
Isoalantolactone (IAL) Regulates Neuro-Inflammation and Neuronal Apoptosis to Curb Pathology of Parkinson’s Disease
Abstract
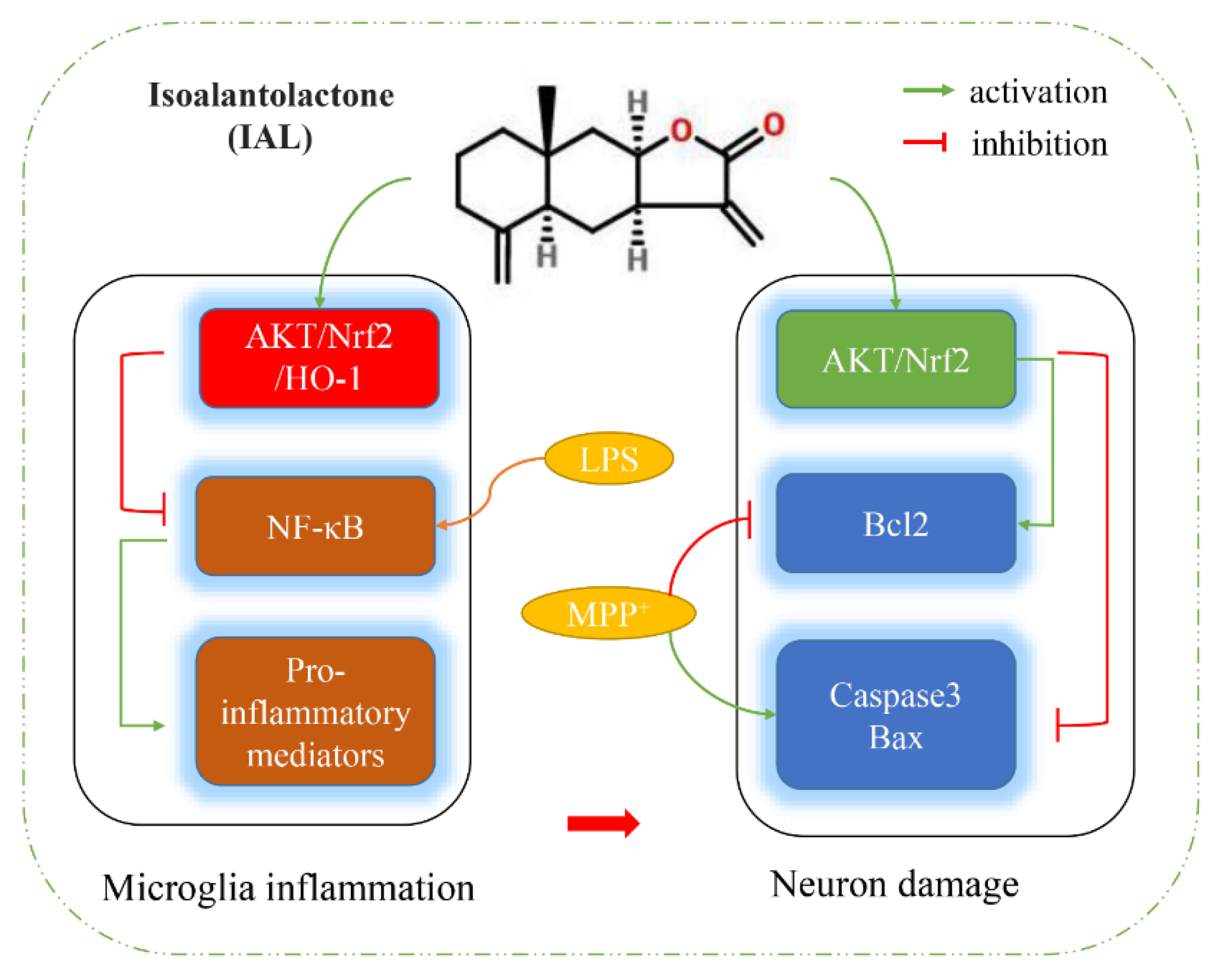
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals and Models
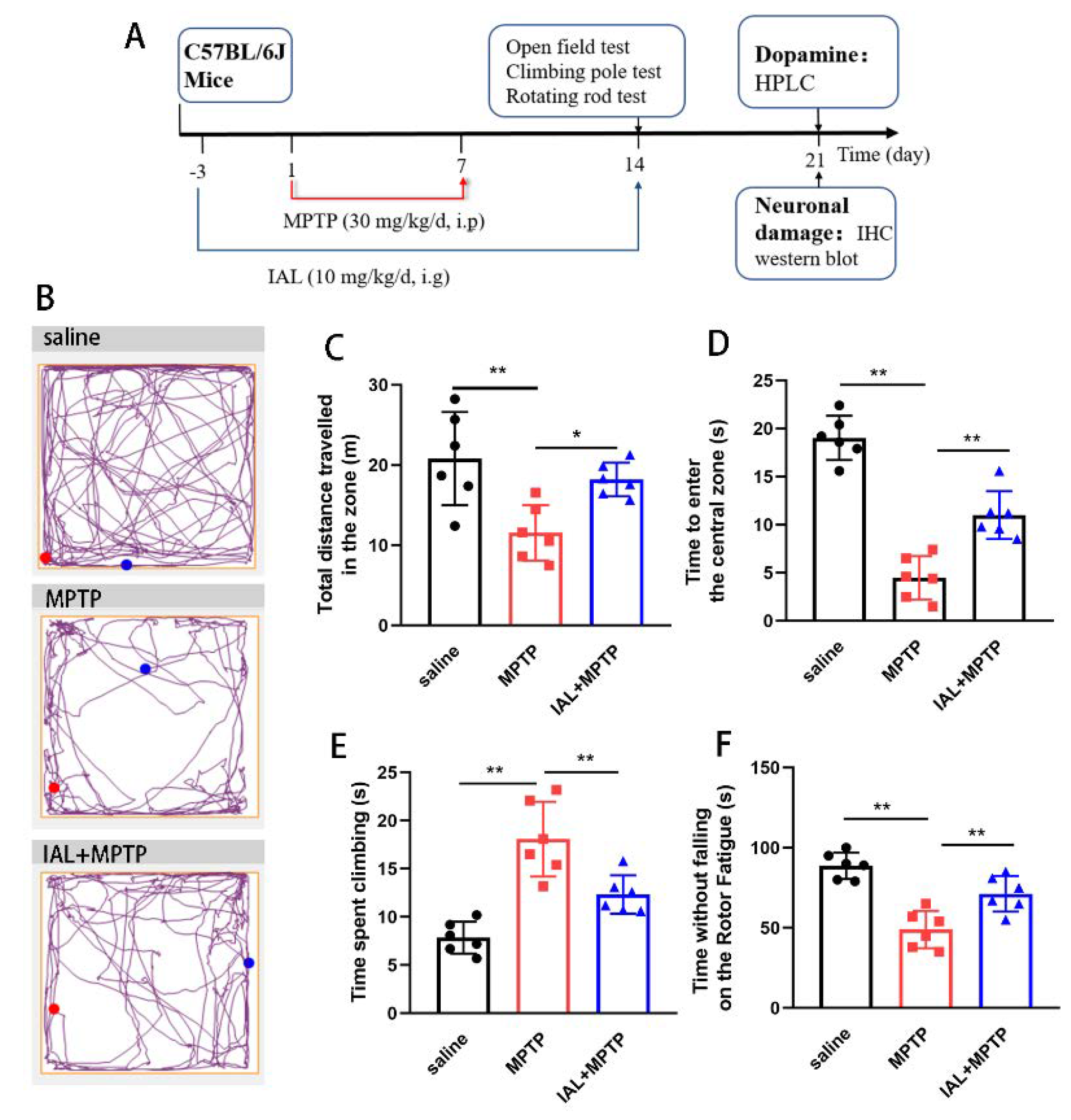
2.3. Behavioral Testing
2.3.1. Open Field Test
2.3.2. Climbing Pole Test
2.3.3. Rotating Rod Test
2.4. Immunohistochemistry (IHC) Staining
2.5. High-Performance Liquid Chromatography (HPLC) Assay
2.6. Cell Culture
2.7. CCK-8 Assay
2.8. ELISA Assay
2.9. LDH Assay
2.10. RT-PCR Assay
2.11. Western Blot Assay
2.12. Flow Cytometry
2.13. Tunel Staining
2.14. Data Analyses
3. Results
3.1. IAL Treatment Alleviates the Mobility of MPTP-Induced PD Mice
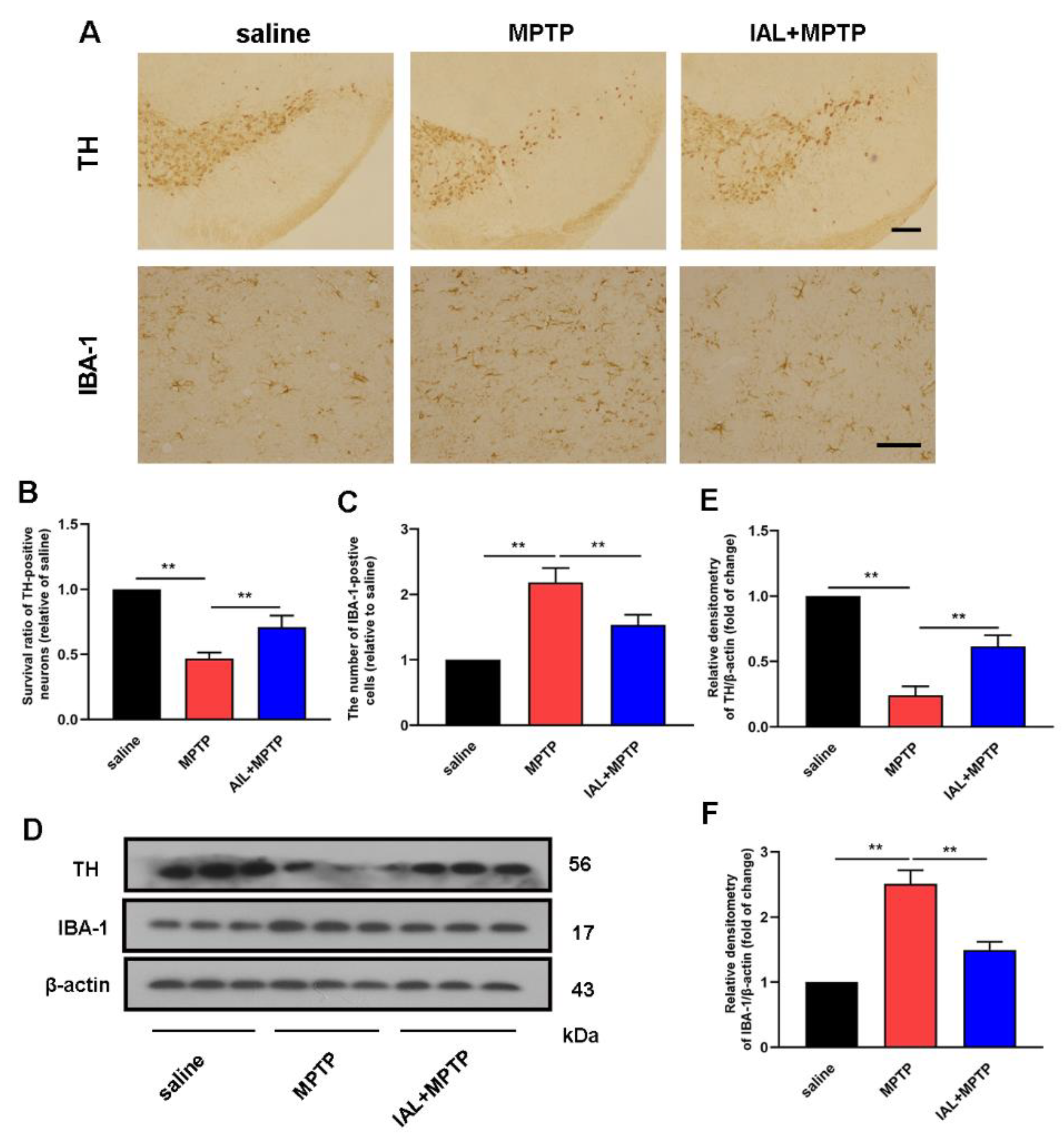
3.2. IAL Treatment Decreases Dopaminergic Neuron Degeneration and Inhibits Microglia Over-Activation in MPTP-Induced PD Mice
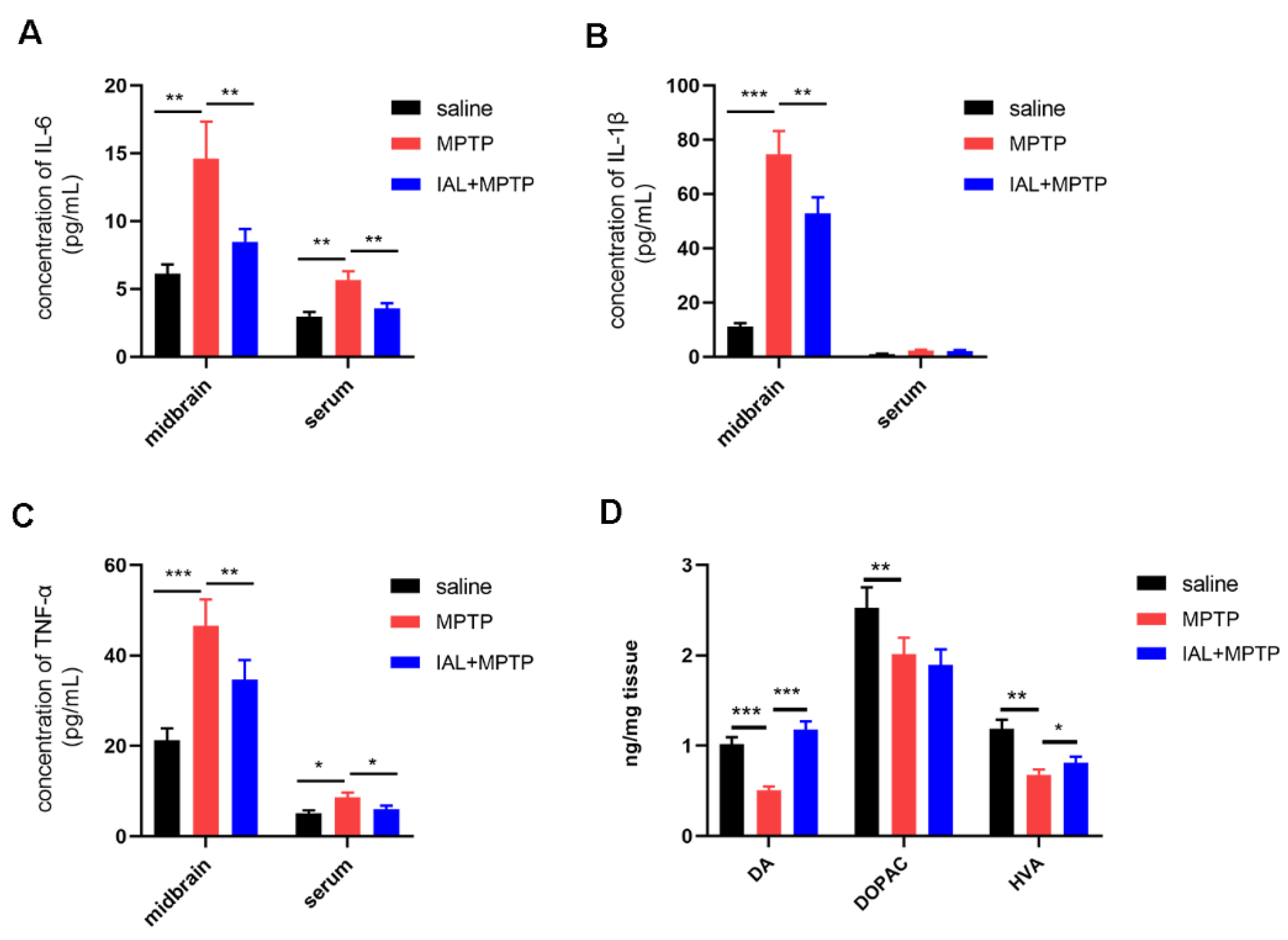
3.3. IAL Treatment Suppresses Midbrain Inflammatory Responses and Increases Striatal Dopamine and Its Metabolite Levels in MPTP-Induced PD Mice
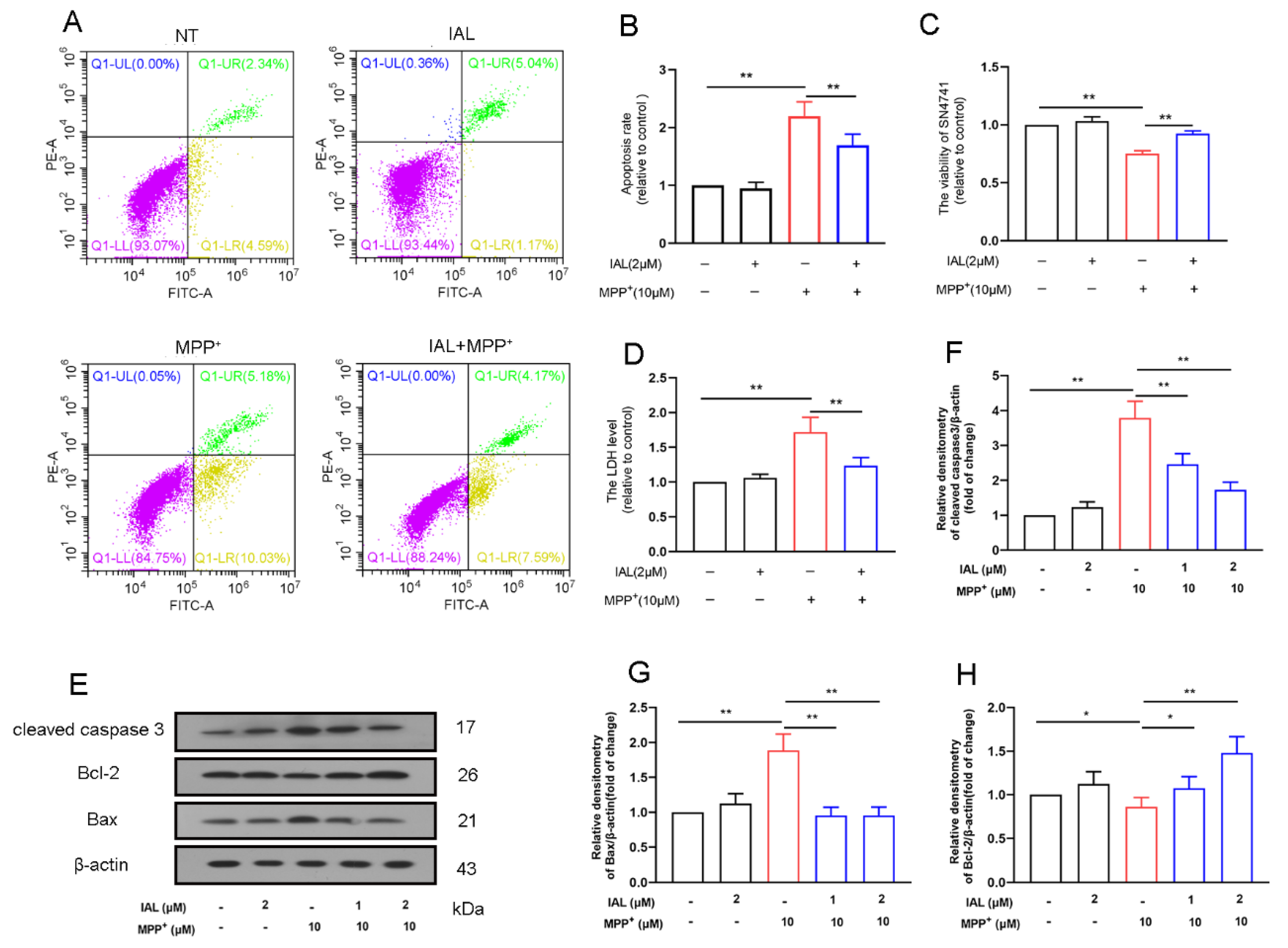
3.4. IAL Treatment Reduces Cells Death in MPP+-Exposed SN4741 Cells
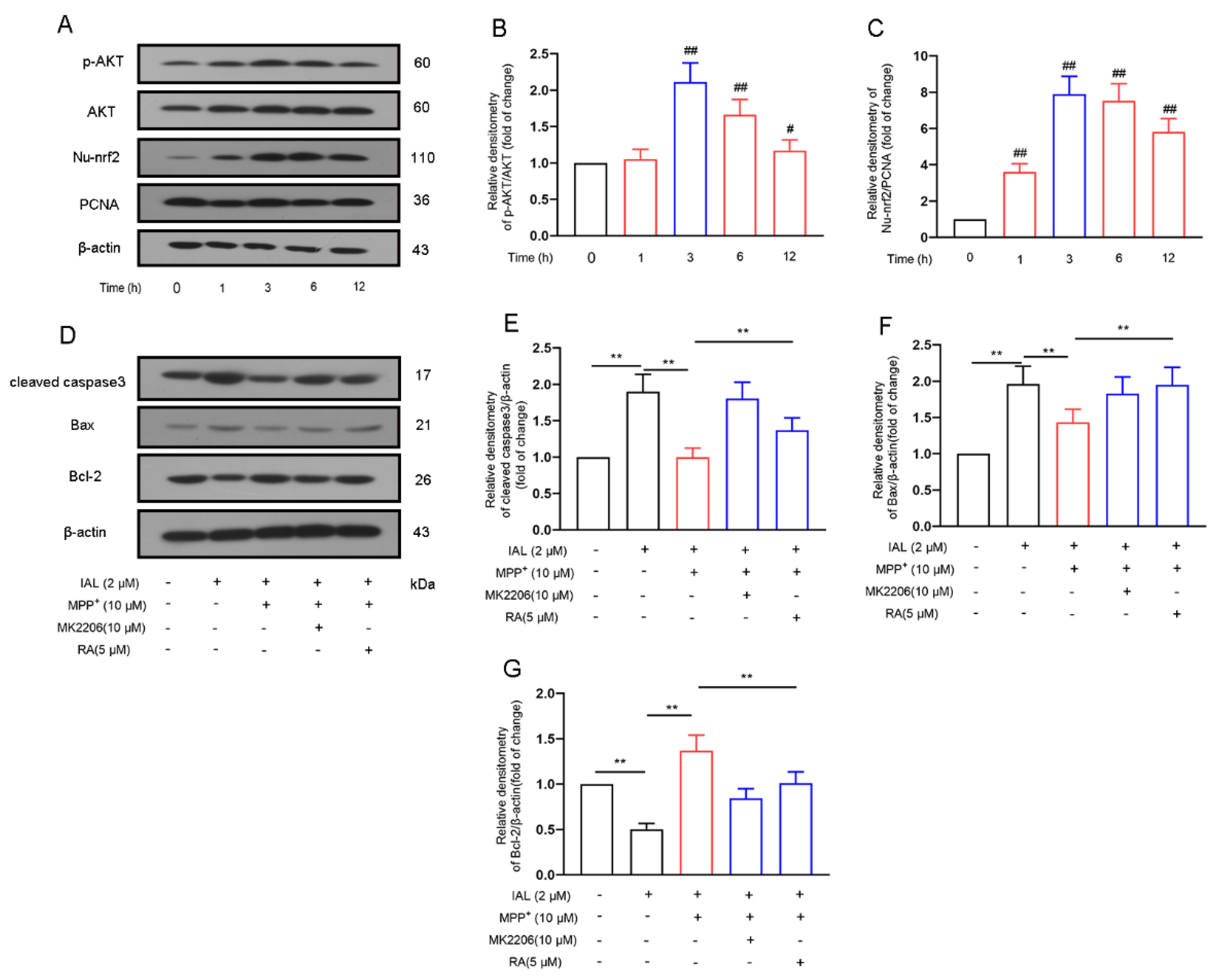
3.5. IAL Treatment Reduces MPP+-Induced Apoptosis of SN4741 Cells by Activating the AKT/Nrf2 Signaling Pathway
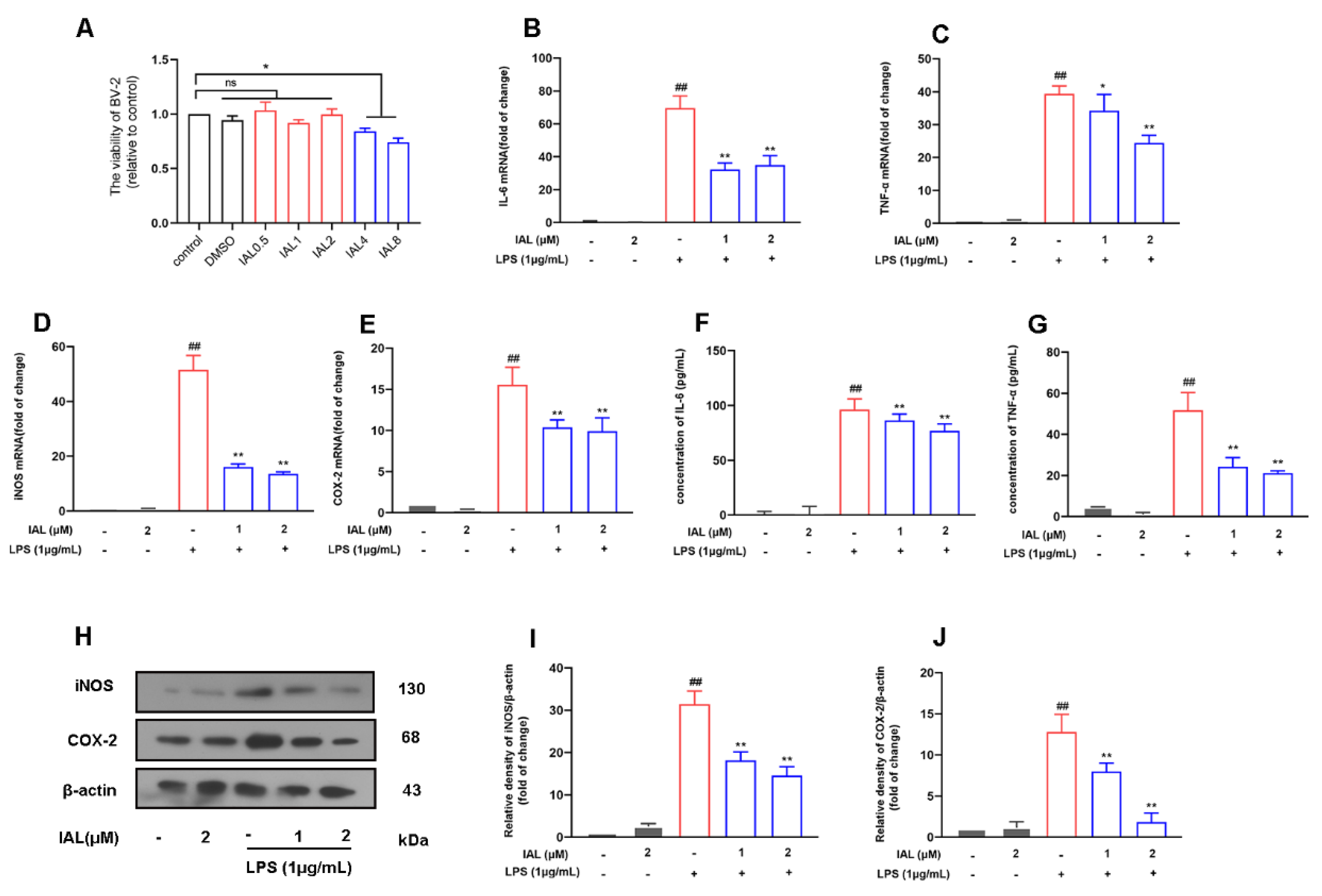
3.6. IAL Treatment Suppresses the Release of Pro-Inflammatory Mediators in LPS-Induced BV2 Cells
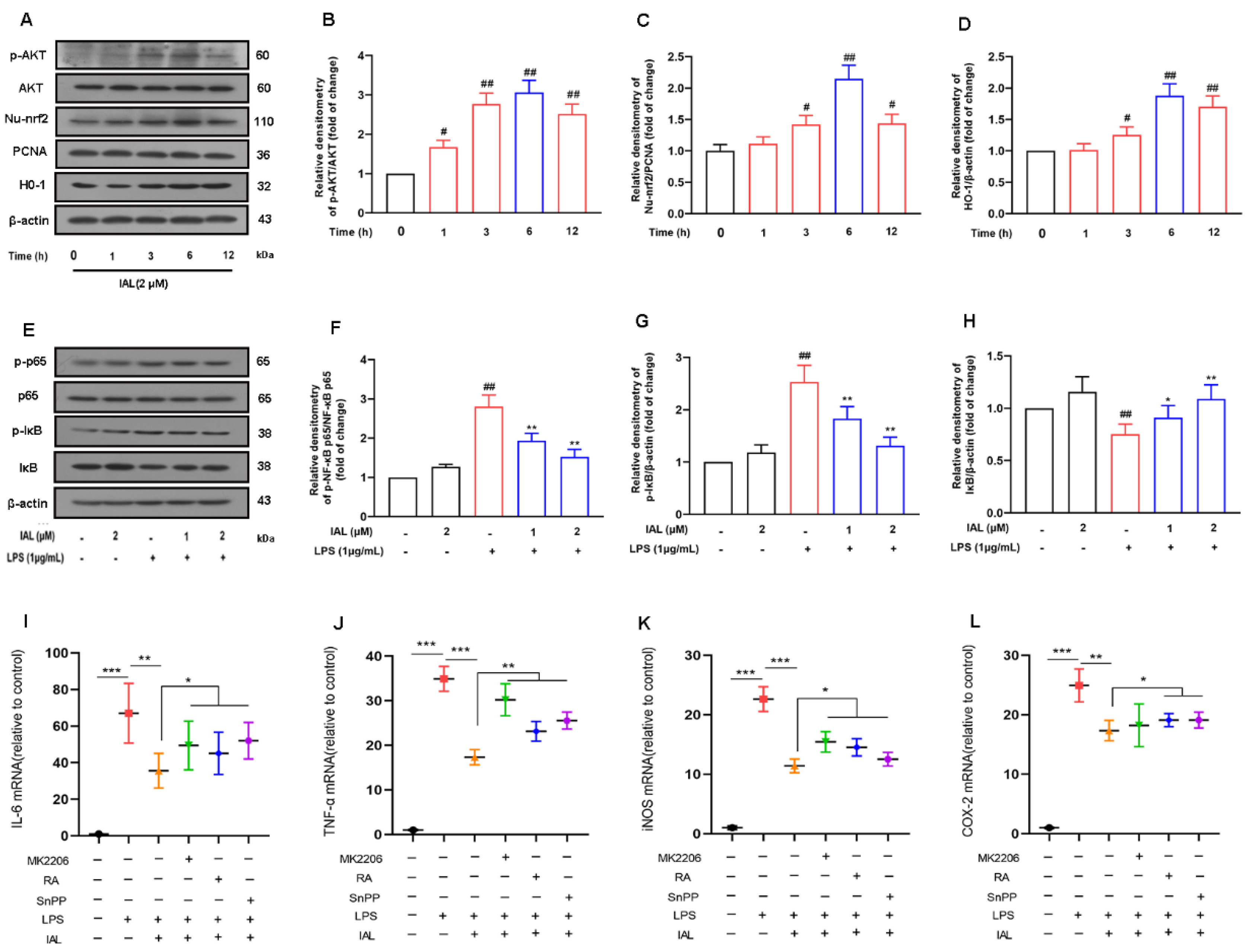
3.7. IAL Treatment Inhibits Neuro-Inflammation by Activating the AKT/Nrf2/HO-1 Pathway and Suppressing the NF-κB Signaling Pathway
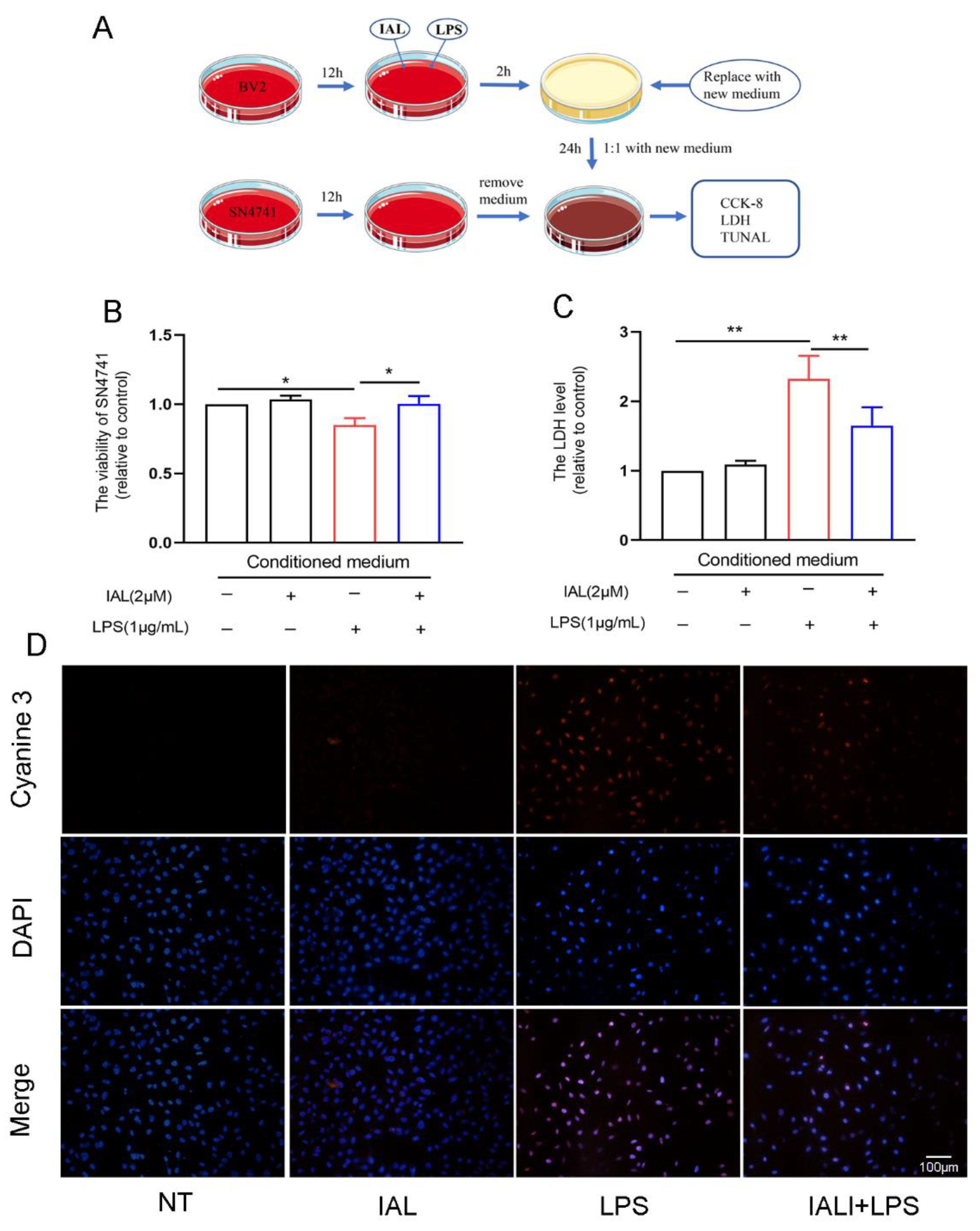
