
The Electrostatic Basis of Diacylglycerol Pyrophosphate—Protein Interaction

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. NMR Spectroscopy
2.4. Charge Determination from Titration Curve Data
2.5. Statistical Analysis
3. Results and Discussion
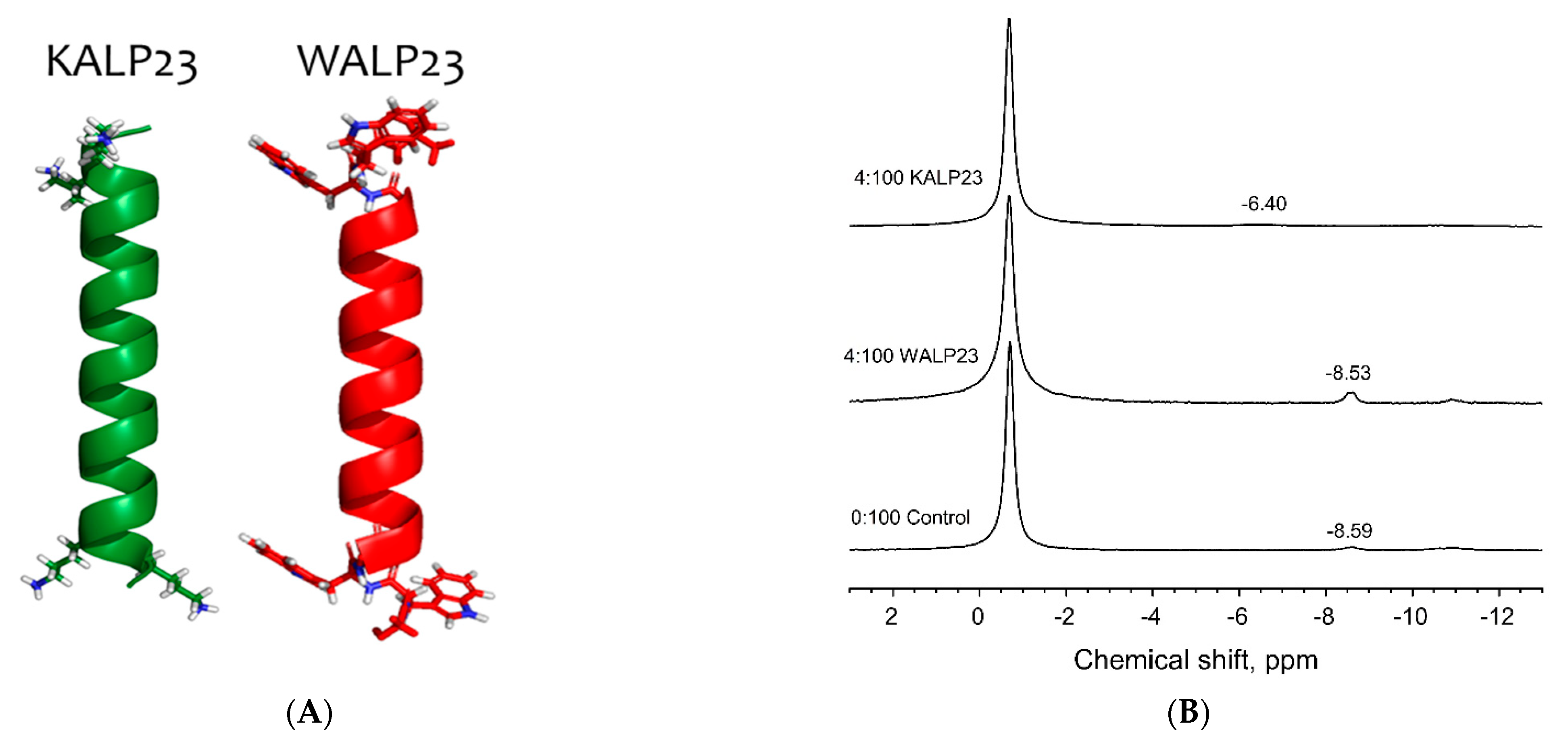
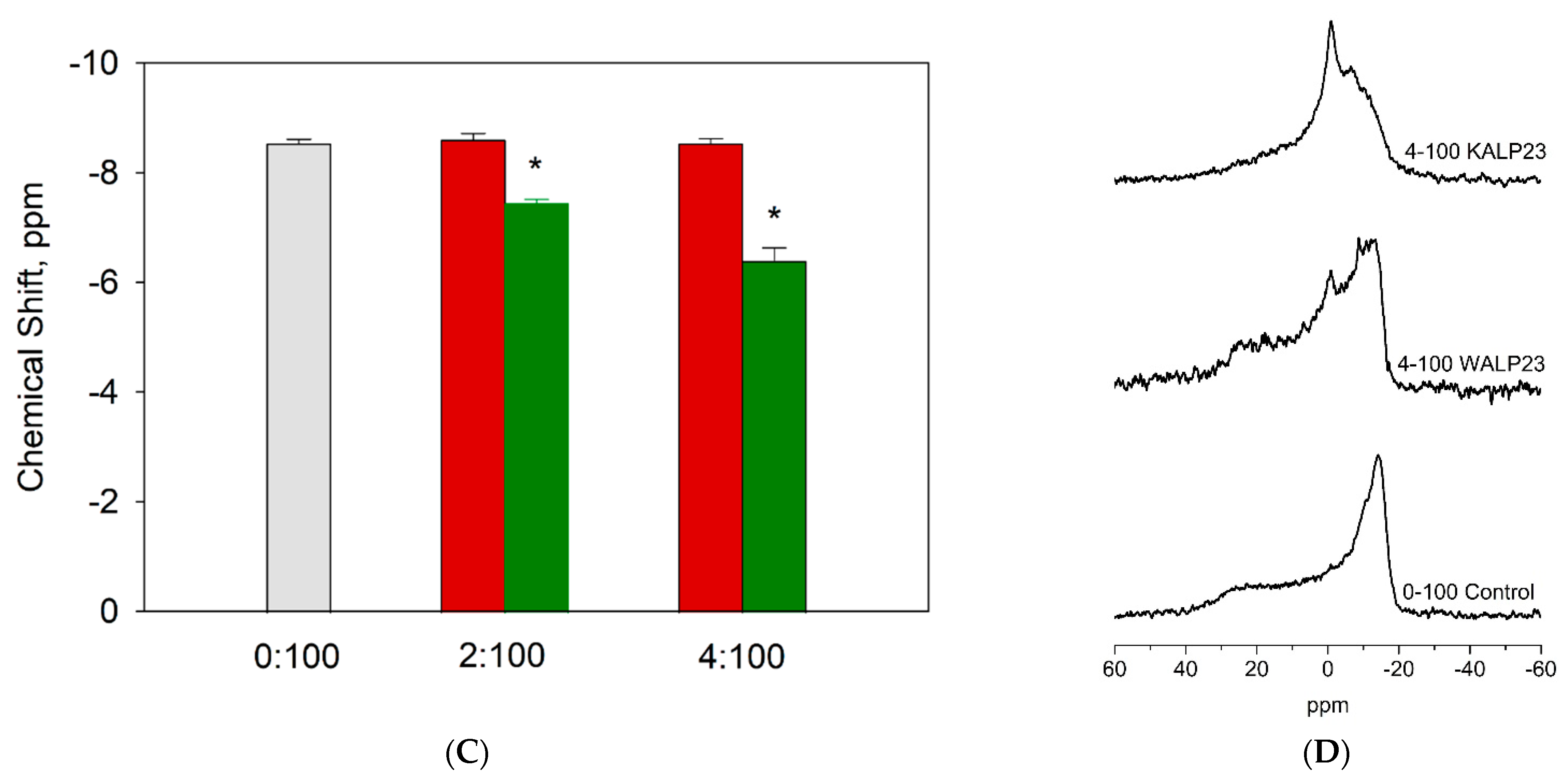
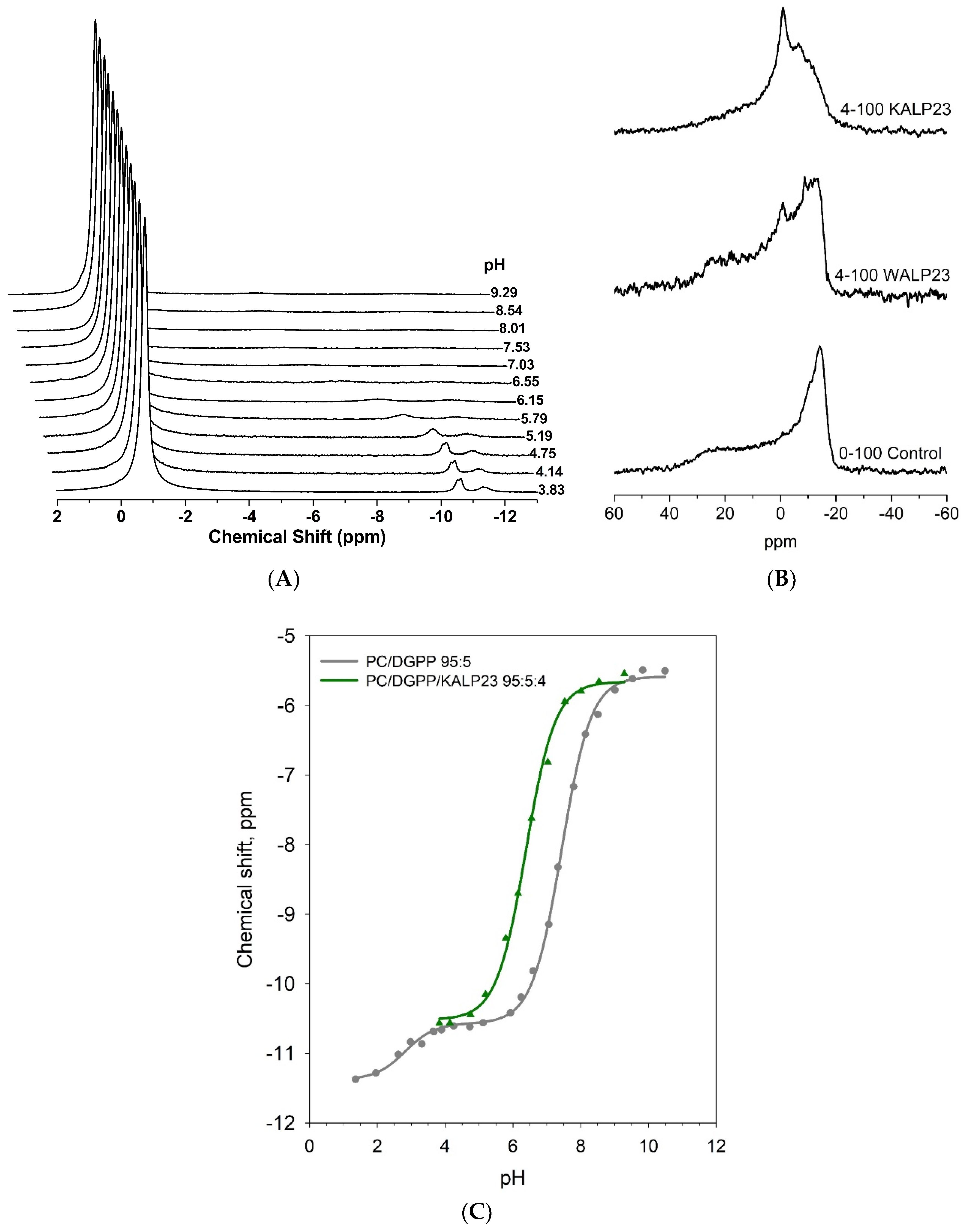
3.1. Lysine Residues Interact Strongly with the Pyrophosphate Headgroup of DGPP and Increase Its Charge
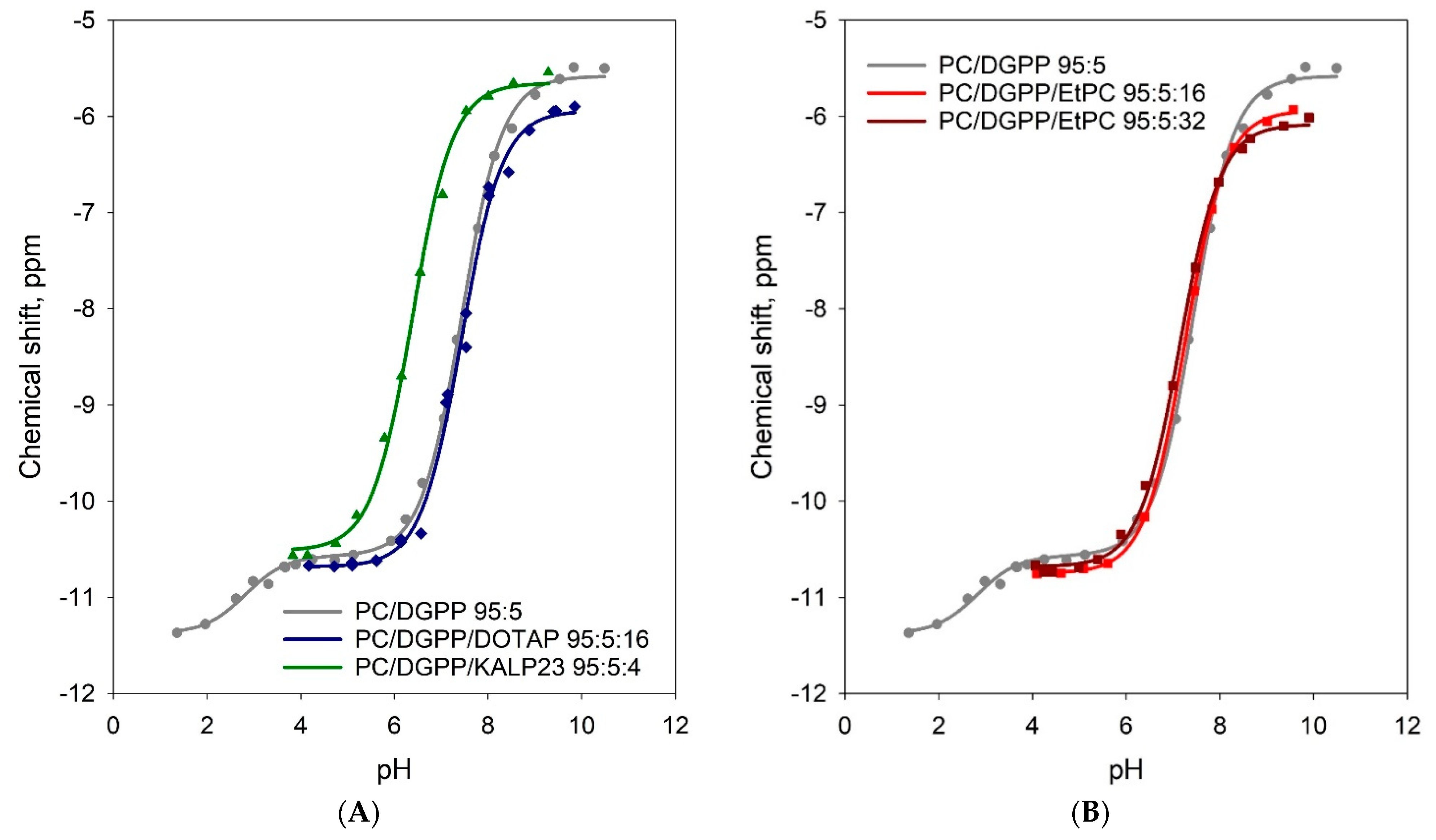
3.2. Positive Charge Alone Does Not Significantly Affect the Charge of the Phosphomonoester of DGPP
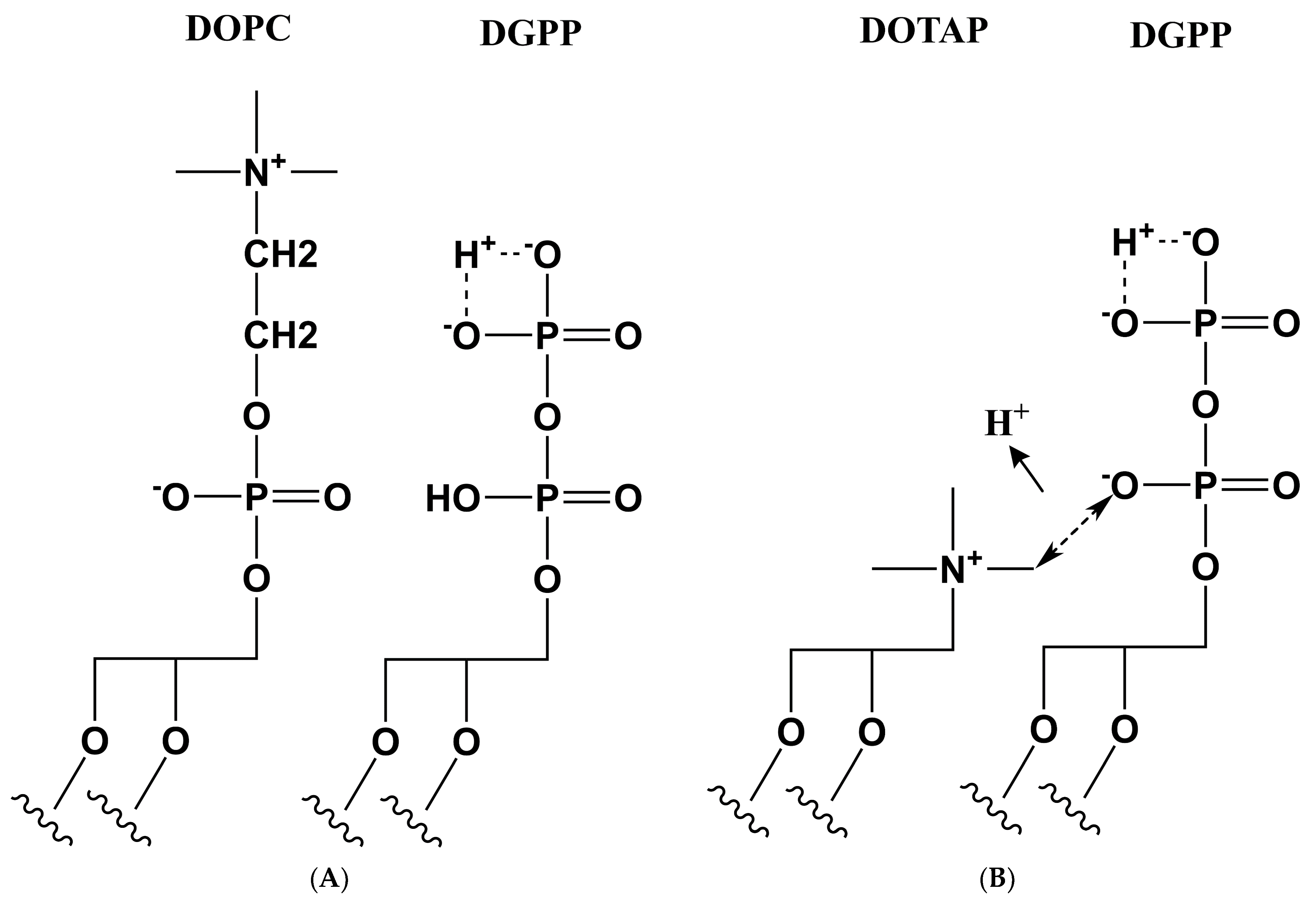
3.3. Proposed Model for the Effect of Positive Charge on the Ionization of DGPP
3.4. Implications for DGPP—Protein Interactions
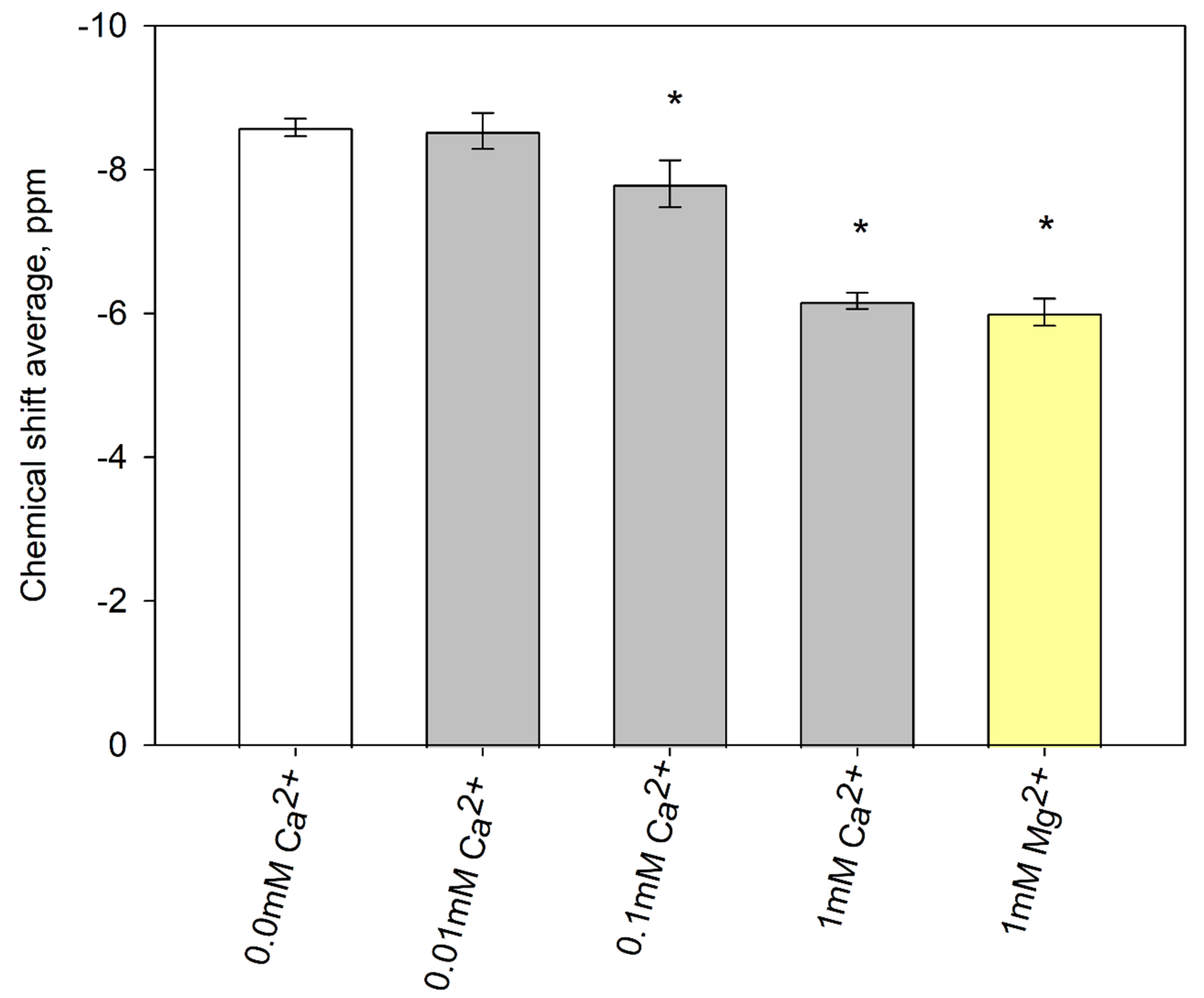
3.5. The interaction of DGPP with Divalent Cations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef]
- De Craene, J.O.; Bertazzi, D.L.; Bär, S.; Friant, S. Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways. Int. J. Mol. Sci. 2017, 18, 634. [Google Scholar] [CrossRef]
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef]
- Kooijman, E.E.; Chupin, V.; de Kruijff, B.; Burger, K.N.J. Modulation of Membrane curvature by phosphatidic acid and lysophosphatidic acid. Traffic 2003, 4, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Schink, K.O.; Tan, K.W.; Stenmark, H. Phosphoinositides in Control of Membrane Dynamics. Annu. Rev. Cell Dev. Biol. 2016, 32, 143–171. [Google Scholar] [CrossRef]
- Stace, C.L.; Ktistakis, N.T. Phosphatidic acid- and phosphatidylserine-binding proteins. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2006, 1761, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Jin, R.; Robustelli, J.; Zimmerman, Z.E.; Baumgart, T. PIP2 Reshapes Membranes through Asymmetric Desorption. Biophys. J. 2019, 117, 962–974. [Google Scholar] [CrossRef]
