
Towards the Standardization of Intestinal In Vitro Advanced Barrier Model for Nanoparticles Uptake and Crossing: The SiO2 Case Study

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Bi-coltures and Tri-Culture Model Assembly on Inserts
- Caco-2 monoculture on inserts was obtained by seeding 2.25 × 103 cells on the Ap compartment of Polyethylene Terephthalate Transwells® insert (PET) with a pore size of 1 and 3 μm (Millipore®) (Merck KGaA, Darmstadt, Germany), allocated in 12-well culture plates (Falcon) with 0.5 mL of complete culture medium in the Ap compartment, while Bl compartment was filled with 1.5 mL. DMEM complete culture medium was changed every two days.
- Caco-2/HT29-MTX bi-cultures were obtained by mixing and seeding the two cell lines at 9:1 ratio at a density of 2.25 × 105 cells/cm2, in the Ap compartment. Inserts were then maintained as reported for Caco-2 monoculture.
- Caco-/Raji and Caco-HT29-MTX/Raji B co-cultures were obtained by adding at the 14th day of co-culture, 5 × 105 Raji B cells in the Bl side of each insert in DMEM:RPMI (1:1) culture medium to allow M phenotype induction in Caco-2 cells. Raji B cells were completely removed and reseeded on day 16th and 19th of co-culture.
- Raji B cells seeded in the Bl compartment. Raji B cells were seeded in the Bl compartment and refreshed at 14th, 16th, and 19th days of culture to prevent their overgrowth which might cause degeneration of the lymphocyte’s population and, consequently, presence of necrotic factors in the model.
