
Neutrophil Extracellular Vesicles and Airway Smooth Muscle Proliferation in the Natural Model of Severe Asthma in Horses
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Protocol
2.2. Pulmonary Function Test
2.3. Endoscopy and Collection of Respiratory Samples
2.4. Neutrophil Isolation
2.5. Neutrophil-Derived Extracellular Vesicles Isolation and Characterization
2.5.1. Extracellular Vesicles Isolation
2.5.2. Nanoparticle Tracking Analysis
2.5.3. Proteomics of Neutrophil Extracellular Vesicles
2.6. Airway Smooth Muscle Isolation and Characterization
2.6.1. Cell Culture
2.6.2. Flow Cytometry
2.6.3. Airway Smooth Muscle Cells’ Real-Time Proliferation Assay
2.7. Statistical Analysis
3. Results
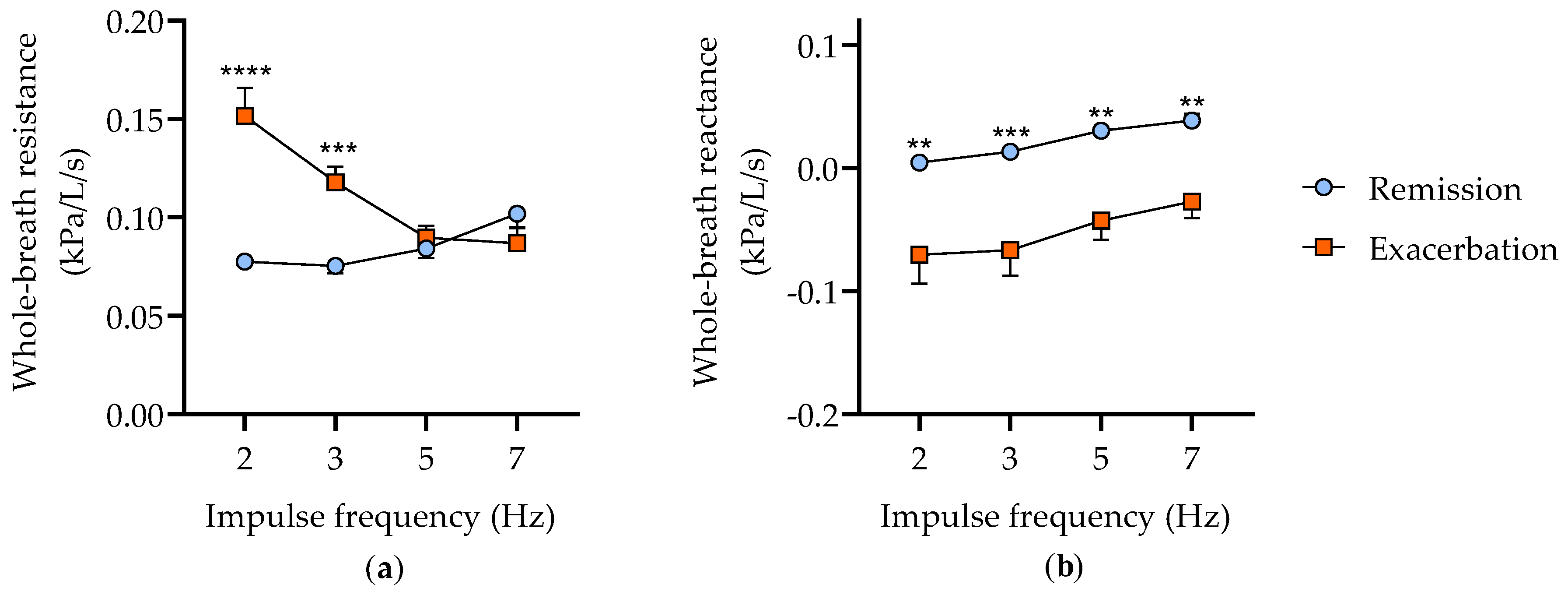
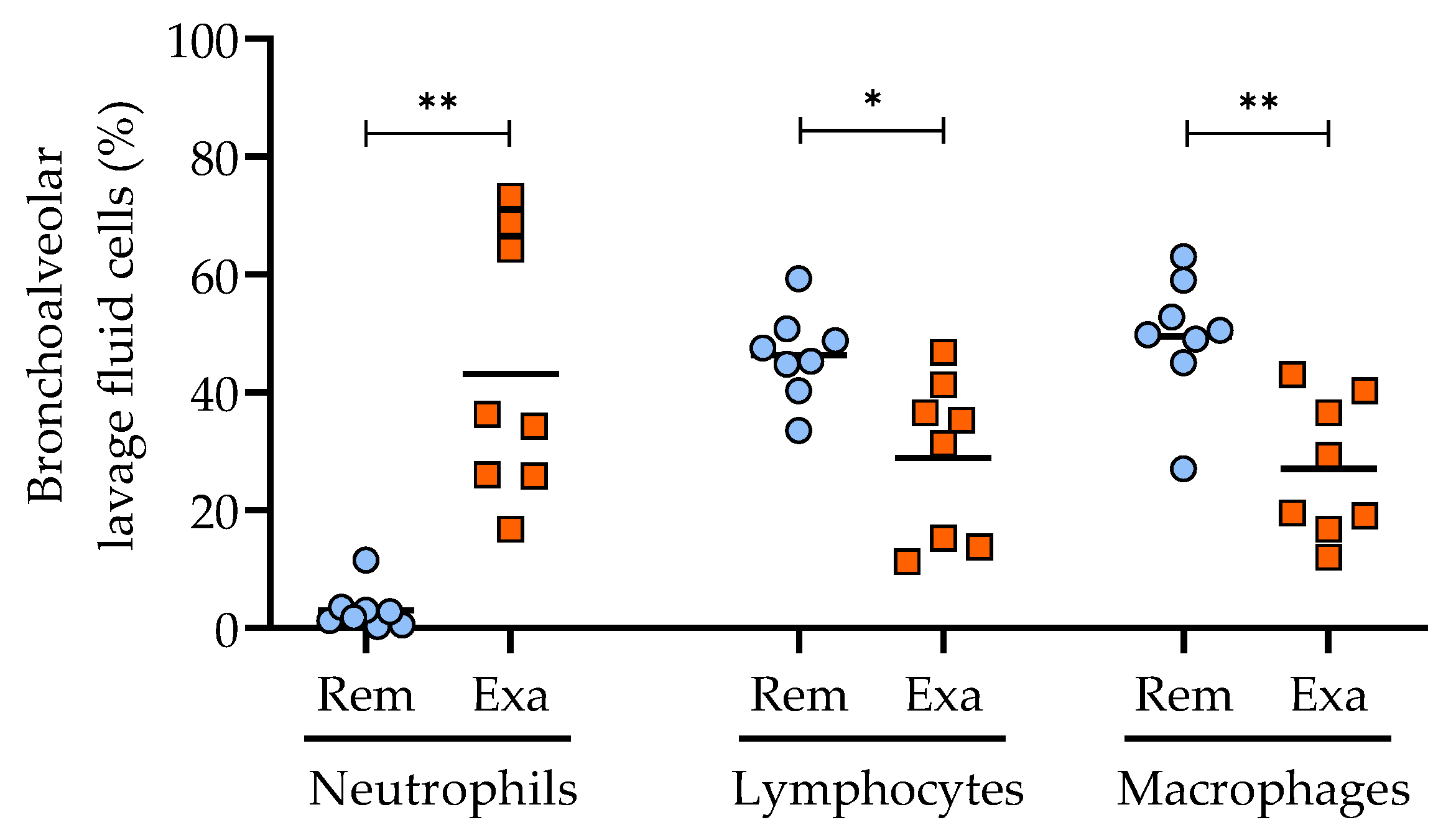
3.1. Characterization of the Clinical Status of Horses with Severe Asthma
3.2. Neutrophil-Derived Extracellular Vesicles
3.2.1. Peripheral Blood Neutrophils Isolation
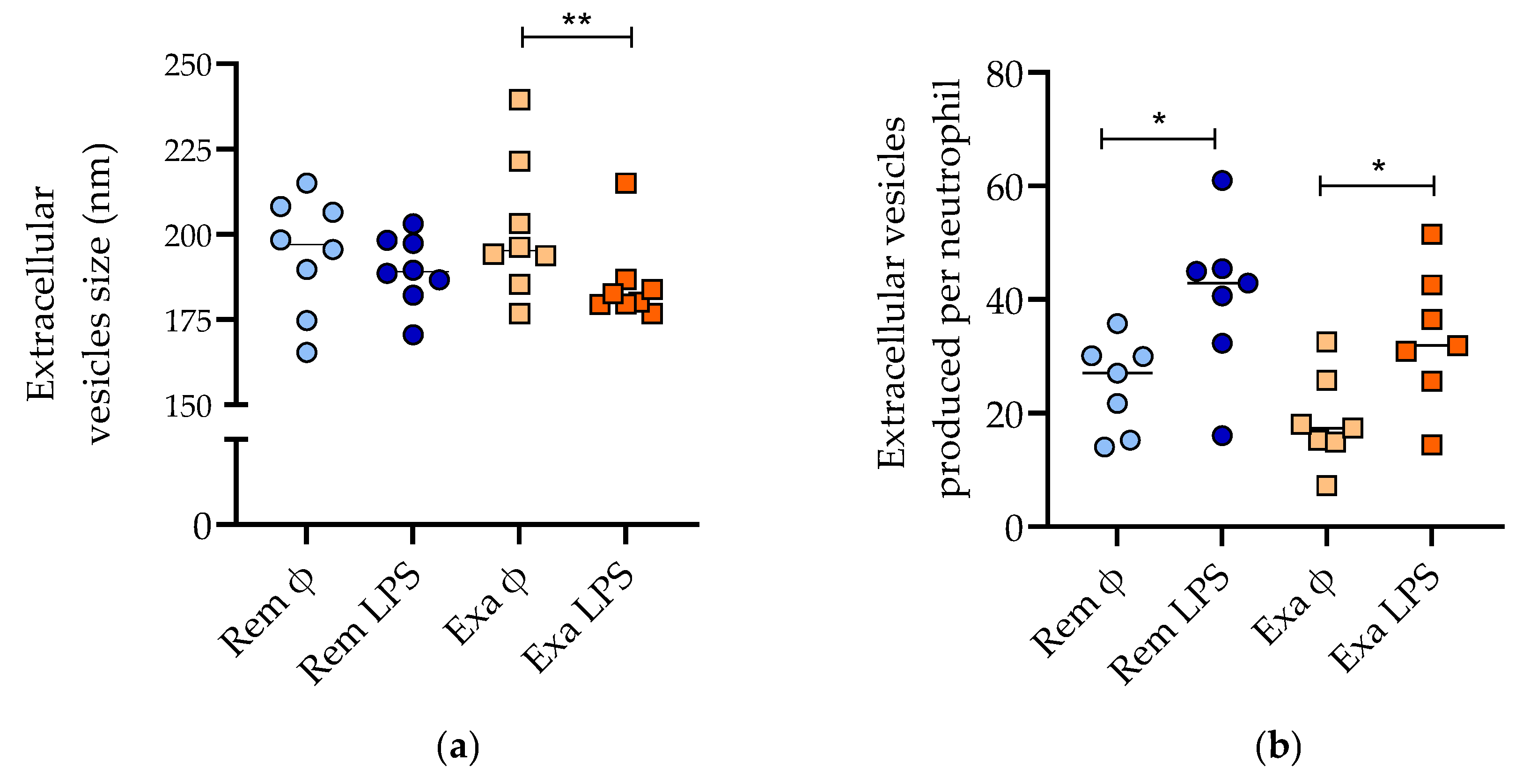
3.2.2. Nanoparticle Tracking Analysis
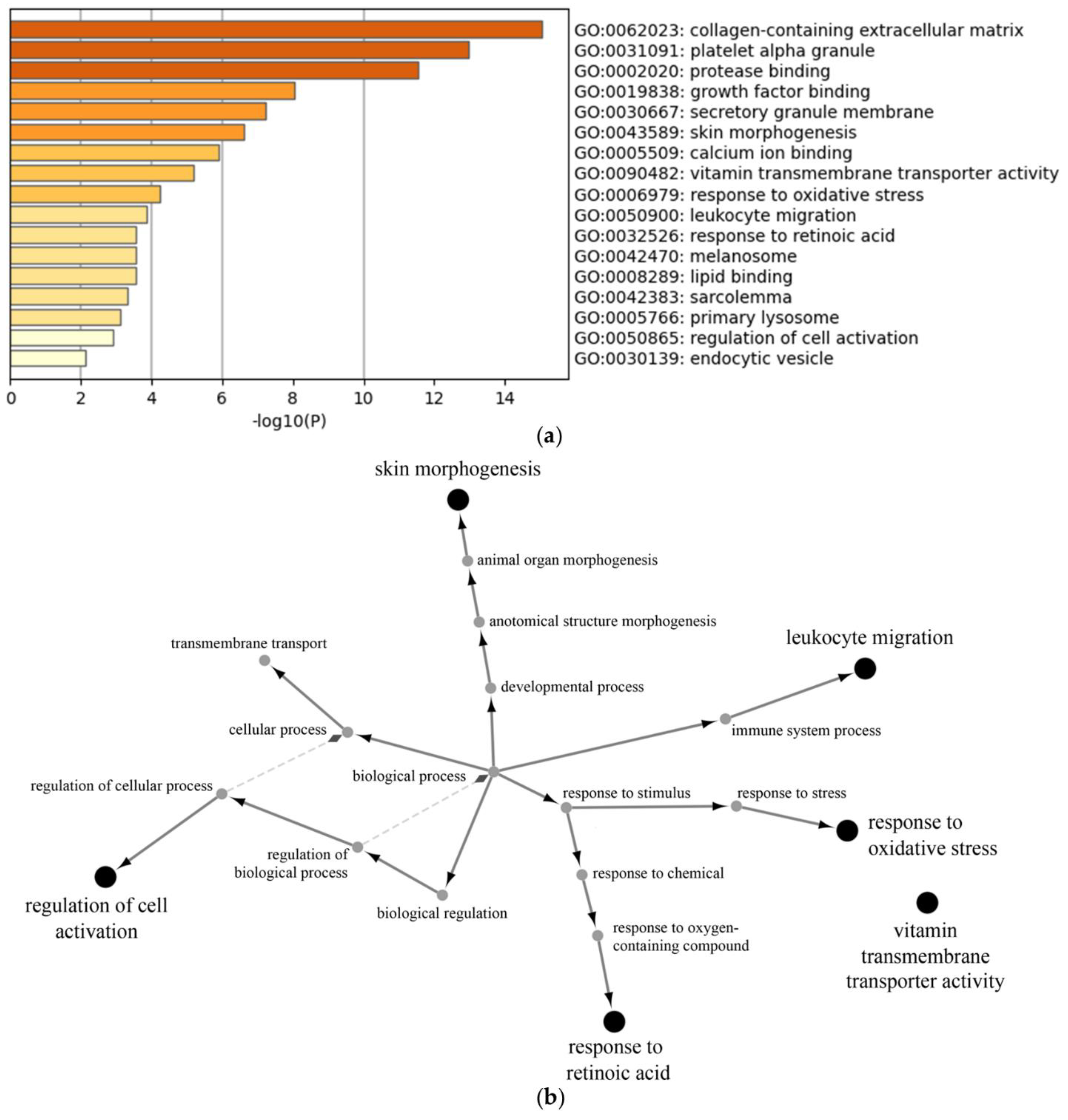
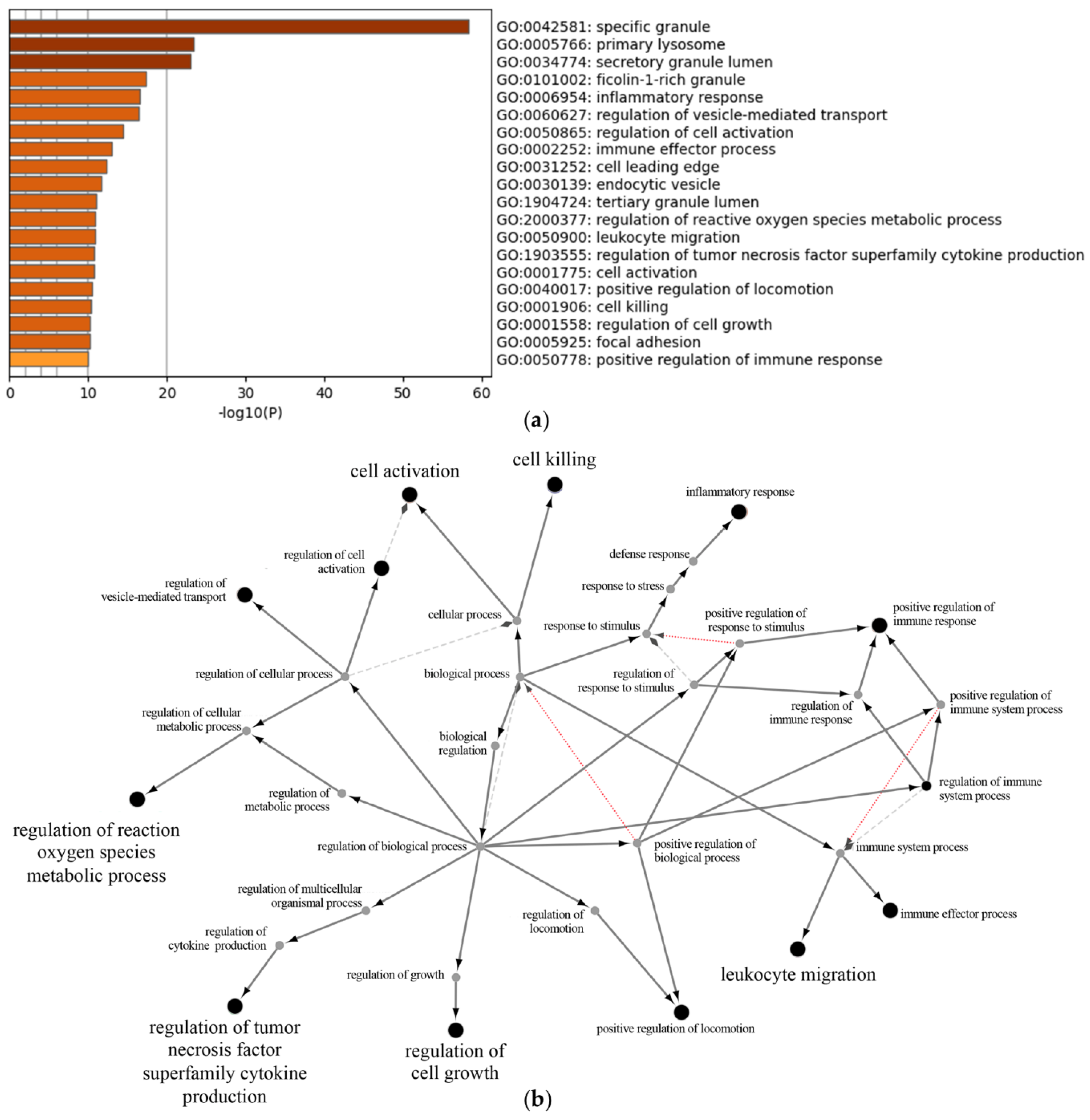
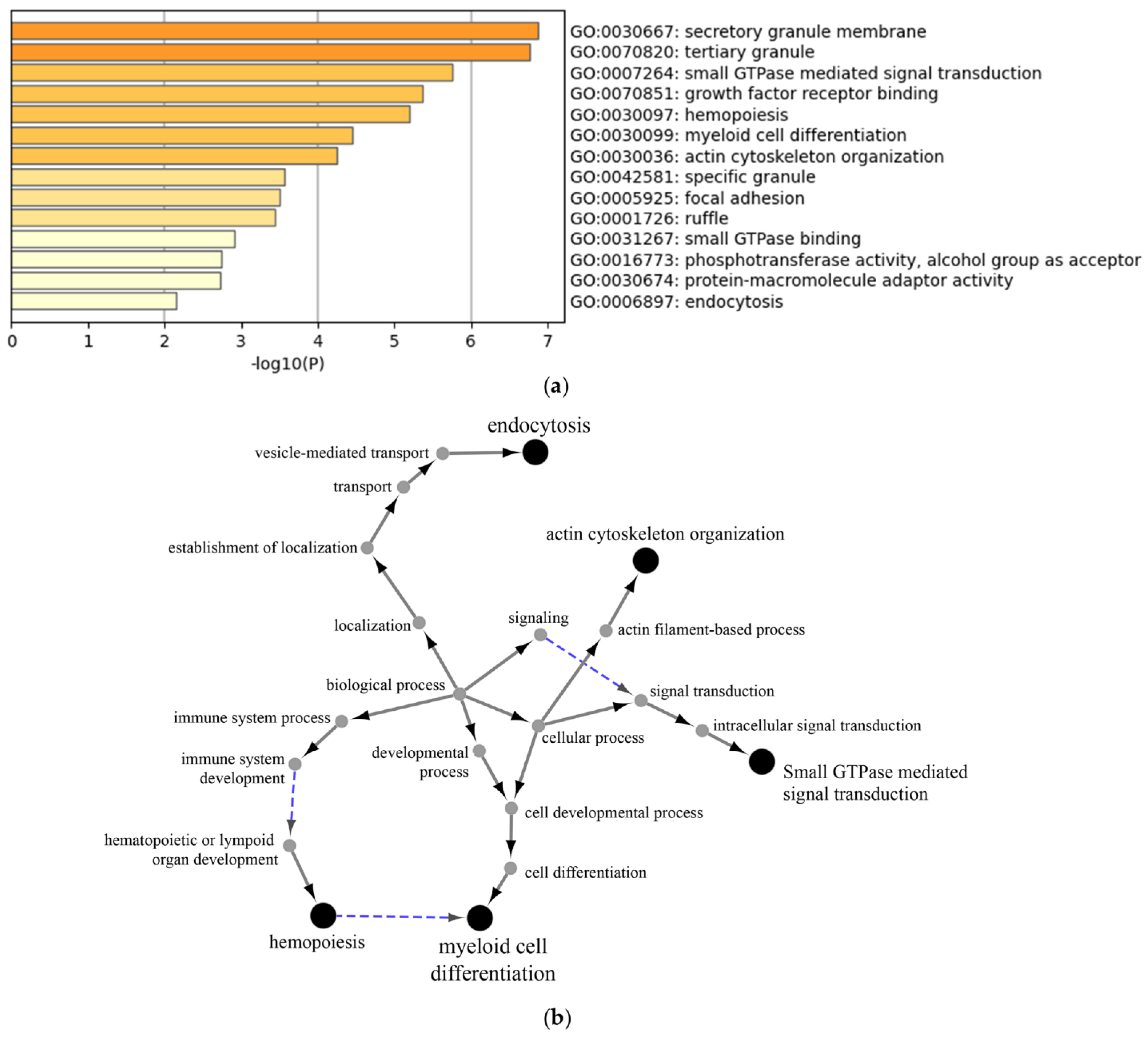
3.2.3. Proteomic Analysis of Neutrophil Extracellular Vesicles
3.3. Proliferative Effects of Extracellular Vesicles on Airway Smooth Muscle Cells
3.3.1. ASM Characterization
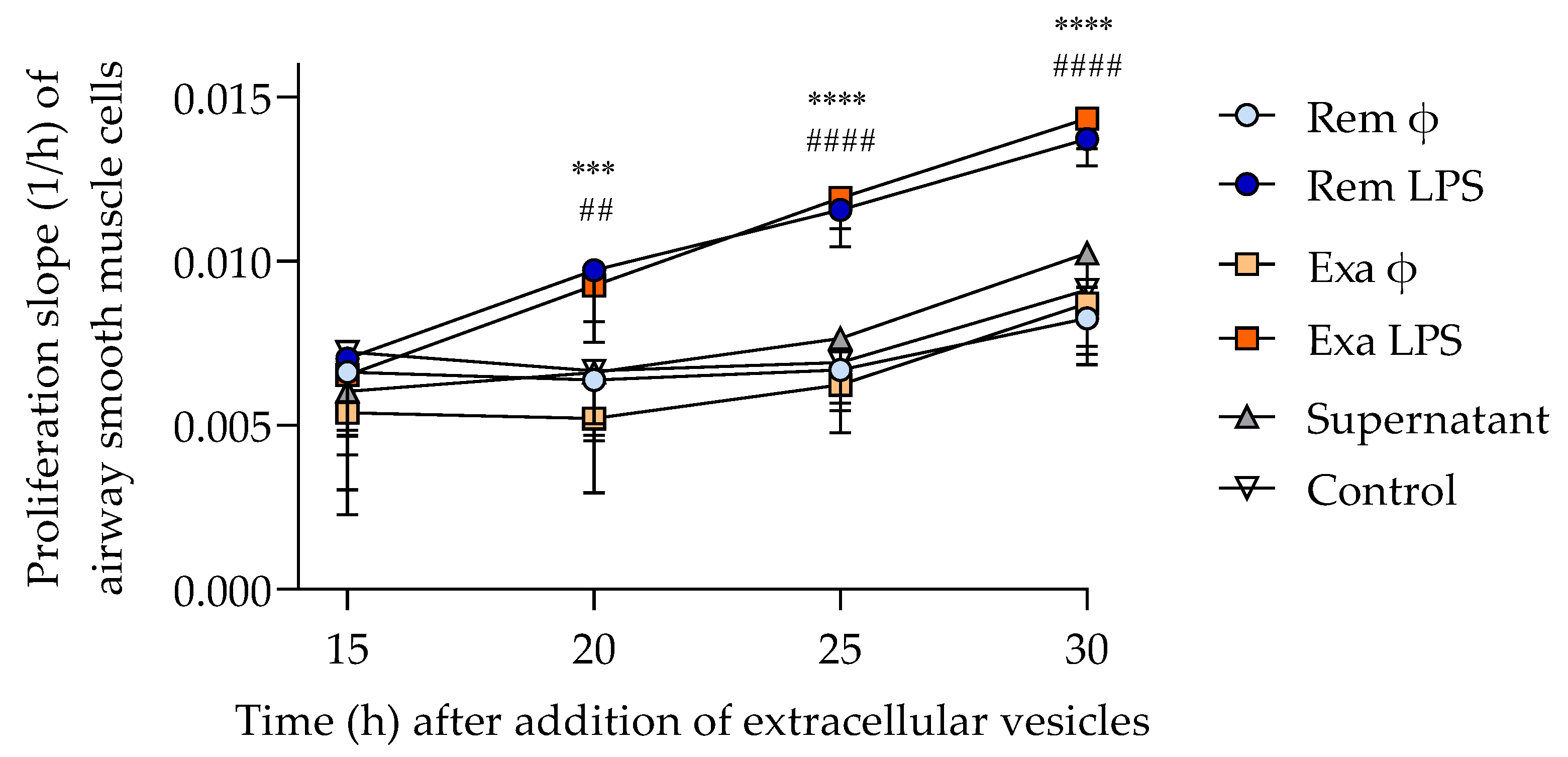
3.3.2. ASM Proliferation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Safiri, S.; Carson-Chahhoud, K.; Karamzad, N.; Sullman, M.J.M.; Nejadghaderi, S.A.; Taghizadieh, A.; Bell, A.W.; Kolahi, A.A.; Ansarin, K.; Mansournia, M.A.; et al. Prevalence, Deaths, and Disability-Adjusted Life-Years Due to Asthma and Its Attributable Risk Factors in 204 Countries and Territories, 1990–2019. Chest 2022, 161, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.E.; Szefler, S.J.; Leung, D.Y.; Sloan, S.I.; Rex, M.D.; Martin, R.J. Bronchoscopic evaluation of severe asthma. Persistent inflammation associated with high dose glucocorticoids. Am. J. Respir. Crit. Care Med. 1997, 156, 737–743. [Google Scholar] [CrossRef]
- Jatakanon, A.; Uasuf, C.; Maziak, W.; Lim, S.; Chung, K.F.; Barnes, P.J. Neutrophilic inflammation in severe persistent asthma. Am. J. Respir. Crit. Care Med. 1999, 160, 1532–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, N.; Carello, S.; Cooke, C.; James, A. Airway structure and inflammatory cells in fatal attacks of asthma. Eur. Respir. J. 1996, 9, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.M.; Zhao, N.; Olivenstein, R.; Lemiere, C.; Hamid, Q.; Martin, J.G. Asthma and fixed airflow obstruction: Long-term trajectories suggest distinct endotypes. Clin. Exp. Allergy 2021, 51, 39–48. [Google Scholar] [CrossRef]