- Den Hartog, M.; Musgrave, A.; Munnik, T. Nod factor-induced phosphatidic acid and diacylglycerol pyrophosphate formation: A role for phospholipase C and D in root hair deformation. Plant J. 2001, 25, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Marchesini, N.; Santander, V.; Machado-Domenech, E. Diacylglycerol pyrophosphate: A novel metabolite in the Trypanosoma cruzi phosphatidic acid metabolism. FEBS Lett. 1998, 436, 377–381. [Google Scholar] [CrossRef]
- Munnik, T.; De Vrije, T.; Irvine, R.F.; Musgrave, A. Identification of diacylglycerol pyrophosphate as a novel metabolic product of phosphatidic acid during G-protein activation in plants. J. Biol. Chem. 1996, 271, 15708–15715. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef]
- Vilchez, A.C.; Peppino Margutti, M.; Reyna, M.; Wilke, N.; Villasuso, A.L. Recovery from chilling modulates the acyl-editing of phosphatidic acid molecular species in barley roots (Hordeum vulgare L.). Plant Physiol. Biochem. 2021, 167, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Zalejski, C.; Zhang, Z.; Quettier, A.L.; Maldiney, R.; Bonnet, M.; Brault, M.; Demandre, C.; Miginiac, E.; Rona, J.P.; Sotta, B.; et al. Diacylglycerol pyrophosphate is a second messenger of abscisic acid signaling in Arabidopsis thaliana suspension cells. Plant J. 2005, 42, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Munnik, T.; Meijer, H.J.G.; Ter Riet, B.; Hirt, H.; Frank, W.; Bartels, D.; Musgrave, A. Hyperosmotic stress stimulates phospholipase D activity and elevates the levels of phosphatidic acid and diacylglycerol pyrophosphate. Plant J. 2000, 22, 147–154. [Google Scholar] [CrossRef]
- van Schooten, B.; Testerink, C.; Munnik, T. Signalling diacylglycerol pyrophosphate, a new phosphatidic acid metabolite. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2006, 1761, 151–159. [Google Scholar] [CrossRef]