- Simplified inverted culture method. In this case Caco-2 and Caco-2 /HT29-MTX cells were seeded on the Bl side of the insert and left for 48 h to allow cells’ adherence to the insert membrane before turning them upside down. Inverted cells were cultured for 14 days and then stimulated with Raji B cells placed in the Ap compartment. The inverted model required an elevated level of expertise coupled with a high risk of contamination during the turning phase of the inserts. For this reason, it was considered not suitable for the next steps in procedure standardization and will not be further discussed.
- Raji B conditioned medium. To investigate if the direct contact between intestinal epithelial cells and lymphocytes is required to induce M-phenotype in Caco-2 cells, conditioned medium from Raji B cells was also used at each cycle of induction (14, 16 and 19 days of co-culture). Lymphocytes conditioned medium from Raj B cells was obtained as follows: the day before each induction, 330 × 103 Raji B cells/mL Raji B cells were cultured in DMEM/RPMI (1:1) for 24 h, then the cellular suspension was centrifuged and 1.5 mL of the supernatant was added to the Bl compartment of the insert. This experiment has been investigated only with 3 µm pore size inserts to maximize the effect of diffusion factor(s).
2.3. Barrier Integrity
2.4. Paracellular Permeability
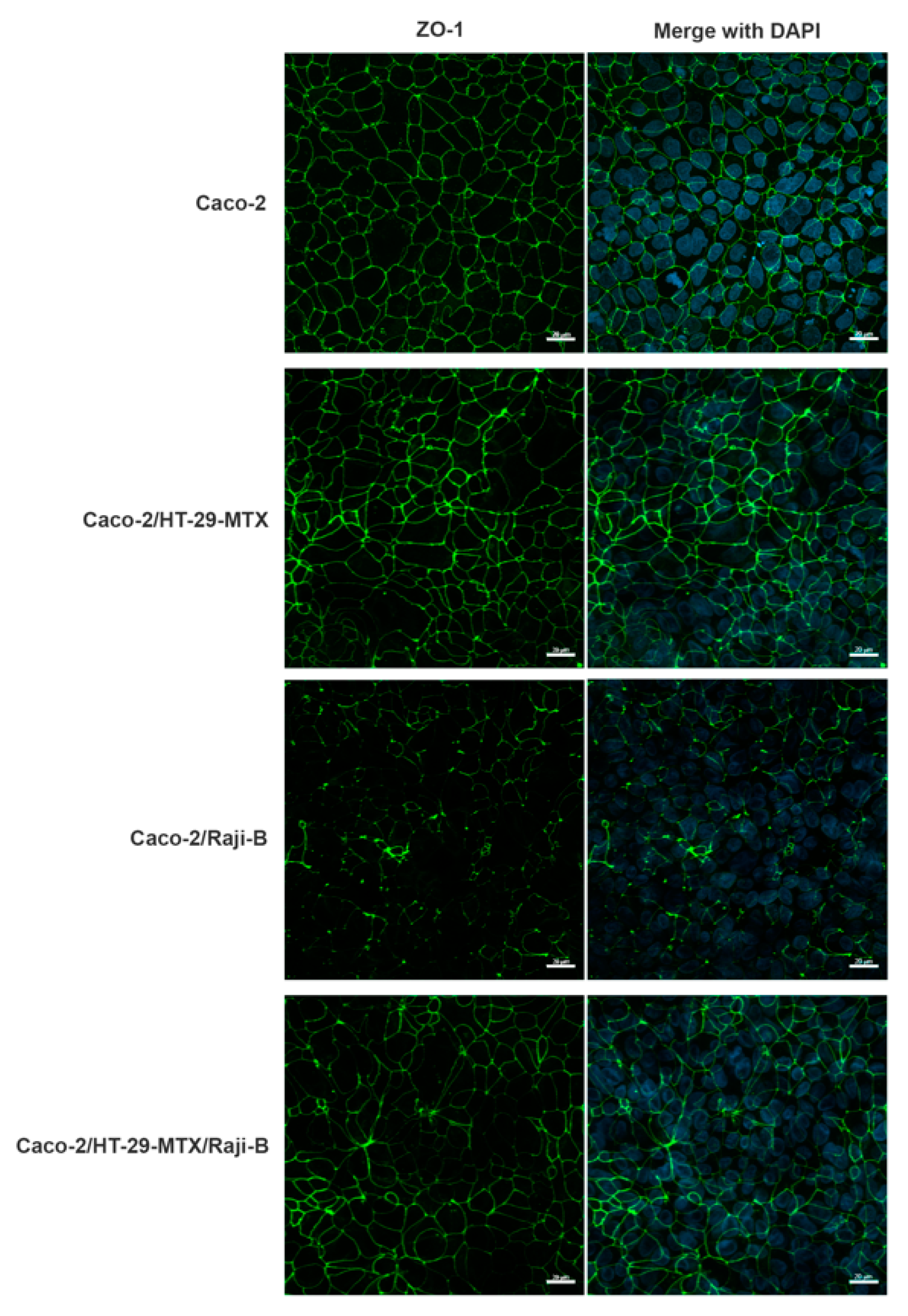
2.5. Localization of Tight Junction ZO-1 Protein
2.6. Mucus Characterization