3.8. IAL Treatment Protects SN4741 from Microglial Activation-Mediated Neurotoxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nagatsu, T.; Nakashima, A.; Ichinose, H.; Kobayashi, K. Human tyrosine hydroxylase in Parkinson’s disease and in related disorders. J. Neural Transm. 2019, 126, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T.; Sawada, M. Biochemistry of postmortem brains in Parkinson’s disease: Historical overview and future prospects. J. Neural Transm. Suppl. 2007, 72, 113–120. [Google Scholar] [CrossRef]
- Christine, C.W.; Aminoff, M.J. Clinical differentiation of parkinsonian syndromes: Prognostic and therapeutic relevance. Am. J. Med. 2004, 117, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Mackie, P.; Lebowitz, J.; Saadatpour, L.; Nickoloff, E.; Gaskill, P.; Khoshbouei, H. The dopamine transporter: An unrecognized nexus for dysfunctional peripheral immunity and signaling in Parkinson’s Disease. Brain Behav. Immun. 2018, 70, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Mauerer, J.; Schlaier, J.; Janzen, A.; Zeman, F.; Bogdahn, U.; Brawanski, A.; Hochreiter, A. Underutilization of deep brain stimulation for Parkinson’s disease? A survey on possible clinical reasons. Acta Neurochir. 2017, 159, 771–778. [Google Scholar] [CrossRef]
- Zhang, S.; Tao, K.; Wang, J.; Duan, Y.; Wang, B.; Liu, X. Substantia Nigra Hyperechogenicity Reflects the Progression of Dopaminergic Neurodegeneration in 6-OHDA Rat Model of Parkinson’s Disease. Front. Cell. Neurosci. 2020, 14, 216. [Google Scholar] [CrossRef]
- Grilo, A.L.; Mantalaris, A. Apoptosis: A mammalian cell bioprocessing perspective. Biotechnol. Adv. 2019, 37, 459–475. [Google Scholar] [CrossRef]
- Henson, P.M. Cell Removal: Efferocytosis. Annu. Rev. Cell Dev. Biol. 2017, 33, 127–144. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis and Clearance of Apoptotic Cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef]