- Putta, P.; Creque, E.; Piontkivska, H.; Kooijman, E.E. Lipid-protein interactions for ECA1 an N-ANTH domain protein involved in stress signaling in plants. Chem. Phys. Lipids 2020, 231, 104919. [Google Scholar] [CrossRef]
- Strawn, L.; Babb, A.; Testerink, C.; Kooijman, E.E. The physical chemistry of the enigmatic phospholipid diacylglycerol pyrophosphate. Front. Plant Sci. 2012, 3, 40. [Google Scholar] [CrossRef]
- Kooijman, E.E.; Tieleman, D.P.; Testerink, C.; Munnik, T.; Rijkers, D.T.S.; Burger, K.N.J.; De Kruijff, B. An electrostatic/hydrogen bond switch as the basis for the specific interaction of phosphatidic acid with proteins. J. Biol. Chem. 2007, 282, 11356–11364. [Google Scholar] [CrossRef] [PubMed]
- Peppino Margutti, M.; Wilke, N.; Villasuso, A.L. Influence of Ca2+ on the surface behavior of phosphatidic acid and its mixture with diacylglycerol pyrophosphate at different pHs. Chem. Phys. Lipids 2020, 228, 104887. [Google Scholar] [CrossRef]
- Villasuso, A.L.; Wilke, N.; Maggio, B.; Machado, E. Zn2+-dependent surface behavior of diacylglycerol pyrophosphate and its mixtures with phosphatidic acid at different pHs. Front. Plant Sci. 2014, 5, 104887. [Google Scholar] [CrossRef]
- Liu, A.; Wenzel, N.; Qi, X. Role of lysine residues in membrane anchoring of saposin C. Arch. Biochem. Biophys. 2005, 443, 101–112. [Google Scholar] [CrossRef]
- Li, L.; Vorobyov, I.; Allen, T.W. The different interactions of lysine and arginine side chains with lipid membranes. J. Phys. Chem. B 2013, 117, 11906–11920. [Google Scholar] [CrossRef]
- Stahelin, R.V. Lipid binding domains: More than simple lipid effectors. J. Lipid Res. 2009, 50, S299–S304. [Google Scholar] [CrossRef] [PubMed]