- Bullone, M.; Lavoie, J.P. Asthma “of horses and men”—How can equine heaves help us better understand human asthma immunopathology and its functional consequences? Mol. Immunol. 2015, 66, 97–105. [Google Scholar] [CrossRef]
- Mainguy-Seers, S.; Boivin, R.; Pourali Dogaheh, S.; Beaudry, F.; Helie, P.; Bonilla, A.G.; Martin, J.G.; Lavoie, J.P. Effects of azithromycin on bronchial remodeling in the natural model of severe neutrophilic asthma in horses. Sci. Rep. 2022, 12, 446. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.D.; Chen, H.H.; Wang, C.H.; Chou, C.L.; Lin, S.M.; Lin, H.C.; Kuo, H.P. Human neutrophil-derived elastase induces airway smooth muscle cell proliferation. Life Sci. 2004, 74, 2479–2492. [Google Scholar] [CrossRef]
- Malavia, N.K.; Raub, C.B.; Mahon, S.B.; Brenner, M.; Panettieri, R.A., Jr.; George, S.C. Airway epithelium stimulates smooth muscle proliferation. Am. J. Respir. Cell Mol. Biol. 2009, 41, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Begueret, H.; Berger, P.; Vernejoux, J.M.; Dubuisson, L.; Marthan, R.; Tunon-de-Lara, J.M. Inflammation of bronchial smooth muscle in allergic asthma. Thorax 2007, 62, 8–15. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Roux-Dalvai, F.; Droit, A.; Lavoie, J.P. Neutrophil-Derived Exosomes: A New Mechanism Contributing to Airway Smooth Muscle Remodeling. Am. J. Respir. Cell Mol. Biol. 2016, 55, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Chen, Z.C.; Li, N.; Wang, Z.H.; Guo, Y.L.; Tian, C.J.; Cheng, D.J.; Tang, X.Y.; Zhang, L.X. Exosomal transfer of activated neutrophil-derived lncRNA CRNDE promotes proliferation and migration of airway smooth muscle cells in asthma. Hum. Mol. Genet 2021, 31, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Lapa e Silva, J.R.; Possebon da Silva, M.D.; Lefort, J.; Vargaftig, B.B. Endotoxins, asthma, and allergic immune responses. Toxicology 2000, 152, 31–35. [Google Scholar] [CrossRef]
- Michel, O.; Ginanni, R.; Duchateau, J.; Vertongen, F.; Le Bon, B.; Sergysels, R. Domestic endotoxin exposure and clinical severity of asthma. Clin. Exp. Allergy 1991, 21, 441–448. [Google Scholar] [CrossRef]
