screening. Nat. Methods 2014, 11, 783–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deniaud, A.; Karuppasamy, M.; Bock, T.; Masiulis, S.; Huard, K.; Garzoni, F.; Kerschgens, K.; Hentze, M.W.; Kulozik, A.E.; Beck, M.; et al. A Network of SMG-8, SMG-9 and SMG-1 C-Terminal Insertion Domain Regulates UPF1 Substrate Recruitment and Phosphorylation. Nucleic Acids Res. 2015, 43, 7600–7611. [Google Scholar] [CrossRef] [Green Version]
- Veronesi, G.; Deniaud, A.; Gallon, T.; Jouneau, P.-H.; Villanova, J.; Delangle, P.; Carrière, M.; Kieffer, I.; Charbonnier, P.; Mintz, E.; et al. Visualization, Quantification and Coordination of Ag+ Ions Released from Silver Nanoparticles in Hepatocytes. Nanoscale 2016, 8, 17012–17021. [Google Scholar] [CrossRef] [Green Version]
- Fey, S.J.; Wrzesinski, K. Determination of Drug Toxicity Using 3D Spheroids Constructed From an Immortal Human Hepatocyte Cell Line. Toxicol. Sci. 2012, 127, 403–411. [Google Scholar] [CrossRef]
- Walker, J.M.; Tsivkovskii, R.; Lutsenko, S. Metallochaperone Atox1 Transfers Copper to the NH2-Terminal Domain of the Wilson’s Disease Protein and Regulates Its Catalytic Activity. J. Biol. Chem. 2002, 277, 27953–27959. [Google Scholar] [CrossRef] [Green Version]
- Hamza, I.; Prohaska, J.; Gitlin, J.D. Essential Role for Atox1 in the Copper-Mediated Intracellular Trafficking of the Menkes ATPase. Proc. Natl. Acad. Sci. USA 2003, 100, 1215–1220. [Google Scholar] [CrossRef] [Green Version]
- Clifford, R.J.; Maryon, E.B.; Kaplan, J.H. Dynamic Internalization and Recycling of a Metal Ion Transporter: Cu Homeostasis and HCTR1, the Human Cu Uptake System. J. Cell Sci. 2016, 129, 1711–1721. [Google Scholar] [CrossRef]
- Petris, M.J.; Smith, K.; Lee, J.; Thiele, D.J. Copper-Stimulated Endocytosis and Degradation of the Human Copper Transporter, HCtr1. J. Biol. Chem. 2003, 278, 9639–9646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, D.E.K.; Stillman, M.J. The “Magic Numbers” of Metallothionein. Metallomics 2011, 3, 444. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Glutathione Synthesis. Biochim. Biophys. Acta BBA-Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, S.N.; Hausen, J.; Nachev, M.; Ottermanns, R.; Schiwy, S.; Hollert, H. Short Exposure to Cadmium Disrupts the Olfactory System of Zebrafish (Danio Rerio)—Relating Altered Gene Expression in the Olfactory Organ to Behavioral Deficits. Aquat. Toxicol. 2020, 226, 105555. [Google Scholar] [CrossRef]
- Cuillel, M.; Chevallet, M.; Charbonnier, P.; Fauquant, C.; Pignot-Paintrand, I.; Arnaud, J.; Cassio, D.; Michaud-Soret, I.; Mintz, E. Interference of CuO Nanoparticles with Metal Homeostasis in Hepatocytes under Sub-Toxic Conditions. Nanoscale 2014, 6, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Alam, J.; Cook, J.L. How Many Transcription Factors Does It Take to Turn On the Heme Oxygenase-1 Gene? Am. J. Respir. Cell Mol. Biol. 2007, 36, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Saporito-Magriñá, C.M.; Musacco-Sebio, R.N.; Andrieux, G.; Kook, L.; Orrego, M.T.; Tuttolomondo, M.V.; Desimone, M.F.; Boerries, M.; Borner, C.; Repetto, M.G. Copper-Induced Cell Death and the Protective Role of Glutathione: The Implication of Impaired Protein Folding Rather than Oxidative Stress. Metallomics 2018, 10, 1743–1754. [Google Scholar] [CrossRef]
- Shimberg, G.D.; Ok, K.; Neu, H.M.; Splan, K.E.; Michel, S.L.J. Cu(I) Disrupts the Structure and Function of the Nonclassical Zinc Finger Protein Tristetraprolin (TTP). Inorg. Chem. 2017, 56, 6838–6848. [Google Scholar] [CrossRef] [Green Version]
- Polishchuk, E.V.; Merolla, A.; Lichtmannegger, J.; Romano, A.; Indrieri, A.; Ilyechova, E.Y.; Concilli, M.; De Cegli, R.; Crispino, R.; Mariniello, M.; et al. Activation of Autophagy, Observed in Liver Tissues From Patients With Wilson Disease and From ATP7B-Deficient Animals, Protects Hepatocytes From Copper-Induced Apoptosis. Gastroenterology 2019, 156, 1173–1189.e5. [Google Scholar] [CrossRef]
- Tardillo Suárez, V.; Karepina, E.; Chevallet, M.; Gallet, B.; Cottet-Rousselle, C.; Charbonnier, P.; Moriscot, C.; Michaud-Soret, I.; Bal, W.; Fuchs, A.; et al. Nuclear Translocation of Silver Ions and Hepatocyte Nuclear Receptor Impairment upon Exposure to Silver Nanoparticles. Environ. Sci. Nano 2020, 7, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Hageman, J.; van Waarde, M.A.W.H.; Zylicz, A.; Walerych, D.; Kampinga, H.H. The Diverse Members of the Mammalian HSP70 Machine Show Distinct Chaperone-like Activities. Biochem. J. 2011, 435, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, E.J.; Place, R.F.; Giardina, C.; Hightower, L.E. Hsp70B′ Regulation and Function. Cell Stress Chaperones 2007, 12, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, E.; Giardina, C.; Hightower, L. Hsp70B′ and Hsp72 Form a Complex in Stressed Human Colon Cells and Each Contributes to Cytoprotection☆. Exp. Cell Res. 2008, 314, 2468–2476. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charbonnier, P.; Chovelon, B.; Ravelet, C.; Ngo, T.D.; Chevallet, M.; Deniaud, A. ATP7B-Deficient Hepatocytes Reveal the Importance of Protein Misfolding Induced at Low Copper Concentration. Cells 2022, 11, 3400. https://doi.org/10.3390/cells11213400
Charbonnier P, Chovelon B, Ravelet C, Ngo TD, Chevallet M, Deniaud A. ATP7B-Deficient Hepatocytes Reveal the Importance of Protein Misfolding Induced at Low Copper Concentration. Cells. 2022; 11(21):3400. https://doi.org/10.3390/cells11213400
Chicago/Turabian StyleCharbonnier, Peggy, Benoît Chovelon, Corinne Ravelet, Tuan Dung Ngo, Mireille Chevallet, and Aurélien Deniaud. 2022. "ATP7B-Deficient Hepatocytes Reveal the Importance of Protein Misfolding Induced at Low Copper Concentration" Cells 11, no. 21: 3400. https://doi.org/10.3390/cells11213400