- Loew, S.; Kooijman, E.E.; May, S. Increased pH-sensitivity of protein binding to lipid membranes through the electrostatic-hydrogen bond switch. Chem. Phys. Lipids 2013, 169, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, D.H.; Kooijman, E.E.; May, S. Ionization properties of mixed lipid membranes: A Gouy-Chapman model of the electrostatic-hydrogen bond switch. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 1985–1992. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Lee, C.S.; Hwang, D.; Ryu, S.H. Understanding of the roles of phospholipase D and phosphatidic acid through their binding partners. Prog. Lipid Res. 2012, 51, 71–81. [Google Scholar] [CrossRef]
- Zegarlińska, J.; Piaścik, M.; Sikorski, A.F.; Czogalla, A. Phosphatidic acid-A simple phospholipid with multiple faces. Acta Biochim. Pol. 2018, 65, 163–171. [Google Scholar] [CrossRef]
- Tanguy, E.; Kassas, N.; Vitale, N. Protein–phospholipid interaction motifs: A focus on phosphatidic acid. Biomolecules 2018, 8, 20. [Google Scholar] [CrossRef]
- Stahelin, R.V.; Scott, J.L.; Frick, C.T. Cellular and molecular interactions of phosphoinositides and peripheral proteins. Chem. Phys. Lipids 2014, 182, 3–18. [Google Scholar] [CrossRef]
- Dowler, S.; Currie, R.A.; Campbell, D.G.; Deak, M.; Kular, G.; Downes, C.P.; Alessi, D.R. Identification of pleckstrin-homology-domain-containing proteins with novel phosphoinositide-binding specificities. Biochem. J. 2000, 351, 19–31. [Google Scholar] [CrossRef]
- Astorquiza, P.L.; Usorach, J.; Racagni, G.; Villasuso, A.L. Diacylglycerol pyrophosphate binds and inhibits the glyceraldehyde-3-phosphate dehydrogenase in barley aleurone. Plant Physiol. Biochem. 2016, 101, 88–95. [Google Scholar] [CrossRef]
- Sirover, M.A. Structural analysis of glyceraldehyde-3-phosphate dehydrogenase functional diversity. Int. J. Biochem. Cell Biol. 2014, 57, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Camacho, P.; Lechleiter, J.D.; Herman, B. Measurement of intracellular calcium. Physiol. Rev. 1999, 79, 1089–1125. [Google Scholar] [CrossRef]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. BioMetals 2002, 15, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium signaling in plants. Int. J. Mol. Sci. 2021, 22, 1159. [Google Scholar] [CrossRef]