- Pirie, R.S.; Collie, D.D.; Dixon, P.M.; McGorum, B.C. Inhaled endotoxin and organic dust particulates have synergistic proinflammatory effects in equine heaves (organic dust-induced asthma). Clin. Exp. Allergy 2003, 33, 676–683. [Google Scholar] [CrossRef]
- Pirie, R.S.; Dixon, P.M.; Collie, D.D.; McGorum, B.C. Pulmonary and systemic effects of inhaled endotoxin in control and heaves horses. Equine. Vet. J. 2001, 33, 311–318. [Google Scholar] [CrossRef]
- Mendy, A.; Wilkerson, J.; Salo, P.M.; Zeldin, D.C.; Thorne, P.S. Endotoxin clustering with allergens in house dust and asthma outcomes in a U.S. national study. Environ. Health 2020, 19, 35. [Google Scholar] [CrossRef] [Green Version]
- Siegers, E.W.; Anthonisse, M.; van Eerdenburg, F.; van den Broek, J.; Wouters, I.M.; Westermann, C.M. Effect of ionization, bedding, and feeding on air quality in a horse stable. J. Vet. Intern. Med. 2018, 32, 1234–1240. [Google Scholar] [CrossRef] [Green Version]
- Jochmans-Lemoine, A.; Picotte, K.; Beauchamp, G.; Vargas, A.; Lavoie, J.P. Effects of a propriety oiled mixed hay feeding system on lung function, neutrophilic airway inflammation and oxidative stress in severe asthmatic horses. Equine. Vet. J. 2020, 52, 564–571. [Google Scholar] [CrossRef]
- van Erck, E.; Votion, D.; Art, T.; Lekeux, P. Measurement of respiratory function by impulse oscillometry in horses. Equine. Vet. J. 2004, 36, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bickel, S.; Popler, J.; Lesnick, B.; Eid, N. Impulse oscillometry: Interpretation and practical applications. Chest 2014, 146, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Desai, U.; Joshi, J.M. Impulse oscillometry. Adv. Respir. Med. 2019, 87, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoie, J.P.; Pasloske, K.; Joubert, P.; Cordeau, M.E.; Mancini, J.; Girard, Y.; Friesen, R.W.; Frenette, R.; Blouin, M.; Young, R.N.; et al. Lack of clinical efficacy of a phosphodiesterase-4 inhibitor for treatment of heaves in horses. J. Vet. Intern. Med. 2006, 20, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Bullone, M.; Chevigny, M.; Allano, M.; Martin, J.G.; Lavoie, J.P. Technical and physiological determinants of airway smooth muscle mass in endobronchial biopsy samples of asthmatic horses. J. Appl. Physiol. 2014, 117, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Vargas, A.; Peltier, A.; Dube, J.; Lefebvre-Lavoie, J.; Moulin, V.; Goulet, F.; Lavoie, J.P. Evaluation of contractile phenotype in airway smooth muscle cells isolated from endobronchial biopsy and tissue specimens from horses. Am. J. Vet. Res. 2017, 78, 359–370. [Google Scholar] [CrossRef]