2.6.1. Mucus Staining
2.6.2. Muc-5AC and Muc-2 Expression and Release
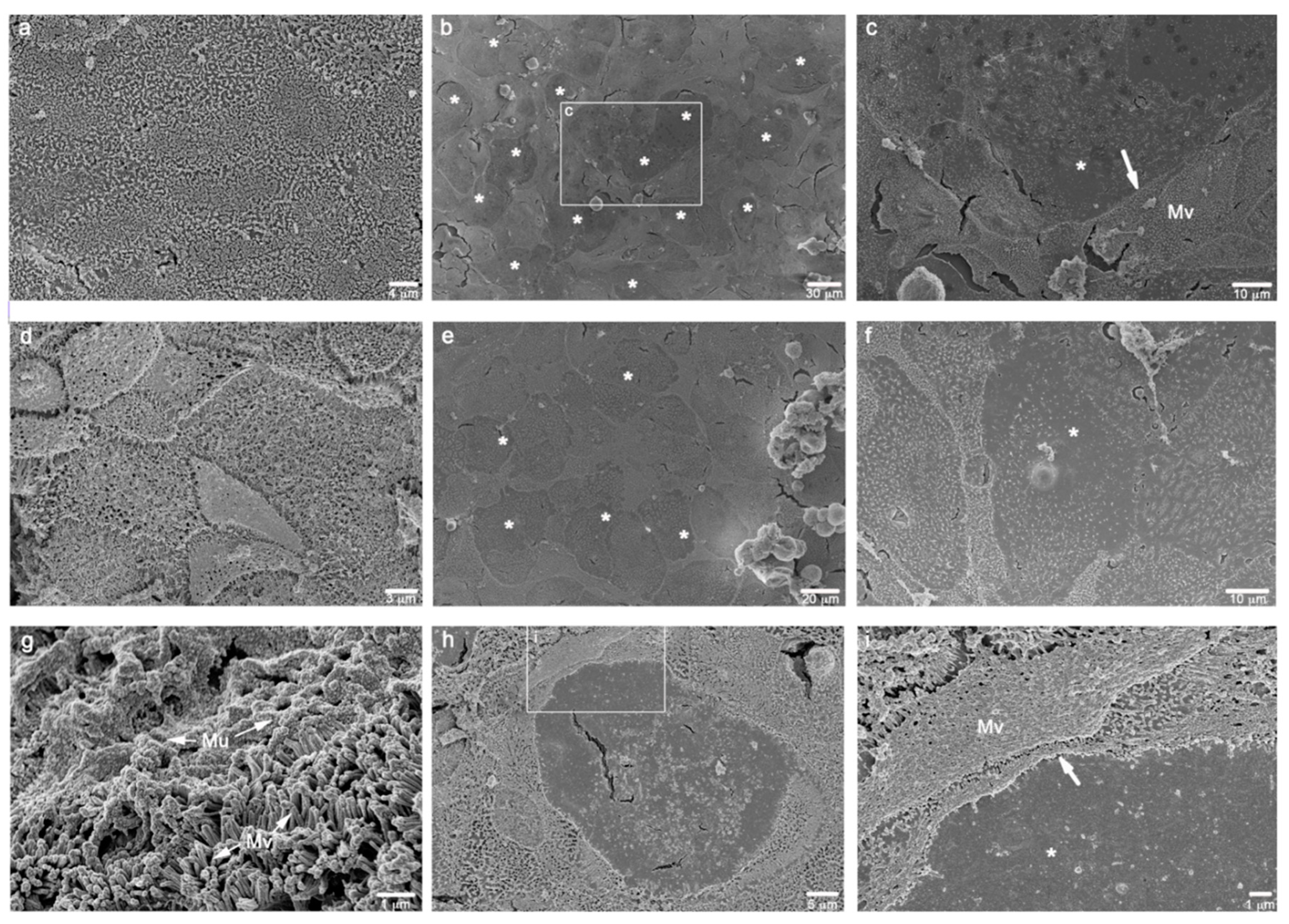
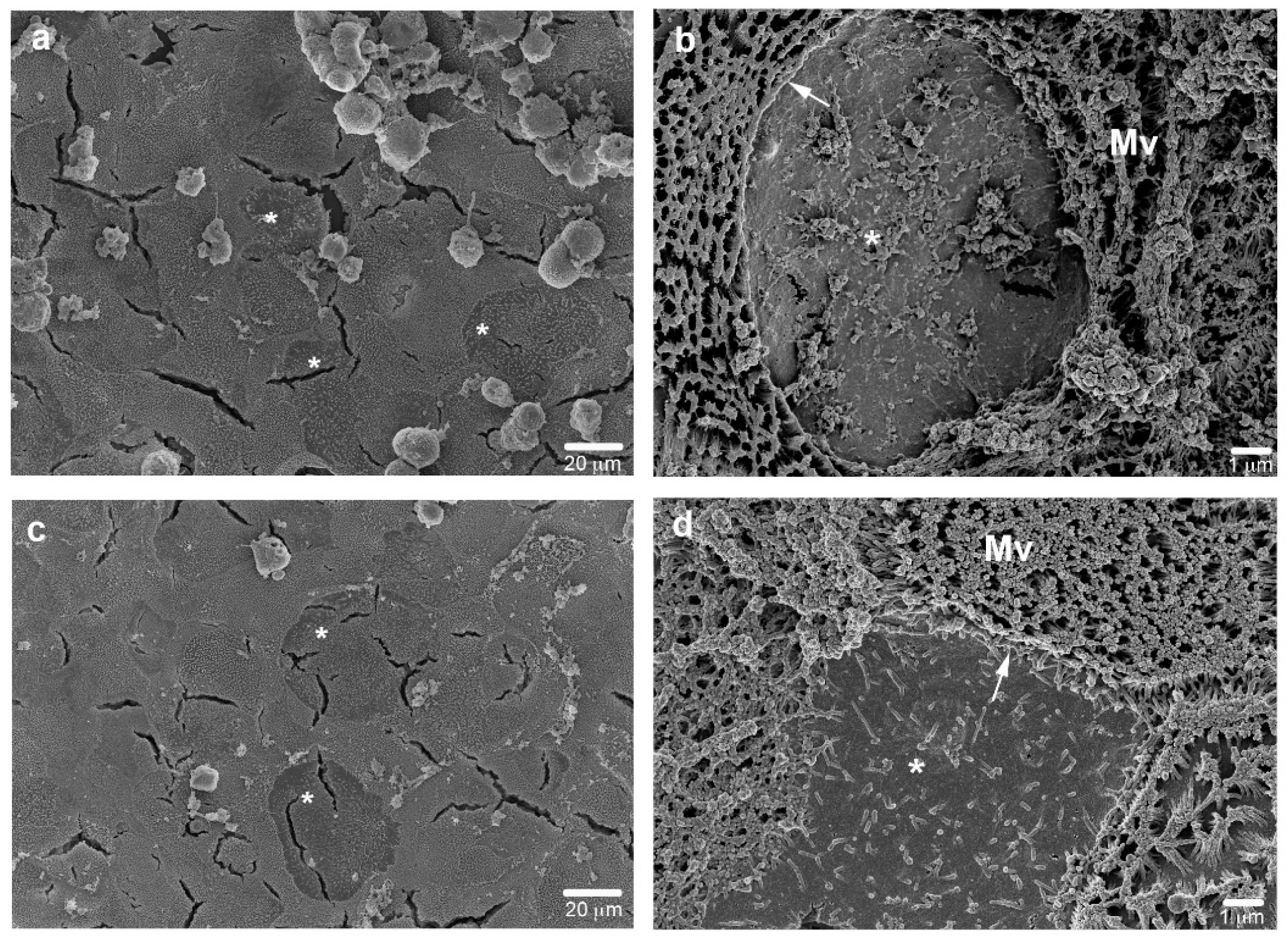
2.7. Electron Microscopy
2.8. SiO2 NPs Dispersion Preparation and Characterization
2.9. SiO2 NPs Translocation and Uptake
2.10. Statistical Analysis
3. Results
3.1. M Cell Phenotype Induction
3.1.1. Raji B Exposure Conditions
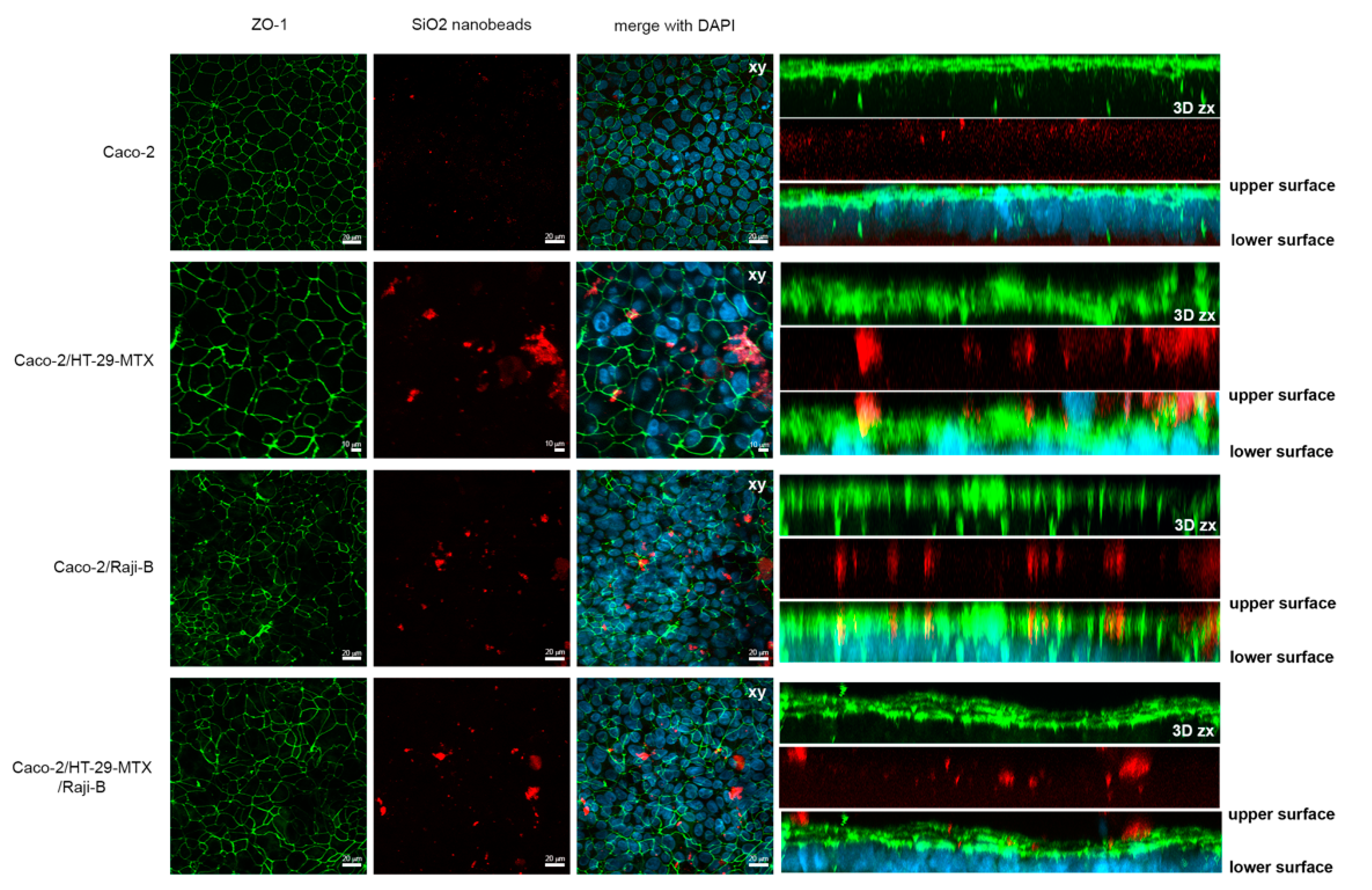
3.1.2. M-Cell Marker: Zonula Occludens (ZO-1) Expression
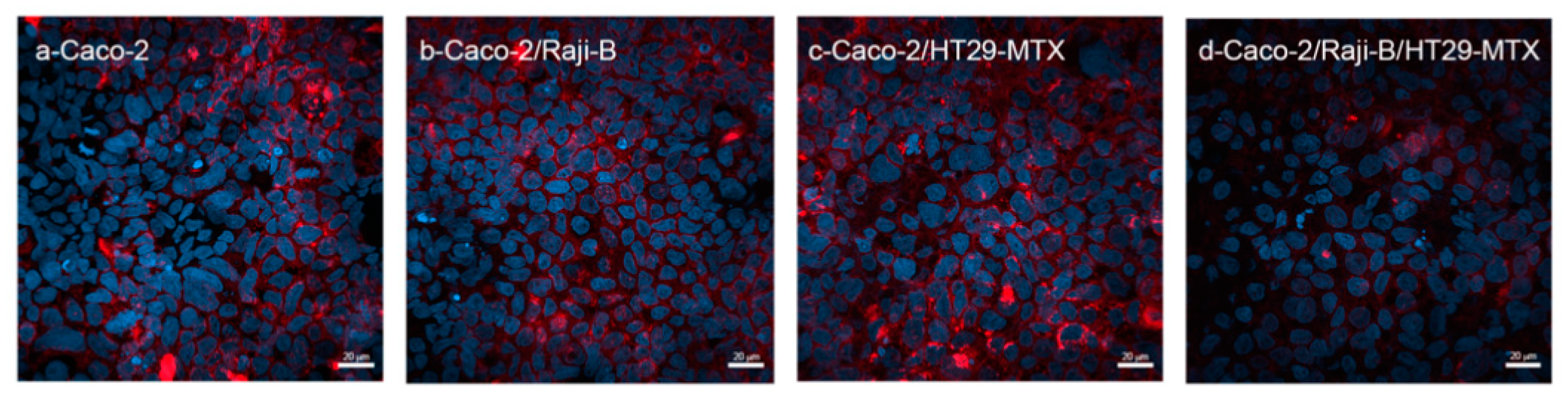
3.1.3. M Cell Marker: Wheat Germ Agglutinin (WGA)
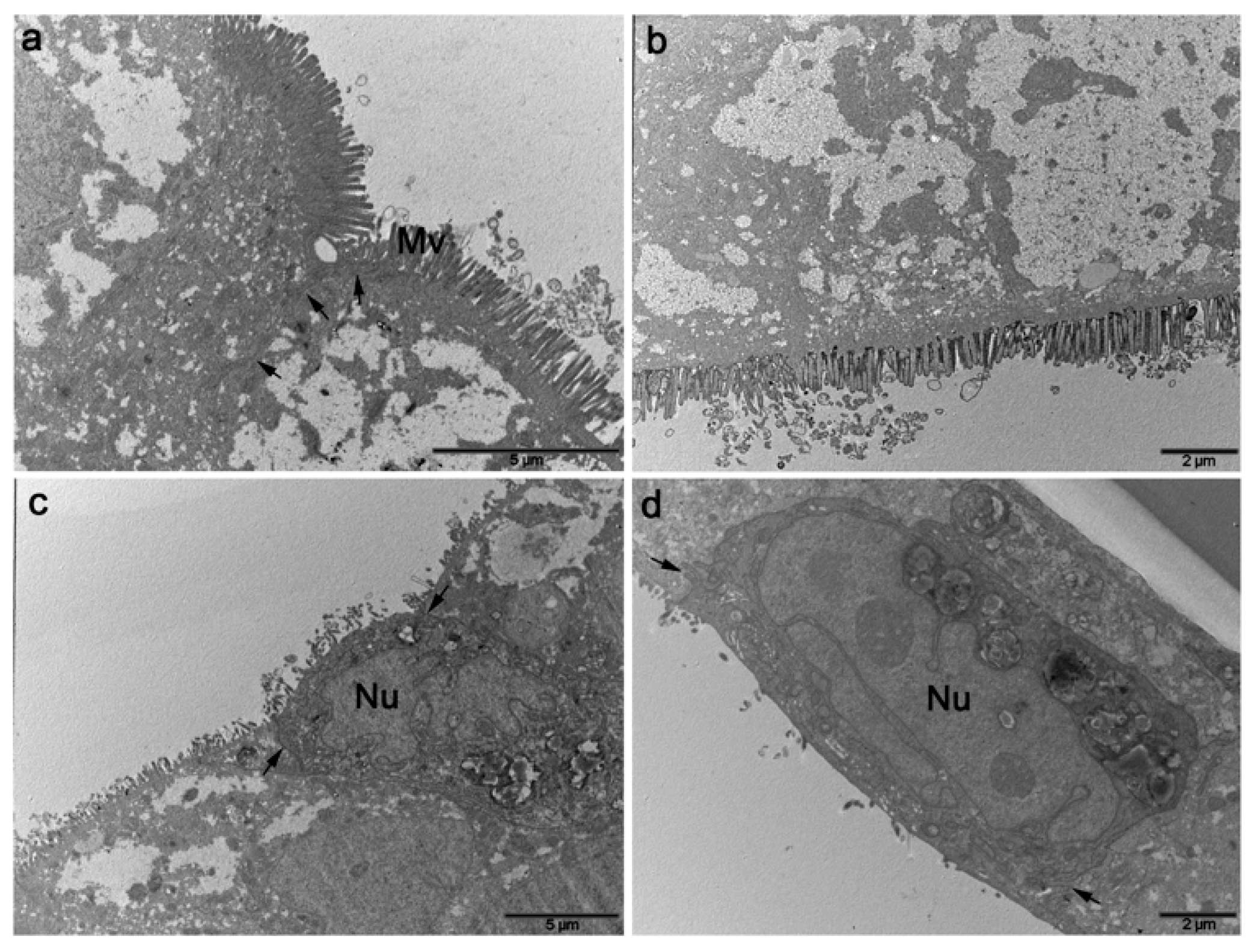
3.1.4. Ultrastructural Characterization
3.2. Barrier Integrity
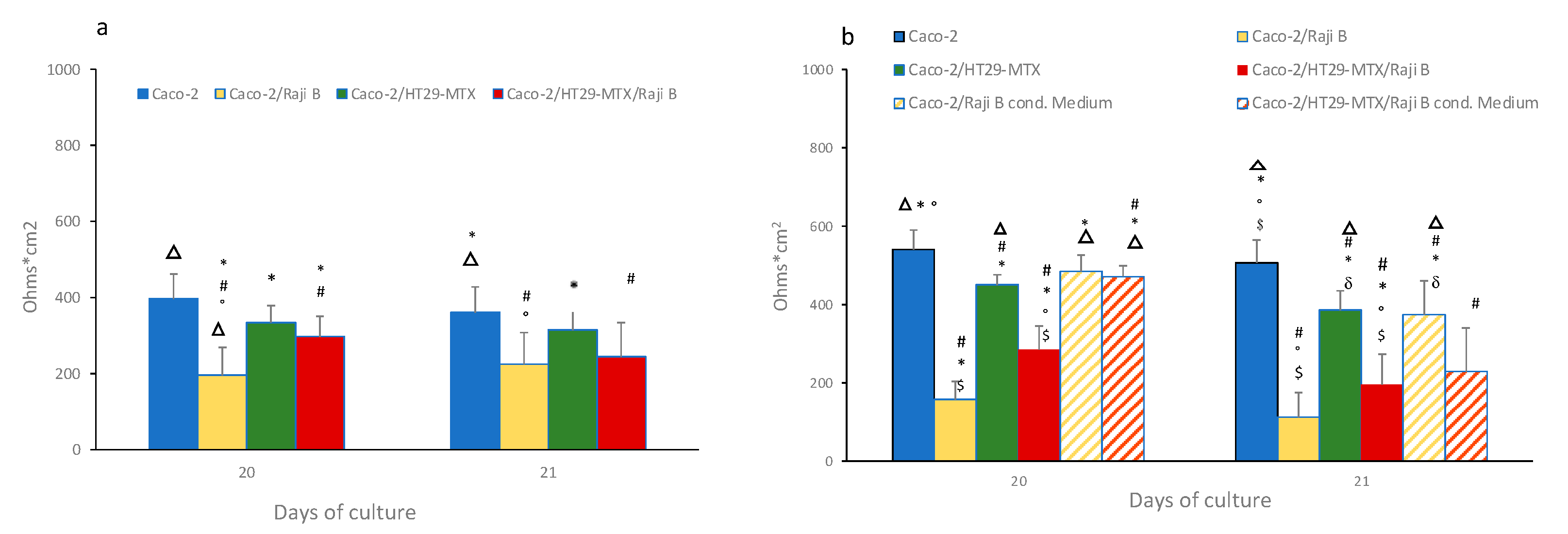
3.2.1. Transepithelial Electrical Resistance (TEER)
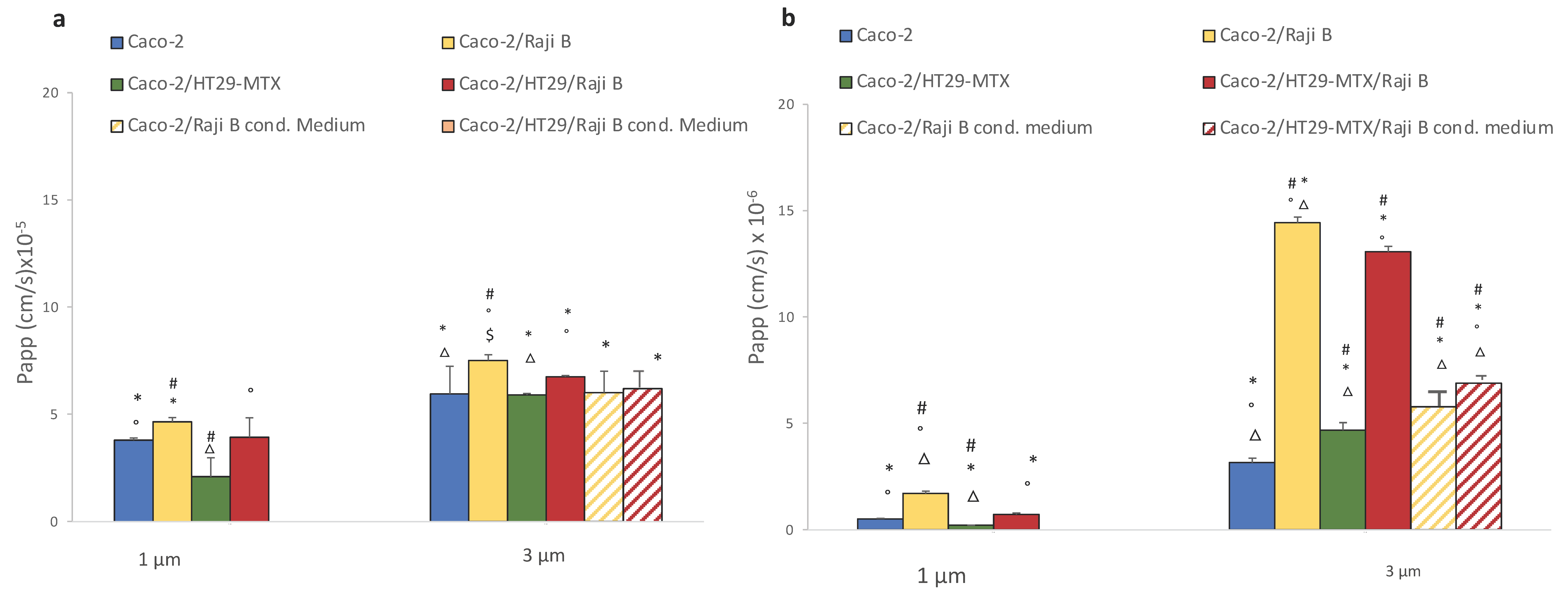
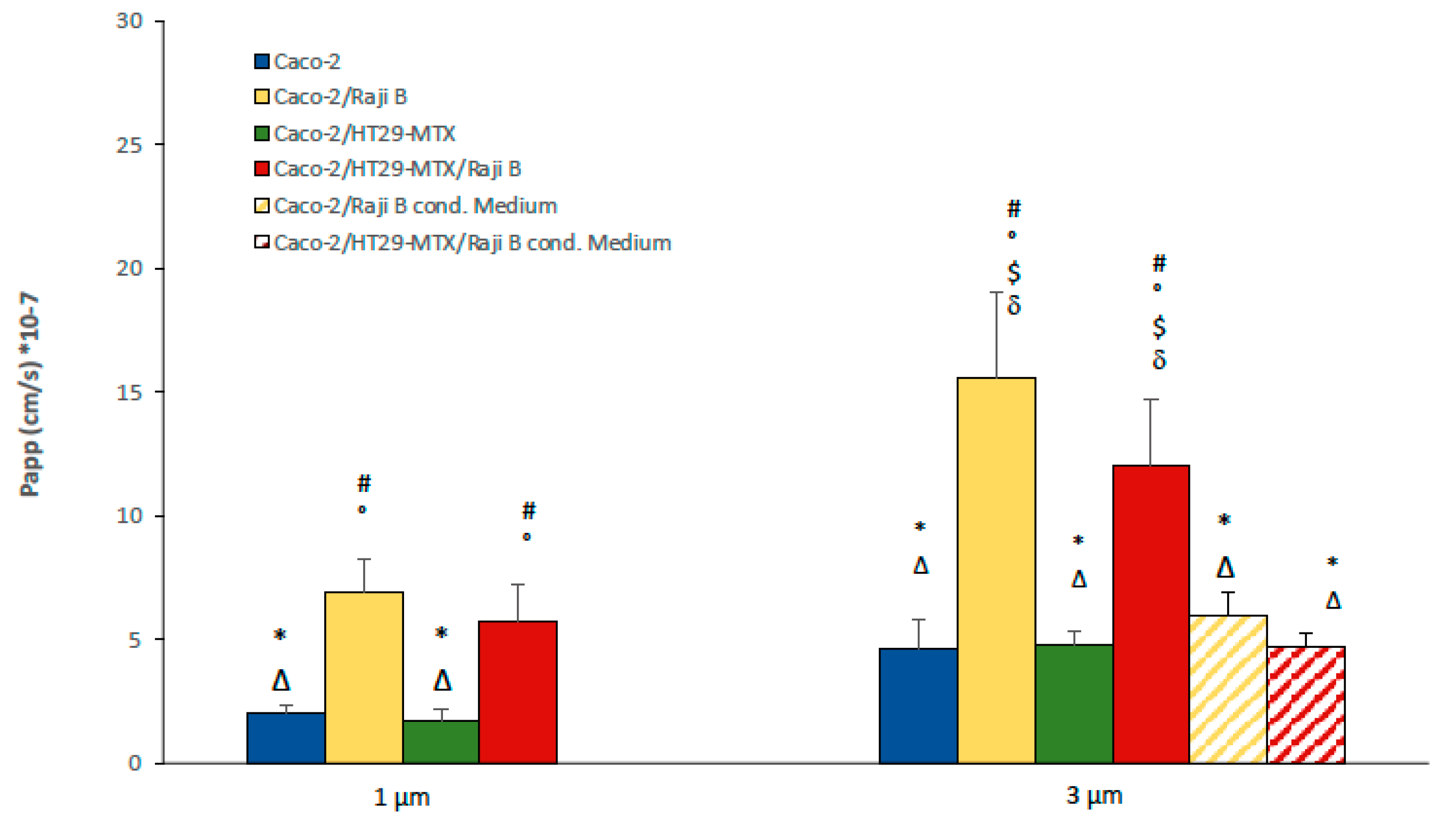
3.2.2. FITC-Dextran and Lucifer Yellow Passage
3.3. Mucus Characterization
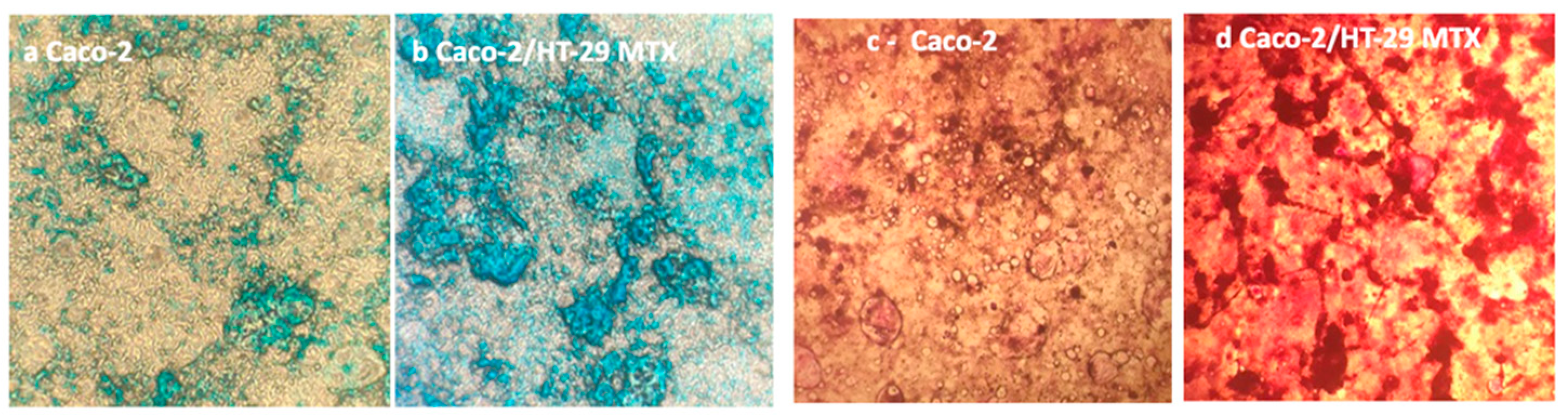
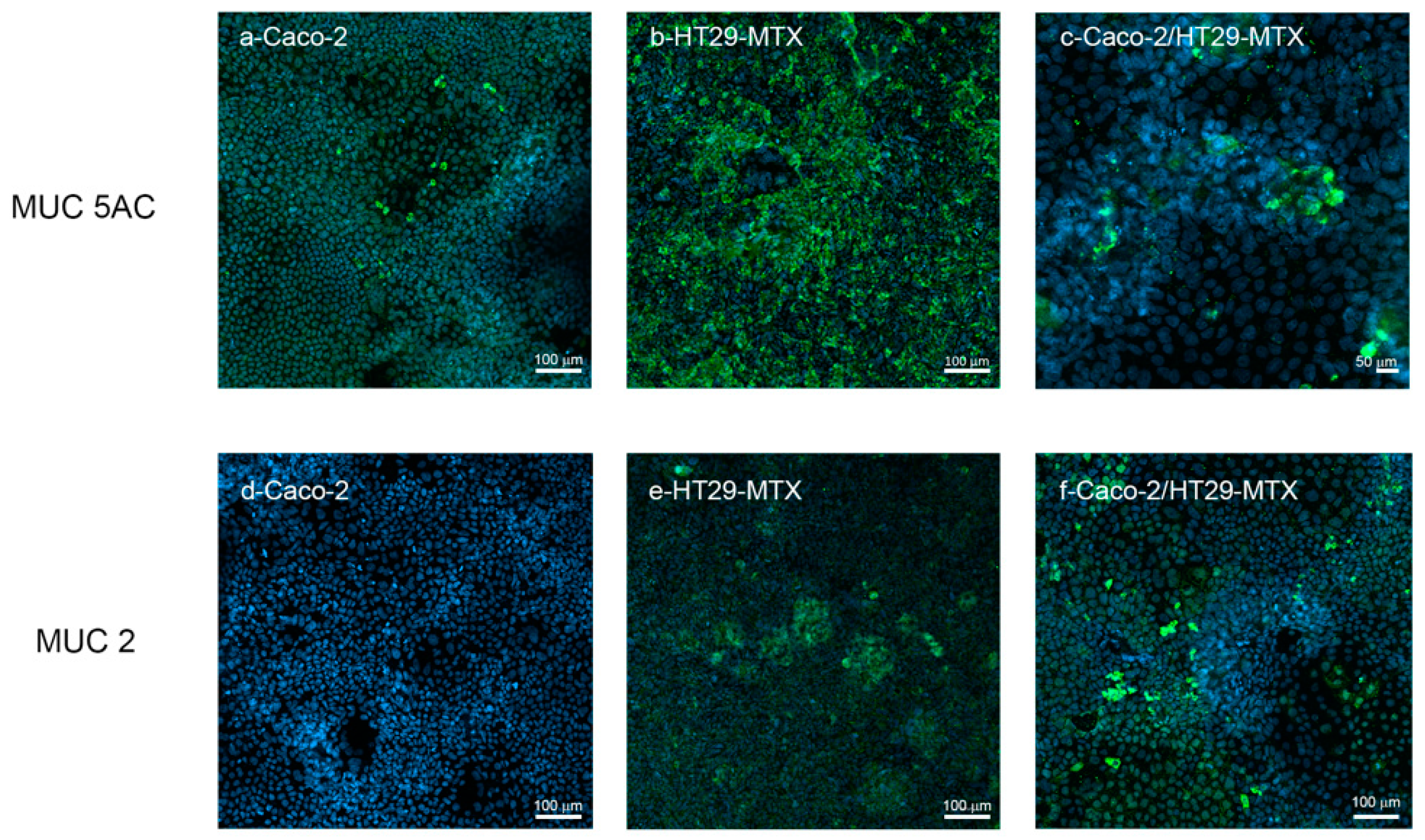
3.3.1. Mucus Staining
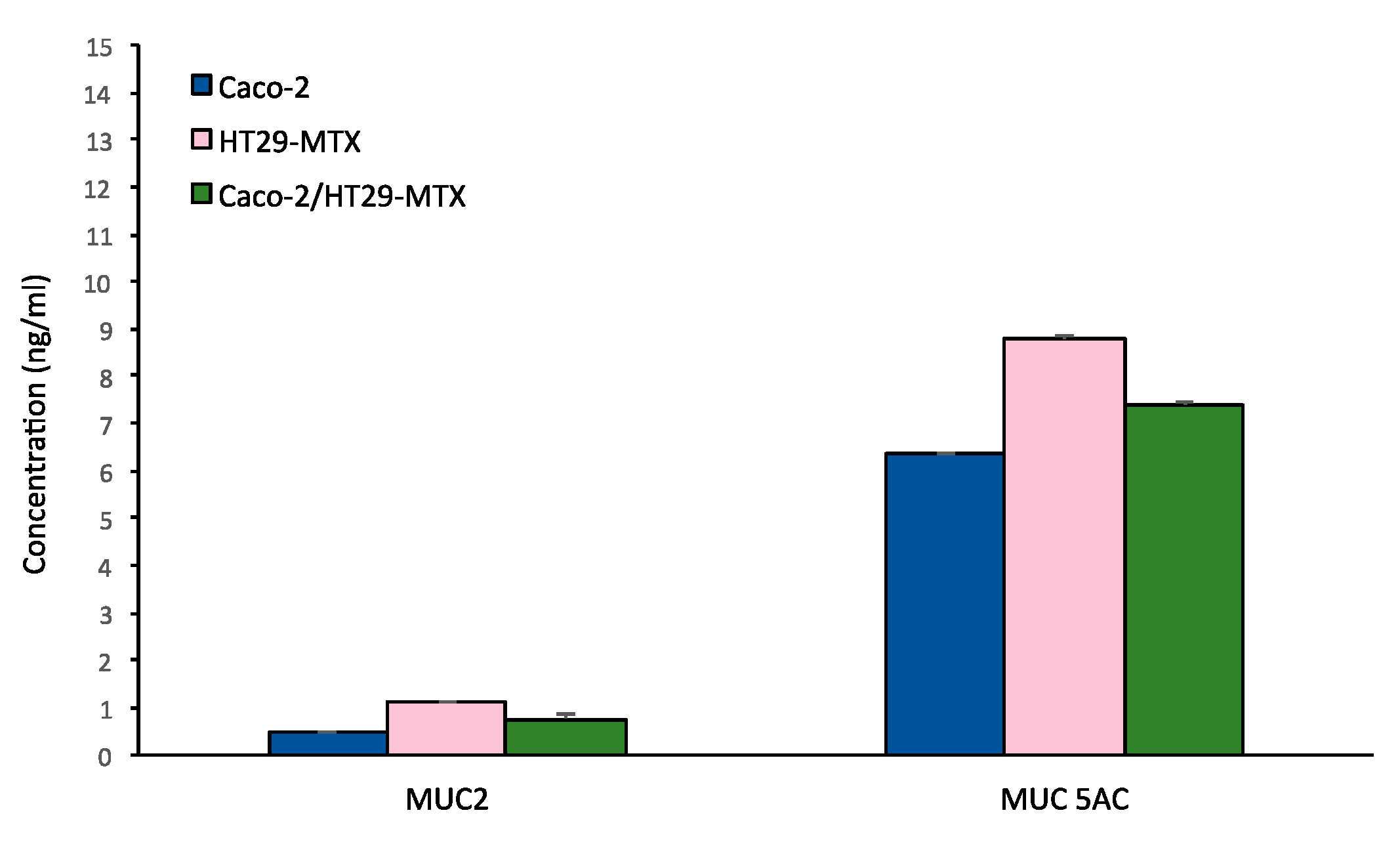
3.3.2. MuC-2 and MuC5 AC Expression and Release
3.4. SiO2 NPs Uptake and Translocation as a Case Study for Tri-Culture Model Efficiency
3.4.1. SIO2 NPs Characterization by DLS
3.4.2. Uptake and Translocation
4. Discussion
5. Conclusions
- M cell phenotype induction: the best protocol is to co-culture Caco-2/HT29 –MTX at 9:1 ratio and, from the 14th day of culture, to add Raji B cells for the following 5 days in the Bl compartment in RPMI/DMEM (1:1) medium, refreshing them for three times. Inserts are put to rest for the next 24 h before any experimental activity. Model is stable for about 48 h;