- Papahadjopoulos, D.; Vail, W.J.; Newton, C.; Nir, S.; Jacobson, K.; Poste, G.; Lazo, R. Studies on membrane fusion. III. The role of calcium-induced phase changes. Biochim. et Biophys. Acta (BBA)-Biomembr. 1977, 465, 579–598. [Google Scholar] [CrossRef]
- Binder, H.; Zschörnig, O. The effect of metal cations on the phase behavior and hydration characteristics of phospholipid membranes. Chem. Phys. Lipids 2002, 115, 39–61. [Google Scholar] [CrossRef]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal. Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef]
- Tang, D.; Dean, W.L.; Borchman, D.; Paterson, C.A. The influence of membrane lipid structure on plasma membrane Ca2+-ATPase activity. Cell Calcium 2006, 39, 209–216. [Google Scholar] [CrossRef]
- Garidel, P.; Blume, A.; Hübner, W. A Fourier transform infrared spectroscopic study of the interaction of alkaline earth cations with the negatively charged phospholipid 1,2-dimyristoyl-sn-glycero-3-phosphoglycerol. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1466, 245–259. [Google Scholar] [CrossRef]
- Tsai, H.H.G.; Lai, W.X.; Lin, H.D.; Lee, J.B.; Juang, W.F.; Tseng, W.H. Molecular dynamics simulation of cation-phospholipid clustering in phospholipid bilayers: Possible role in stalk formation during membrane fusion. Biochim. Biophys. Acta-Biomembr. 2012, 1818, 2742–2755. [Google Scholar] [CrossRef]
- Wang, Y.H.; Collins, A.; Guo, L.; Smith-Dupont, K.B.; Gai, F.; Svitkina, T.; Janmey, P.A. Divalent cation-induced cluster formation by polyphosphoinositides in model membranes. J. Am. Chem. Soc. 2012, 134, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Düzgüne, N.; Nir, S.; Wilschut, J.; Bentz, J.; Newton, C.; Portis, A.; Papahadjopoulos, D. Calcium- and magnesium-induced fusion of mixed phosphatidylserine/phosphatidylcholine vesicles: Effect of ion binding. J. Membr. Biol. 1981, 59, 115–125. [Google Scholar] [CrossRef]