- Kolonics, F.; Szeifert, V.; Timar, C.I.; Ligeti, E.; Lorincz, A.M. The Functional Heterogeneity of Neutrophil-Derived Extracellular Vesicles Reflects the Status of the Parent Cell. Cells 2020, 9, 2718. [Google Scholar] [CrossRef]
- Alashkar Alhamwe, B.; Potaczek, D.P.; Miethe, S.; Alhamdan, F.; Hintz, L.; Magomedov, A.; Garn, H. Extracellular Vesicles and Asthma-More Than Just a Co-Existence. Int. J. Mol. Sci. 2021, 22, 4984. [Google Scholar] [CrossRef]
- Abraham, E.; Nick, J.A.; Azam, T.; Kim, S.H.; Mira, J.P.; Svetkauskaite, D.; He, Q.; Zamora, M.; Murphy, J.; Park, J.S.; et al. Peripheral blood neutrophil activation patterns are associated with pulmonary inflammatory responses to lipopolysaccharide in humans. J. Immunol. 2006, 176, 7753–7760. [Google Scholar] [CrossRef] [Green Version]
- Baines, K.J.; Simpson, J.L.; Bowden, N.A.; Scott, R.J.; Gibson, P.G. Differential gene expression and cytokine production from neutrophils in asthma phenotypes. Eur. Respir. J. 2010, 35, 522–531. [Google Scholar] [CrossRef]
- Herteman, N.; Grimes, C.; Lavoie, J.P. Retrospective investigation of automated hematology analyzer-determined indicators of neutrophil activation in blood samples from horses with asthma. Am. J. Vet. Res. 2021, 82, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Vachier, I.; Chanez, P.; Le Doucen, C.; Damon, M.; Descomps, B.; Godard, P. Enhancement of reactive oxygen species formation in stable and unstable asthmatic patients. Eur. Respir. J. 1994, 7, 1585–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, L.; Patino, P.J.; Montoya, F.; Vanegas, A.C.; Echavarria, A.; Garcia de Olarte, D. A study of granulocyte respiratory burst in patients with allergic bronchial asthma. Inflammation 1998, 22, 45–54. [Google Scholar] [CrossRef]
- Mann, B.S.; Chung, K.F. Blood neutrophil activation markers in severe asthma: Lack of inhibition by prednisolone therapy. Respir. Res. 2006, 7, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazil, T.J.; Dagleish, M.P.; McGorum, B.C.; Dixon, P.M.; Haslett, C.; Chilvers, E.R. Kinetics of pulmonary neutrophil recruitment and clearance in a natural and spontaneously resolving model of airway inflammation. Clin. Exp. Allergy 2005, 35, 854–865. [Google Scholar] [CrossRef]