- Bandres-Ciga, S.; Saez-Atienzar, S.; Kim, J.J.; Makarious, M.B.; Faghri, F.; Diez-Fairen, M.; Iwaki, H.; Leonard, H.; Botia, J.; Ryten, M.; et al. Large-scale pathway specific polygenic risk and transcriptomic community network analysis identifies novel functional pathways in Parkinson disease. Acta Neuropathol. 2020, 140, 341–358. [Google Scholar] [CrossRef]
- Dionisio, P.A.; Amaral, J.D.; Rodrigues, C.M.P. Oxidative stress and regulated cell death in Parkinson’s disease. Ageing Res. Rev. 2021, 67, 101263. [Google Scholar] [CrossRef] [PubMed]
- Zolezzi, J.M.; Santos, M.J.; Bastias-Candia, S.; Pinto, C.; Godoy, J.A.; Inestrosa, N.C. PPARs in the central nervous system: Roles in neurodegeneration and neuroinflammation. Biol. Rev. Camb. Philos. Soc. 2017, 92, 2046–2069. [Google Scholar] [CrossRef] [PubMed]
- Kalola, J.; Shah, R.; Patel, A.; Lahiri, S.K.; Shah, M.B. Anti-inflammatory and immunomodulatory activities of Inula cappa roots (Compositae). J. Complement. Integr. Med. 2017, 14, 20160083. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.Y.; Park, J.; Kim, H.J.; Lee, I.A.; Lim, J.S.; Lim, S.S.; Choi, S.J.; Park, J.H.; Kang, H.J.; Kim, J.S. Isoalantolactone from Inula helenium caused Nrf2-mediated induction of detoxifying enzymes. J. Med. Food 2009, 12, 1038–1045. [Google Scholar] [CrossRef]
- Song, Y.; Li, X.; Liu, F.; Zhu, H.; Shen, Y. Isoalantolactone alleviates ovalbumininduced asthmatic inflammation by reducing alternatively activated macrophage and STAT6/PPARgamma/KLF4 signals. Mol. Med. Rep. 2021, 24, 1791–2997. [Google Scholar] [CrossRef]
- He, G.; Zhang, X.; Chen, Y.; Chen, J.; Li, L.; Xie, Y. Isoalantolactone inhibits LPS-induced inflammation via NF-kappaB inactivation in peritoneal macrophages and improves survival in sepsis. Biomed. Pharmacother. 2017, 90, 598–607. [Google Scholar] [CrossRef]
- Ding, Y.H.; Song, Y.D.; Wu, Y.X.; He, H.Q.; Yu, T.H.; Hu, Y.D.; Zhang, D.P.; Jiang, H.C.; Yu, K.K.; Li, X.Z.; et al. Isoalantolactone suppresses LPS-induced inflammation by inhibiting TRAF6 ubiquitination and alleviates acute lung injury. Acta Pharmacol. Sin. 2019, 40, 64–74. [Google Scholar] [CrossRef]