- Martín-Molina, A.; Rodríguez-Beas, C.; Faraudo, J. Effect of calcium and magnesium on phosphatidylserine membranes: Experiments and all-atomic simulations. Biophys. J. 2012, 102, 2095–2103. [Google Scholar] [CrossRef]
- Graber, Z.T.; Gericke, A.; Kooijman, E.E. Phosphatidylinositol-4,5-bisphosphate ionization in the presence of cholesterol, calcium or magnesium ions. Chem. Phys. Lipids 2014, 182, 62–72. [Google Scholar] [CrossRef] [PubMed]
- De Planque, M.R.R.; Kruijtzer, J.A.W.; Liskamp, R.M.J.; Marsh, D.; Greathouse, D.V.; Koeppe, R.E.; De Kruijff, B.; Killian, J.A. Different membrane anchoring positions of tryptophan and lysine in synthetic transmembrane α-helical peptides. J. Biol. Chem. 1999, 274, 20839–20846. [Google Scholar] [CrossRef]
- De Planque, M.R.R.; Goormaghtigh, E.; Greathouse, D.V.; Koeppe, R.E.; Kruijtzer, J.A.W.; Liskamp, R.M.J.; De Kruijff, B.; Killian, J.A. Sensitivity of single membrane-spanning α-helical peptides to hydrophobic mismatch with a lipid bilayer: Effects on backbone structure, orientation, and extent of membrane incorporation. Biochemistry 2001, 40, 5000–5010. [Google Scholar] [CrossRef]
- Strandberg, E.; Morein, S.; Rijkers, D.T.S.; Liskamp, R.M.J.; Van der Wel, P.C.A.; Antoinette Killian, J. Lipid dependence of membrane anchoring properties and snorkeling behavior of aromatic and charged residues in transmembrane peptides. Biochemistry 2002, 41, 7190–7198. [Google Scholar] [CrossRef]
- Rouser, G.; Fleischer, S.; Yamamoto, A. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Graber, Z.T.; Kooijman, E.E. Ionization behavior of polyphosphoinositides determined via the preparation of pH titration curves using solid-state 31P NMR. Methods Mol. Biol. 2013, 1009, 129–142. [Google Scholar] [CrossRef]