- Aloor, J.J.; Azzam, K.M.; Guardiola, J.J.; Gowdy, K.M.; Madenspacher, J.H.; Gabor, K.A.; Mueller, G.A.; Lin, W.C.; Lowe, J.M.; Gruzdev, A.; et al. Leucine-rich repeats and calponin homology containing 4 (Lrch4) regulates the innate immune response. J. Biol. Chem. 2019, 294, 1997–2008. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.L.; Grissell, T.V.; Douwes, J.; Scott, R.J.; Boyle, M.J.; Gibson, P.G. Innate immune activation in neutrophilic asthma and bronchiectasis. Thorax 2007, 62, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Esnault, S.; Kelly, E.A.; Schwantes, E.A.; Liu, L.Y.; DeLain, L.P.; Hauer, J.A.; Bochkov, Y.A.; Denlinger, L.C.; Malter, J.S.; Mathur, S.K.; et al. Identification of genes expressed by human airway eosinophils after an in vivo allergen challenge. PLoS ONE 2013, 8, e67560. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.H.; Li, Z.; Zeng, T.; Chen, L.; Li, H.; Huang, T.; Cai, Y.D. Detecting the Multiomics Signatures of Factor-Specific Inflammatory Effects on Airway Smooth Muscles. Front. Genet. 2020, 11, 599970. [Google Scholar] [CrossRef]
- Davis, K.U.; Sheats, M.K. Differential gene expression and Ingenuity Pathway Analysis of bronchoalveolar lavage cells from horses with mild/moderate neutrophilic or mastocytic inflammation on BAL cytology. Vet. Immunol. Immunopathol. 2021, 234, 110195. [Google Scholar] [CrossRef]
- Hauk, P.J.; Krawiec, M.; Murphy, J.; Boguniewicz, J.; Schiltz, A.; Goleva, E.; Liu, A.H.; Leung, D.Y. Neutrophilic airway inflammation and association with bacterial lipopolysaccharide in children with asthma and wheezing. Pediatr. Pulmonol. 2008, 43, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Vignola, A.M.; Bonanno, A.; Profita, M.; Riccobono, L.; Scichilone, N.; Spatafora, M.; Bousquet, J.; Bonsignore, G.; Bellia, V. Effect of age and asthma duration upon elastase and alpha1-antitrypsin levels in adult asthmatics. Eur. Respir. J. 2003, 22, 795–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasaian, M.T.; Lee, J.; Brennan, A.; Danto, S.I.; Black, K.E.; Fitz, L.; Dixon, A.E. Proteomic analysis of serum and sputum analytes distinguishes controlled and poorly controlled asthmatics. Clin. Exp. Allergy 2018, 48, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Raulo, S.M.; Sorsa, T.; Tervahartiala, T.; Pirila, E.; Maisi, P. MMP-9 as a marker of inflammation in tracheal epithelial lining fluid (TELF) and in bronchoalveolar fluid (BALF) of COPD horses. Equine. Vet. J. 2001, 33, 128–136. [Google Scholar] [CrossRef]
- Simonen-Jokinen, T.; Pirie, R.S.; McGorum, B.; Maisi, P. Dose responses to inhalation of endotoxin, hay dust suspension and Aspergillus fumigatus extract in horses as measured by levels and activation of matrix metalloproteinase-9. Equine. Vet. J. 2005, 37, 155–160. [Google Scholar] [CrossRef]