- Yuan, C.B.; Tian, L.; Yang, B.; Zhou, H.Y. Isoalantolactone protects LPS-induced acute lung injury through Nrf2 activation. Microb. Pathog. 2018, 123, 213–218. [Google Scholar] [CrossRef]
- Seo, J.Y.; Lim, S.S.; Kim, J.; Lee, K.W.; Kim, J.S. Alantolactone and Isoalantolactone Prevent Amyloid beta25-35 -induced Toxicity in Mouse Cortical Neurons and Scopolamine-induced Cognitive Impairment in Mice. Phytother. Res. 2017, 31, 801–811. [Google Scholar] [CrossRef]
- Wang, M.; Wang, K.; Gao, X.; Zhao, K.; Chen, H.; Xu, M. Anti-inflammatory effects of isoalantolactone on LPS-stimulated BV2 microglia cells through activating GSK-3beta-Nrf2 signaling pathway. Int. Immunopharmacol. 2018, 65, 323–327. [Google Scholar] [CrossRef]
- Jiang, P.E.; Lang, Q.H.; Yu, Q.Y.; Tang, X.Y.; Liu, Q.Q.; Li, X.Y.; Feng, X.Z. Behavioral Assessments of Spontaneous Locomotion in a Murine MPTP-induced Parkinson’s Disease Model. J. Vis. Exp. 2019, 163, 58653. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Hu, G.; Zhou, A.; Liu, Y.; Huang, B.; Su, Y.; Wang, H.; Ye, B.; He, Y.; Gao, X.; et al. Echinocystic Acid Inhibits Inflammation and Exerts Neuroprotective Effects in MPTP-Induced Parkinson’s Disease Model Mice. Front. Pharmacol. 2021, 12, 787771. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.E.; Ahn, S.M.; Jung, D.H.; Lee, H.J.; Ha, K.T.; Shin, H.K.; Choi, B.T. Electroacupuncture Therapy Ameliorates Motor Dysfunction via Brain-Derived Neurotrophic Factor and Glial Cell Line-Derived Neurotrophic Factor in a Mouse Model of Parkinson’s Disease. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Xie, J.; Li, X.; Li, Y.; Thirupathi, A.; Zhang, J.; Yu, P.; Gao, G.; Chang, Y.; Shi, Z. Ferritinophagy-Mediated Ferroptosis Involved in Paraquat-Induced Neurotoxicity of Dopaminergic Neurons: Implication for Neurotoxicity in PD. Oxid. Med. Cell. Longev. 