- Kooijman, E.E.; Carter, K.M.; Van Laar, E.G.; Chupin, V.; Burger, K.N.J.; De Kruijff, B. What makes the bioactive lipids phosphatidic acid and lysophosphatidic acid so special? Biochemistry 2005, 44, 17007–17015. [Google Scholar] [CrossRef]
- Appleton, T.G.; Hall, J.R.; Ralph, S.F.; Thompson, C.S.M. NMR study of acid-base equilibria and other reactions of ammineplatinum complexes with aqua and hydroxo ligands. Inorg. Chem. 1989, 28, 1989–1993. [Google Scholar] [CrossRef]
- De Planque, M.R.R.; Killian, J.A. Protein-lipid interactions studied with designed transmembrane peptides: Role of hydrophobic matching and interfacial anchoring (Review). Mol. Membr. Biol. 2003, 20, 271–284. [Google Scholar] [CrossRef]
- Lu, J.X.; Damodaraiv, K.; Blazyk, J.; Lorigan, G.A. Solid-state nuclear magnetic resonance relaxation studies of the interaction mechanism of antimicrobial peptides with phospholipid bilayer membranes. Biochemistry 2005, 44, 10208–10217. [Google Scholar] [CrossRef]
- Dufourc, E.J.; Mayer, C.; Stohrer, J.; Althoff, G.; Kothe, G. Dynamics of phosphate head groups in biomembranes. Comprehensive analysis using phosphorus-31 nuclear magnetic resonance lineshape and relaxation time measurements. Biophys. J. 1992, 61, 42–57. [Google Scholar] [CrossRef]
- Cullis, P.R.; De Kruyff, B.; Richards, R.E. Factors affecting the motion of the polar headgroup in phospholipid bilayers. A 31P NMR study of unsonicated phosphatidylcholine liposomes. BBA-Biomembr. 1976, 426, 433–446. [Google Scholar] [CrossRef]
- Kooijman, E.E.; King, K.E.; Gangoda, M.; Gericke, A. Ionization Properties of Phosphatidylinositol Polyphosphates in Mixed Model Membranes. Biochemistry 2009, 48, 9360–9371. [Google Scholar] [CrossRef] [PubMed]