- Kawagoe, J.; Kono, Y.; Togashi, Y.; Ishiwari, M.; Toriyama, K.; Yajima, C.; Nakayama, H.; Kasagi, S.; Abe, S.; Setoguchi, Y. Serum Neutrophil Gelatinase-associated Lipocalin (NGAL) Is Elevated in Patients with Asthma and Airway Obstruction. Curr. Med. Sci. 2021, 41, 323–328. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, M.; Yan, X.; Cao, L.; Barnes, P.J.; Adcock, I.M.; Huang, M.; Yao, X. Increased neutrophil gelatinase-associated lipocalin (NGAL) promotes airway remodelling in chronic obstructive pulmonary disease. Clin. Sci. 2017, 131, 1147–1159. [Google Scholar] [CrossRef]
- Lavoie, J.P.; Lefebvre-Lavoie, J.; Leclere, M.; Lavoie-Lamoureux, A.; Chamberland, A.; Laprise, C.; Lussier, J. Profiling of differentially expressed genes using suppression subtractive hybridization in an equine model of chronic asthma. PLoS ONE 2012, 7, e29440. [Google Scholar] [CrossRef]
- Svensson Holm, A.C.; Bengtsson, T.; Grenegard, M.; Lindstrom, E.G. Platelets stimulate airway smooth muscle cell proliferation through mechanisms involving 5-lipoxygenase and reactive oxygen species. Platelets 2008, 19, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A. Inflammation signals airway smooth muscle cell proliferation in asthma pathogenesis. Multidiscip. Respir. Med. 2013, 8, 11. [Google Scholar] [CrossRef]
- Naveed, S.U.; Clements, D.; Jackson, D.J.; Philp, C.; Billington, C.K.; Soomro, I.; Reynolds, C.; Harrison, T.W.; Johnston, S.L.; Shaw, D.E.; et al. Matrix Metalloproteinase-1 Activation Contributes to Airway Smooth Muscle Growth and Asthma Severity. Am. J. Respir. Crit. Care Med. 2017, 195, 1000–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Wang, Y.; Liao, G.; Chen, B.; Panettieri, R.A., Jr.; Penn, R.B.; Tang, D.D. Abi1 mediates airway smooth muscle cell proliferation and airway remodeling via Jak2/STAT3 signaling. iScience 2022, 25, 103833. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Noveral, J.P.; Bhala, A.; Nunn, S.E.; Herrick, D.J.; Grunstein, M.M. Leukotriene D4 facilitates airway smooth muscle cell proliferation via modulation of the IGF axis. Am. J. Physiol. 1995, 269, L151–L157. [Google Scholar] [CrossRef] [PubMed]