2021, 2021, 9961628. [Google Scholar] [CrossRef]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- Hernandez-Baltazar, D.; Nadella, R.; Mireya Zavala-Flores, L.; Rosas-Jarquin, C.J.; Rovirosa-Hernandez, M.J.; Villanueva-Olivo, A. Four main therapeutic keys for Parkinson’s disease: A mini review. Iran J. Basic Med. Sci. 2019, 22, 716–721. [Google Scholar] [CrossRef]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Chiba, K.; Trevor, A.; Castagnoli, N., Jr. Metabolism of the neurotoxic tertiary amine, MPTP, by brain monoamine oxidase. Biochem. Biophys. Res. Commun. 1984, 120, 574–578. [Google Scholar] [CrossRef]
- Agim, Z.S.; Cannon, J.R. Dietary factors in the etiology of Parkinson’s disease. Biomed. Res. Int. 2015, 2015, 672838. [Google Scholar] [CrossRef]
- Kurzawski, M.; Bialecka, M.; Drozdzik, M. Pharmacogenetic considerations in the treatment of Parkinson’s disease. Neurodegener. Dis. Manag. 2015, 5, 27–35. [Google Scholar] [CrossRef]
- Forloni, G.; La Vitola, P.; Cerovic, M.; Balducci, C. Inflammation and Parkinson’s disease pathogenesis: Mechanisms and therapeutic insight. Prog. Mol. Biol. Transl. Sci. 2021, 177, 175–202. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.; Manda, G.; Bosca, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Hou, L.; Jing, L.; Ruan, Z.; Peng, B.; Zhang, X.; Hong, J.S.; Zhao, J.; Wang, Q. Microglial activation contributes to cognitive impairments in rotenone-induced mouse Parkinson’s disease model. J. Neuroinflammation 2021, 18, 4. [Google Scholar] [CrossRef]
- Chinta, S.J.; Andersen, J.K. Dopaminergic neurons. Int. J. Biochem. Cell Biol. 2005, 37, 942–946. [Google Scholar] [CrossRef]