- M cell marker: no specific marker for M cells has been identified. Electron microscopy (SEM or TEM) is now the unique way to identify them but further investigations are required to find a more practical and cheaper approach;
- Determination of barrier integrity is an important parameter of the model, easy and fast to measure. Both TEER and LY are simple and reproducible indicators of this parameter and, based on present data and literature evidence, the following acceptance criteria are proposed: TEER values must not be below 150 ohms × cm2 and the LY Papp value must not be greater than 15 × 10−6.
- Insert pore size is important aspect for NPs translocation experiments; present data indicate that 3 µm pore dimension is preferable;
- Co-culture of Caco-2/HT29 –MTX at 9:1 ratio produces an abundant mucus layer in which MUC-5AC are predominant. Alcian blue and Periodic Shiff staining can be used as routine method to verify the presence of the mucus layer in the (co)-culture.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD. Test No. 318: Dispersion Stability of Nanomaterials in Simulated Environmental Media. In OECD Guidelines for the Testing of Chemicals, Section 3; OECD Publishing: Paris, France, 2017. [Google Scholar] [CrossRef]
- Fadeel, B.; Farcal, L.; Hardy, B.; Vázquez-Campos, S.; Hristozov, D.; Marcomini, A.; Lynch, I.; Valsami-Jones, E.; Alenius, H.; Savolainen, K. Advanced tools for the safety assessment of nanomaterials. Nat. Nanotechnol. 2018, 13, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Clift, M.J.D.; Jenkins, G.J.S.; Doak, S.H. An Alternative Perspective towards Reducing the Risk of Engineered Nanomaterials to Human Health. Small 2020, 16, e2002002. [Google Scholar] [CrossRef] [PubMed]
- Guggenheim, E.J.; Milani, S.; Röttgermann, P.J.; Dusinska, M.; Saout, C.; Salvati, A.; Rädler, J.O.; Lynch, I. Refining in vitro models for nanomaterial exposure to cells and tissues. NanoImpact 2018, 10, 121–142. [Google Scholar] [CrossRef]
- Romeo, D.; Salieri, B.; Hischier, R.; Nowack, B.; Wick, P. An integrated pathway based on in vitro data for the human hazard assessment of nanomaterials. Environ. Int. 2020, 137, 105505. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J.F. Toxic Heavy Metal and Metalloid Accumulation in Crop Plants and Foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef] [Green Version]
- McCracken, C.; Dutta, P.K.; Waldman, W.J. Critical assessment of toxicological effects of ingested nano-particles. Environ. Sci. Nano 2016, 3, 256–282. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Zhao, Z.; Ng, K.W. Transformation of Nanomaterials and Its Implications in Gut Nanotoxicology. Small 2020, 16, e2001246. [Google Scholar] [CrossRef]