- Colotta, F.; Re, F.; Polentarutti, N.; Sozzani, S.; Mantovani, A. Modulation of granulocyte survival and programmed cell death by cytokines and bacterial products. Blood 1992, 80, 2012–2020. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.L.; Townsend, H.G.G.; Singh, B. Role of toll-like receptor 4 and caspase-3, -8, and -9 in lipopolysaccharide-induced delay of apoptosis in equine neutrophils. Am. J. Vet. Res. 2018, 79, 424–432. [Google Scholar] [CrossRef]
- Chen, J.Y.; Cheng, W.H.; Lee, K.Y.; Kuo, H.P.; Chung, K.F.; Chen, C.L.; Chen, B.C.; Lin, C.H. Abnormal ADAM17 expression causes airway fibrosis in chronic obstructive asthma. Biomed. Pharm. 2021, 140, 111701. [Google Scholar] [CrossRef]
- Do, A.N.; Chun, Y.; Grishina, G.; Grishin, A.; Rogers, A.J.; Raby, B.A.; Weiss, S.T.; Vicencio, A.; Schadt, E.E.; Bunyavanich, S. Network study of nasal transcriptome profiles reveals master regulator genes of asthma. J. Allergy Clin. Immunol. 2021, 147, 879–893. [Google Scholar] [CrossRef]
- Green, T.D.; Crews, A.L.; Park, J.; Fang, S.; Adler, K.B. Regulation of mucin secretion and inflammation in asthma: A role for MARCKS protein? Biochim. Biophys. Acta 2011, 1810, 1110–1113. [Google Scholar] [CrossRef] [Green Version]
- Sai, J.; Raman, D.; Liu, Y.; Wikswo, J.; Richmond, A. Parallel phosphatidylinositol 3-kinase (PI3K)-dependent and Src-dependent pathways lead to CXCL8-mediated Rac2 activation and chemotaxis. J. Biol. Chem. 2008, 283, 26538–26547. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.K.; Starnes, T.W.; Deng, Q.; Huttenlocher, A. Lyn is a redox sensor that mediates leukocyte wound attraction in vivo. Nature 2011, 480, 109–112. [Google Scholar] [CrossRef]
- Frelin, C.; Ofran, Y.; Ruston, J.; Hayun, M.; Derdikman, Y.; Khier, Y.; Rozales, K.; Brenner, B.; Iscove, N.; Pawson, T.; et al. Grb2 regulates the proliferation of hematopoietic stem and progenitors cells. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2449–2459. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.; Taveira-Gomes, T.; Sokhatska, O.; Palmares, C.; Costa, R.; Negrao, R.; Guimaraes, J.T.; Delgado, L.; Soares, R.; Moreira, A. Increased circulating platelet microparticles as a potential biomarker in asthma. Allergy 2013, 68, 1073–1075. [Google Scholar] [CrossRef] [PubMed]
- Elliot, J.G.; Jones, R.L.; Abramson, M.J.; Green, F.H.; Mauad, T.; McKay, K.O.; Bai, T.R.; James, A.L. Distribution of airway smooth muscle remodelling in asthma: Relation to airway inflammation. Respirology 2015, 20, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; McGettrick, H.M.; Stone, P.C.; Rainger, G.E.; Nash, G.B. The roles of integrins in function of human neutrophils after their migration through endothelium into interstitial matrix. PLoS ONE 2015, 10, e0118593. [Google Scholar] [CrossRef] [Green Version]