- Tsutsumi, R.; Hori, Y.; Seki, T.; Kurauchi, Y.; Sato, M.; Oshima, M.; Hisatsune, A.; Katsuki, H. Involvement of exosomes in dopaminergic neurodegeneration by microglial activation in midbrain slice cultures. Biochem. Biophys. Res. Commun. 2019, 511, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zeng, W.; Tao, K.; Lu, F.; Gao, G.; Yang, Q. Myricitrin alleviates MPP(+)-induced mitochondrial dysfunction in a DJ-1-dependent manner in SN4741 cells. Biochem. Biophys. Res. Commun. 2015, 458, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Yoo, M.S.; Chun, H.S.; Son, J.J.; DeGiorgio, L.A.; Kim, D.J.; Peng, C.; Son, J.H. Oxidative stress regulated genes in nigral dopaminergic neuronal cells: Correlation with the known pathology in Parkinson’s disease. Mol. Brain Res. 2003, 110, 76–84. [Google Scholar] [CrossRef]
- Bahdoudi, S.; Ghouili, I.; Hmiden, M.; do Rego, J.L.; Lefranc, B.; Leprince, J.; Chuquet, J.; do Rego, J.C.; Marcher, A.B.; Mandrup, S.; et al. Neuroprotective effects of the gliopeptide ODN in an in vivo model of Parkinson’s disease. Cell. Mol. Life Sci. 2018, 75, 2075–2091. [Google Scholar] [CrossRef]
- Lim, H.S.; Moon, B.C.; Lee, J.; Choi, G.; Park, G. The insect molting hormone 20-hydroxyecdysone protects dopaminergic neurons against MPTP-induced neurotoxicity in a mouse model of Parkinson’s disease. Free Radic. Biol. Med. 2020, 159, 23–36. [Google Scholar] [CrossRef]
- Deng, Q.; Yang, X. Protective effects of Gynostemma pentaphyllum polysaccharides on PC12 cells impaired by MPP(+). Int. J. Biol. Macromol. 2014, 69, 171–175. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Y.; Zhu, Z.; Mo, L.; Lin, C.; Wang, Q.; Wang, H.; Gong, X.; He, X.; Lu, G.; et al. MiR-124 Regulates Apoptosis and Autophagy Process in MPTP Model of Parkinson’s Disease by Targeting to Bim. Brain Pathol. 2016, 26, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.K. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic. Biol. Med. 2004, 36, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Luo, X.; Wu, X.; Zhang, T.; Gu, L.; Wang, Y.; Gao, M.; Cheng, Y.; Xie, Z. Activation of the Melanocortin-1 Receptor by NDP-MSH Attenuates Oxidative Stress and Neuronal Apoptosis through PI3K/Akt/Nrf2 Pathway after Intracerebral Hemorrhage in Mice. Oxid. Med. Cell. Longev. 2020, 2020, 8864100. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Zhang, W.; Wu, Z.; Tian, X.; Xiang, J.; Li, L.; Li, Z.; Peng, X.; Wei, S.; Ma, X.; et al. Baicalin and the liver-gut system: Pharmacological bases explaining its therapeutic effects. Pharmacol. Res. 2021, 165, 105444. [Google Scholar] [CrossRef]
- Monzani, E.; Nicolis, S.; Dell’Acqua, S.; Capucciati, A.; Bacchella, C.; Zucca, F.A.; Mosharov, E.V.; Sulzer, D.; Zecca, L.; Casella, L. Dopamine, Oxidative Stress and Protein-Quinone Modifications in Parkinson’s and Other Neurodegenerative Diseases. Angew. Chem. Int. Ed. Engl. 2019, 58, 6512–6527. [Google Scholar] [CrossRef]
- Chen, F.; Shi, X. NF-kappaB, a pivotal transcription factor in silica-induced diseases. Mol. Cell. Biochem. 2002, 234–235, 169–176. [Google Scholar] [CrossRef]
- Sivandzade, F.; Prasad, S.; Bhalerao, A.; Cucullo, L. NRF2 and NF-B interplay in cerebrovascular and neurodegenerative disorders: Molecular mechanisms and possible therapeutic approaches. Redox. Biol. 2019, 21, 101059. [Google Scholar] [CrossRef]
- Wang, H.M.; Fu, L.; Cheng, C.C.; Gao, R.; Lin, M.Y.; Su, H.L.; Belinda, N.E.; Nguyen, T.H.; Lin, W.H.; Lee, P.C.; et al. Inhibition of LPS-Induced Oxidative Damages and Potential Anti-Inflammatory Effects of Phyllanthus emblica Extract via Down-Regulating NF-kappaB, COX-2, and iNOS in RAW 264.7 Cells. Antioxidants 2019, 8, 270. [Google Scholar] [CrossRef]