- Pietroiusti, A.; Bergamaschi, E.; Campagna, M.; Campagnolo, L.; De Palma, G.; Iavicoli, S.; Leso, V.; Magrini, A.; Miragoli, M.; Pedata, P.; et al. The unrecognized occupational relevance of the interaction between engineered nanomaterials and the gastro-intestinal tract: A consensus paper from a multidisciplinary working group. Part. Fibre Toxicol. 2017, 14, 1–23. [Google Scholar] [CrossRef]
- Lefebvre, D.E.; Venema, K.; Gombau, L.; Jr, L.G.V.; Raju, J.; Bondy, G.S.; Bouwmeester, H.; Singh, R.P.; Clippinger, A.J.; Collnot, E.-M.; et al. Utility of models of the gastrointestinal tract for assessment of the digestion and absorption of engineered nanomaterials released from food matrices. Nanotoxicology 2014, 9, 523–542. [Google Scholar] [CrossRef]
- Sohal, I.S.; O’Fallon, K.S.; Gaines, P.; Demokritou, P.; Bello, D. Ingested engineered nanomaterials: State of science in nanotoxicity testing and future research needs. Part. Fibre Toxicol. 2018, 15, 1–31. [Google Scholar] [CrossRef] [Green Version]
- EFSA Scientific Committee; Hardy, A.; Benford, D.; Halldorsson, T.; Jeger, M.J.; Knutsen, H.K.; More, S.; Naegeli, H.; Noteborn, H.; Ockleford, C.; et al. Guidance on risk assessment of the application of nanoscience and nanotechnologies in the food and feed chain: Part 1, human and animal health. EFSA J. 2018, 16, e05327. [Google Scholar] [CrossRef] [Green Version]
- ISO/TR 18401:2017; Nanotechnologies-Plain language explanation of selected terms from the ISO/IEC 80004 series. ISO publishing: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/62384.html (accessed on 18 August 2022).