- Putta, P.; Rankenberg, J.; Korver, R.A.; van Wijk, R.; Munnik, T.; Testerink, C.; Kooijman, E.E. Phosphatidic acid binding proteins display differential binding as a function of membrane curvature stress and chemical properties. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 2709–2716. [Google Scholar] [CrossRef]
- Graber, Z.T.; Wang, W.; Singh, G.; Kuzmenko, I.; Vaknin, D.; Kooijman, E.E. Competitive cation binding to phosphatidylinositol-4,5-bisphosphate domains revealed by X-ray fluorescence. RSC Adv. 2015, 5, 106536–106542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Composition | pKa2 a | Phosphomonoester Charge at pH 7.2 | Total Charge at pH 7.2 |
|---|---|---|---|
| (±0.1) b | (±0.1) b | ||
| DOPC/DGPP 95:5 | 7.44 ± 0.02 | −1.38 | −1.38 |
| DOPC/DGPP/KALP23 95:5:4 | 6.38 ± 0.04 | −1.84 | −2.84 |
| DOPC/DGPP/DOTAP 95:5:16 | 7.43 ± 0.03 | −1.37 | −2.37 |
| DOPC/DGPP/EtPC 95:5:16 | 7.25 ± 0.01 | −1.47 | −2.47 |
| DOPC/DGPP/EtPC 95:5:32 | 7.14 ± 0.02 | −1.53 | −2.53 |
| Cation Content | Phosphomonoester Charge ± 0.1 at pH 7.20 |
|---|---|
| 0 mM | −1.40 |
| 0.01 mM Ca2+ | −1.41 |
| 0.1 mM Ca2+ | −1.55 |
| 1 mM Ca2+ | −1.88 |
| 1 mM Mg2+ | −1.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graber, Z.; Owusu Kwarteng, D.; Lange, S.M.; Koukounas, Y.; Khalifa, H.; Mutambuze, J.W.; Kooijman, E.E. The Electrostatic Basis of Diacylglycerol Pyrophosphate—Protein Interaction. Cells 2022, 11, 290. https://doi.org/10.3390/cells11020290
Graber Z, Owusu Kwarteng D, Lange SM, Koukounas Y, Khalifa H, Mutambuze JW, Kooijman EE. The Electrostatic Basis of Diacylglycerol Pyrophosphate—Protein Interaction. Cells. 2022; 11(2):290. https://doi.org/10.3390/cells11020290
Chicago/Turabian StyleGraber, Zachary, Desmond Owusu Kwarteng, Shannon M. Lange, Yannis Koukounas, Hady Khalifa, Jean W. Mutambuze, and Edgar E. Kooijman. 2022. "The Electrostatic Basis of Diacylglycerol Pyrophosphate—Protein Interaction" Cells 11, no. 2: 290. https://doi.org/10.3390/cells11020290