- Xing, J.S.; Wang, X.; Lan, Y.L.; Lou, J.C.; Ma, B.; Zhu, T.; Zhang, H.; Wang, D.; Yu, Z.; Yuan, Z.; et al. Isoalantolactone inhibits IKKbeta kinase activity to interrupt the NF-kappaB/COX-2-mediated signaling cascade and induces apoptosis regulated by the mitochondrial translocation of cofilin in glioblastoma. Cancer Med. 2019, 8, 1655–1670. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Pachori, A.S.; Ward, C.A.; Davis, J.P.; Gnecchi, M.; Kong, D.; Zhang, L.; Murduck, J.; Yet, S.F.; Perrella, M.A.; et al. Heme oxygenase-1 (HO-1) inhibits postmyocardial infarct remodeling and restores ventricular function. FASEB J. 2006, 20, 207–216. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences | Length (bp) |
|---|---|---|
| IL-6 | (F) 5′-GACAAAGCCAGAGTCCTTCAGA-3′ (R) 5′-TGTGACTCCAGCTTATCTCTTGG-3′ | 76 |
| TNF-α | (F) 5′-ACTGAACTTCGGGGTGATCG-3′ (R) 5′-TGGTGGTTTGTGAGTGTGAGG-3′ | 102 |
| iNOS | (F) 5′-CAACAGGGAGAAAGCGCAAAA-3′ (R) 5′-TACTGTGGACGGGTCGATGT-3′ | 175 |
| COX-2 | (F) 5′-TGAGTACCGCAAACGCTTCT-3′ (R) 5′- CAGCCATTTCCTTCTCTCCTGT-3′ | 74 |
| β-actin | (F) 5′-GTCAGGTCATCACTATCGGCAAT-3′ (R) 5′-AGAGGTCTTTACGGATGTCAACGT-3′ | 147 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, D.; Liu, Y.; Li, J.; Wang, H.; Ye, B.; He, Y.; Li, Z.; Gao, X.; Fu, S.; Liu, D. Isoalantolactone (IAL) Regulates Neuro-Inflammation and Neuronal Apoptosis to Curb Pathology of Parkinson’s Disease. Cells 2022, 11, 2927. https://doi.org/10.3390/cells11182927
He D, Liu Y, Li J, Wang H, Ye B, He Y, Li Z, Gao X, Fu S, Liu D. Isoalantolactone (IAL) Regulates Neuro-Inflammation and Neuronal Apoptosis to Curb Pathology of Parkinson’s Disease. Cells. 2022; 11(18):2927. https://doi.org/10.3390/cells11182927
Chicago/Turabian StyleHe, Dewei, Yanting Liu, Jie Li, Hefei Wang, Bojian Ye, Yuan He, Zhe Li, Xiyu Gao, Shoupeng Fu, and Dianfeng Liu. 2022. "Isoalantolactone (IAL) Regulates Neuro-Inflammation and Neuronal Apoptosis to Curb Pathology of Parkinson’s Disease" Cells 11, no. 18: 2927. https://doi.org/10.3390/cells11182927