- Gerloff, K.; Pereira, D.; Faria, N.; Boots, A.W.; Kolling, J.; Förster, I.; Albrecht, C.; Powell, J.J.; Schins, R.P. Influence of simulated gastrointestinal conditions on particle-induced cytotoxicity and interleukin-8 regulation in differentiated and undifferentiated Caco-2 cells. Nanotoxicology 2012, 7, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bove, P.; Malvindi, M.A.; Kote, S.S.; Bertorelli, R.; Summa, M.; Sabella, S. Dissolution test for risk assessment of nanoparticles: A pilot study. Nanoscale 2017, 9, 6315–6326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Carnovale, C.; Guarnieri, D.; Di Cristo, L.; De Angelis, I.; Veronesi, G.; Scarpellini, A.; Malvindi, M.; Barone, F.; Pompa, P.; Sabella, S. Biotransformation of Silver Nanoparticles into Oro-Gastrointestinal Tract by Integrated In Vitro Testing Assay: Generation of Exposure-Dependent Physical Descriptors for Nanomaterial Grouping. Nanomater. 2021, 11, 1587. [Google Scholar] [CrossRef]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Béduneau, A.; Tempesta, C.; Fimbel, S.; Pellequer, Y.; Jannin, V.; Demarne, F.; Lamprecht, A. A tunable Caco-2/HT29-MTX co-culture model mimicking variable permeabilities of the human intestine obtained by an original seeding procedure. Eur. J. Pharm. Biopharm. 2014, 87, 290–298. [Google Scholar] [CrossRef]
- Hilgendorf, C.; Spahn-Langguth, H.; Regårdh, C.G.; Lipka, E.; Amidon, G.L.; Langguth, P. Caco-2 versus Caco-2/HT29-MTX Co-cultured Cell Lines: Permeabilities Via Diffusion, Inside- and Outside-Directed Carrier-Mediated Transport. J. Pharm. Sci. 2000, 89, 63–75. [Google Scholar] [CrossRef]
- Araújo, F.; Sarmento, B. Towards the characterization of an in vitro triple co-culture intestine cell model for permeability studies. Int. J. Pharm. 2013, 458, 128–134. [Google Scholar] [CrossRef]
- Schimpel, C.; Teubl, B.; Absenger, M.; Meindl, C.; Fröhlich, E.; Leitinger, G.; Zimmer, A.; Roblegg, E. Development of an Advanced Intestinal in Vitro Triple Culture Permeability Model To Study Transport of Nanoparticles. Mol. Pharm. 2014, 11, 808–818. [Google Scholar] [CrossRef]
- Rubio, L.; Marcos, R.; Hernández, A. Potential adverse health effects of ingested micro- and nanoplastics on humans. Lessons learned from in vivo and in vitro mammalian models. J. Toxicol. Environ. Heal. Part B 2019, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Youhanna, S.; Lauschke, V.M. The Past, Present and Future of Intestinal In Vitro Cell Systems for Drug Absorption Studies. J. Pharm. Sci. 2020, 110, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Beloqui, A.; Brayden, D.J.; Artursson, P.; Préat, V.; Rieux, A.D. A human intestinal M-cell-like model for investigating particle, antigen and microorganism translocation. Nat. Protoc. 2017, 12, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Mabbott, N.A.; Donaldson, D.S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013, 6, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Gullberg, E.; Leonard, M.; Karlsson, J.; Hopkins, A.; Brayden, D.; Baird, A.; Artursson, P. Expression of Specific Markers and Particle Transport in a New Human Intestinal M-Cell Model. Biochem. Biophys. Res. Commun. 2000, 279, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Corr, S.C.; Gahan, C.C.; Hill, C. M-cells: Origin, morphology and role in mucosal immunity and microbial pathogenesis. FEMS Immunol. Med. Microbiol. 2008, 52, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.M.; Walker, S.L.; Lo, D.D. Epithelial Microvilli Establish an Electrostatic Barrier to Microbial Adhesion. Infect. Immun. 2014, 82, 2860–2871. [Google Scholar] [CrossRef] [Green Version]
- Mabbott, N.A.; Sehgal, A.; Donaldson, D.S. M Cells. In eLS; John Wiley & Sons Publisher: Chichester, UK, 2015; pp. 1–9. [Google Scholar] [CrossRef]
- Dillon, A.; Lo, D.D. M Cells: Intelligent Engineering of Mucosal Immune Surveillance. Front. Immunol. 2019, 10, 1499. [Google Scholar] [CrossRef]
- Wood, M.B.; Rios, D.; Williams, I.R. TNF-α augments RANKL-dependent intestinal M cell differentiation in enteroid cultures. Am. J. Physiol. Physiol. 2016, 311, C498–C507. [Google Scholar] [CrossRef] [Green Version]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [Green Version]
- Lock, J.Y.; Carlson, T.L.; Carrier, R.L. Mucus models to evaluate the diffusion of drugs and particles. Adv. Drug Deliv. Rev. 2017, 124, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Porchet, N.; Aubert, J.-P.; Swallow, D.; Gum, J.R.; Real, F.X.; Zweibaum, A. Differential Expression of the Human Mucin Genes MUC1 to MUC5 in Relation to Growth and Differentiation of Different Mucus-Secreting HT-29 Cell Subpopula-tions. J. Cell Sci. 1993, 106, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Bao, L.; Wang, X.; Chen, C. The Nano–Intestine Interaction: Understanding the Location-Oriented Effects of Engineered Nanomaterials in the Intestine. Small 2020, 16, e1907665. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Powell, D.N.; Kalman, D. Layered defense: How mucus and tight junctions seal the intestinal barrier. Klin. Wochenschr. 2017, 95, 927–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozoya-Agullo, I.; Araújo, F.; González-Álvarez, I.; Merino-Sanjuán, M.; González-Álvarez, M.; Bermejo, M.; Sarmento, B. Usefulness of Caco-2/HT29-MTX and Caco-2/HT29-MTX/Raji B Coculture Models to Predict Intestinal and Colonic Permeability Compared to Caco-2 Monoculture. Mol. Pharm. 2017, 14, 1264–1270. [Google Scholar] [CrossRef]
- Lesuffleur, T.; Barbat, A.; Dussaulx, E.; Zwibaum, A. Growth Adaptation to Methotrexate of HT-29 Human Colon Carcinoma Cells Is Associated with Their Ability to Differentiate into Columnar Absorptive and Mucus-secreting Cells. Cancer Res. 1990, 50, 6334–6343. [Google Scholar]
- Rieux, A.D.; Fievez, V.; Théate, I.; Mast, J.; Préat, V.; Schneider, Y.-J. An improved in vitro model of human intestinal follicle-associated epithelium to study nanoparticle transport by M cells. Eur. J. Pharm. Sci. 2007, 30, 380–391. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; Vila, L.; Cortés, C.; Hernández, A.; Marcos, R. Exploring the usefulness of the complex in vitro intestinal epithelial model Caco-2/HT29/Raji-B in nanotoxicology. Food Chem. Toxicol. 2018, 113, 162–170. [Google Scholar] [CrossRef]
- Pereira, C.; Costa, J.; Sarmento, B.; Araújo, F. Cell-based in vitro models for intestinal permeability studies. In Concepts and Models for Drug Permeability Studies; Sarmento, B., Ed.; Woodhead Publisher: Sawston, UK, 2016; pp. 57–81. [Google Scholar] [CrossRef]