- Tak, T.; Hilvering, B.; Tesselaar, K.; Koenderman, L. Similar activation state of neutrophils in sputum of asthma patients irrespective of sputum eosinophilia. Clin. Exp. Immunol. 2015, 182, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Baines, K.J.; Simpson, J.L.; Scott, R.J.; Gibson, P.G. Immune responses of airway neutrophils are impaired in asthma. Exp. Lung Res. 2009, 35, 554–569. [Google Scholar] [CrossRef]
- Brazil, T.J. Pulmonary Neutrophil Recruitment, Activation and Clearance in Equine Chronic Obstructive Pulmonary Disease. Ph.D. Thesis, The University of Edinburgh, Edinburgh, Scotland, 1999. [Google Scholar]
- Dunkel, B.; Rickards, K.J.; Werling, D.; Page, C.P.; Cunningham, F.M. Neutrophil and platelet activation in equine recurrent airway obstruction is associated with increased neutrophil CD13 expression, but not platelet CD41/61 and CD62P or neutrophil-platelet aggregate formation. Vet. Immunol. Immunopathol. 2009, 131, 25–32. [Google Scholar] [CrossRef]
- Koblinger, K.; Hecker, K.; Nicol, J.; Wasko, A.; Fernandez, N.; Leguillette, R. Bronchial collapse during bronchoalveolar lavage in horses is an indicator of lung inflammation. Equine. Vet. J. 2014, 46, 50–55. [Google Scholar] [CrossRef]
- Douanne, N.; Dong, G.; Amin, A.; Bernardo, L.; Blanchette, M.; Langlais, D.; Olivier, M.; Fernandez-Prada, C. Leishmania parasites exchange drug-resistance genes through extracellular vesicles. Cell Rep. 2022, 40, 111121. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mainguy-Seers, S.; Beaudry, F.; Fernandez-Prada, C.; Martin, J.G.; Lavoie, J.-P. Neutrophil Extracellular Vesicles and Airway Smooth Muscle Proliferation in the Natural Model of Severe Asthma in Horses. Cells 2022, 11, 3347. https://doi.org/10.3390/cells11213347
Mainguy-Seers S, Beaudry F, Fernandez-Prada C, Martin JG, Lavoie J-P. Neutrophil Extracellular Vesicles and Airway Smooth Muscle Proliferation in the Natural Model of Severe Asthma in Horses. Cells. 2022; 11(21):3347. https://doi.org/10.3390/cells11213347
Chicago/Turabian StyleMainguy-Seers, Sophie, Francis Beaudry, Christopher Fernandez-Prada, James G. Martin, and Jean-Pierre Lavoie. 2022. "Neutrophil Extracellular Vesicles and Airway Smooth Muscle Proliferation in the Natural Model of Severe Asthma in Horses" Cells 11, no. 21: 3347. https://doi.org/10.3390/cells11213347