- Kämpfer, A.A.; Urbán, P.; La Spina, R.; Jiménez, I.O.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Ongoing inflammation enhances the toxicity of engineered nanomaterials: Application of an in vitro co-culture model of the healthy and inflamed intestine. Toxicol. Vitr. 2019, 63, 104738. [Google Scholar] [CrossRef]
- George, I.; Vranic, S.; Boland, S.; Courtois, A.; Baeza-Squiban, A. Development of an in vitro model of human bronchial epithelial barrier to study nanoparticle translocation. Toxicol. Vitro 2015, 29, 51–58. [Google Scholar] [CrossRef]
- Antunes, F.; Andrade, F.; Araújo, F.; Ferreira, D.; Sarmento, B. Establishment of a triple co-culture in vitro cell models to study intestinal absorption of peptide drugs. Eur. J. Pharm. Biopharm. 2012, 83, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Åberg, C. Quantitative analysis of nanoparticle transport through in vitro blood-brain barrier models. Tissue Barriers 2016, 4, e1143545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitia, L.; Catelani, T.; Guarnieri, D.; Pompa, P.P. In Vitro Blood–Brain Barrier Models for Nanomedicine: Particle-Specific Effects and Methodological Drawbacks. ACS Appl. Bio. Mater. 2019, 2, 3279–3289. [Google Scholar] [CrossRef]
- Bernardo, A.; Malara, M.; Bertuccini, L.; De Nuccio, C.; Visentin, S.; Minghetti, L. The Antihypertensive Drug Telmisartan Protects Oligodendrocytes from Cholesterol Accumulation and Promotes Differentiation by a PPAR-γ-Mediated Mechanism. Int. J. Mol. Sci. 2021, 22, 9434. [Google Scholar] [CrossRef] [PubMed]
- Negroni, A.; Costanzo, M.; Vitali, R.; Superti, F.; Bertuccini, L.; Tinari, A.; Minelli, F.; Di Nardo, G.; Nuti, F.; Pierdomenico, M.; et al. Characterization of adherent-invasive Escherichia coli isolated from pediatric patients with inflammatory bowel disease. Inflamm. Bowel. Dis. 2012, 18, 913–924. [Google Scholar] [CrossRef]
- Pielage, J.F.; Cichon, C.; Greune, L.; Hirashima, M.; Kucharzik, T.; Schmidt, M.A. Reversible differentiation of Caco-2 cells reveals galectin-9 as a surface marker molecule for human follicle-associated epithelia and M cell-like cells. Int. J. Biochem. Cell Biol. 2007, 39, 1886–1901. [Google Scholar] [CrossRef]
- Phillips, T.E.; Frisch, E.B. Secretory glycoconjugates of a mucin-synthesizing human colonic adenocarcinoma cell line. Histochem. Cell Biol. 1990, 93, 311–317. [Google Scholar] [CrossRef]
- Wikman-Larhed, A.; Artursson, P. Co-cultures of human intestinal goblet (HT29-H) and absorptive (Caco-2) cells for studies of drug and peptide absorption. Eur. J. Pharm. Sci. 1995, 3, 171–183. [Google Scholar] [CrossRef]
- Pan, F.; Han, L.; Zhang, Y.; Yu, Y.; Liu, J. Optimization of Caco-2 and HT29 co-culture in vitro cell models for permeability studies. Int. J. Food Sci. Nutr. 2015, 66, 680–685. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; Vila, L.; Cortés, C.; Hernández, A.; Marcos, R. Effects of differently shaped TiO2NPs (nanospheres, nanorods and nanowires) on the in vitro model (Caco-2/HT29) of the intestinal barrier. Part. Fibre. Toxicol. 2018, 15, 1–16. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Ermund, A.; Johansson, M.E.V.; Schütte, A.; Hansson, G.C.; Sjövall, H. An ex vivo method for studying mucus formation, properties, and thickness in human colonic biopsies and mouse small and large intestinal explants. Am. J. Physiol. Liver Physiol. 2012, 302, G430–G438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernéis, S.; Bogdanova, A.; Kraehenbuhl, J.-P.; Pringault, E. Conversion by Peyer’s Patch Lymphocytes of Human Enterocytes into M Cells that Transport Bacteria. Science 1997, 277, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Savidge, T.C.; Smith, M.W. Evidence that Membranous (M) Cell Genesis is Immuno-Regulated. In Advances in Mucosal Immunology; Springer: New York, NY, USA, 1995; pp. 239–241. [Google Scholar] [CrossRef]
- Honbo, A.; Masaoka, Y.; Kataoka, M.; Sakuma, S.T.; Ragnarsson, E.G.E.; Artursson, P.; Yamashita, S. Development of an in vitro M-cell model for macromolecule transport. In Proceedings of the 19th Annual Meeting of JSSX, Kanazawa, Japan, 17–19 November 2004; Volume 19, p. 18PE-93. [Google Scholar] [CrossRef]
- Jang, M.H.; Kweon, M.-N.; Iwatani, K.; Yamamoto, M.; Terahara, K.; Sasakawa, C.; Suzuki, T.; Nochi, T.; Yokota, Y.; Rennert, P.D.; et al. Intestinal villous M cells: An antigen entry site in the mucosal epithelium. Proc. Natl. Acad. Sci. 2004, 101, 6110–6115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardelli, L.; Pacheco, D.P.; Ziccarelli, A.; Tunesi, M.; Caspani, O.; Fusari, A.; Briatico Vangosa, F.; Giordano, C.; & Petrini, P. Towards bioinspired in vitro models of intestinal mucus. RSC Advances, RSC. Adv. 2019, 9, 15887–15899. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dispersant | Time (Hours) | Z-Ave ± SD (nm) | PDI ± SD |
|---|---|---|---|
| H2O | 0 | 82. ± 2 | 0.09 ± 0.01 |
| DMEM 1%FCS | 0 | 155 ± 2 | 0.22 ± 0.02 |
| 24 | 125.7 ± 2 | 0.29 ± 0.08 |
| Parameters | Indicators | Protocol Optimization |
|---|---|---|
| Transwell support | Insert pore size | 3 µm pore inserts more suitable for NPs translocation studies |
| Barrier integrity | TEER and LY | Yes, acceptance criteria proposed |
| Mucus production | Alcian blue and Periodic Schiff staining | Yes, but only qualitative data |
| M-cell phenotype induction | Biochemical markers, TEM/SEM images | Yes, for TEM/SEM investigation (qualitative data) No specific biochemical markers identified |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vincentini, O.; Prota, V.; Cecchetti, S.; Bertuccini, L.; Tinari, A.; Iosi, F.; De Angelis, I. Towards the Standardization of Intestinal In Vitro Advanced Barrier Model for Nanoparticles Uptake and Crossing: The SiO2 Case Study. Cells 2022, 11, 3357. https://doi.org/10.3390/cells11213357
Vincentini O, Prota V, Cecchetti S, Bertuccini L, Tinari A, Iosi F, De Angelis I. Towards the Standardization of Intestinal In Vitro Advanced Barrier Model for Nanoparticles Uptake and Crossing: The SiO2 Case Study. Cells. 2022; 11(21):3357. https://doi.org/10.3390/cells11213357
Chicago/Turabian StyleVincentini, Olimpia, Valentina Prota, Serena Cecchetti, Lucia Bertuccini, Antonella Tinari, Francesca Iosi, and Isabella De Angelis. 2022. "Towards the Standardization of Intestinal In Vitro Advanced Barrier Model for Nanoparticles Uptake and Crossing: The SiO2 Case Study" Cells 11, no. 21: 3357. https://doi.org/10.3390/cells11213357